- Виды иммунитета

Содержание
- 2. Антигены – любые вещества, чаще белки или гликопротеиды, которые, попадая в организм, вызывают образование специфических антител
- 3. Рецепторы – макромолекулы на клетках, специфически связывающие различные биологически активные вещества (лиганды) Цитокины – медиаторы межклеточных
- 4. Иммунитет – эволюционно обусловленная совокупность реакций взаимодействия между системой иммунитета и биологически активными агентами (антигенами) Эти
- 5. Полезные феномены: Противоинфекционный иммунитет –невосприимчивость организма к конкретным инфекционным агентам возбудителям заболеваний (микробам, вирусам) Толерантность –
- 6. Патологические феномены: Гиперчувствительность – повышенная реакция СИ на антигены-аллергены – приводит к развитию двух видов патологии:
- 7. Виды иммунитета Видовой иммунитет (конституциональный, наследственный) – вариант неспецифической резистентности организма, генетически обусловленный особенностями обмена веществ
- 8. Естественный врожденный иммунитет (неспецифические естественные факторы иммунитета): Гуморальные факторы: система комплемента, С-реактивный белок, фермент лизоцим, интерфероны,
- 9. Приобретенный (адаптивный) иммунитет возникает в течение жизни каждого индивида, может быть противоинфекционный и неинфекционный Противоинфекционный: антимикробный
- 10. Искусственный активный иммунитет возникает после введения в организм вакцин и анатоксинов, которые содержат микроорганизмы или их
- 11. Виды неинфекционного иммунитета: Трансплантационный иммунитет возникает при пересадке органов и тканей от донора к реципиенту, в
- 12. Репродуктивный иммунитет в системе "мать-плод". Это совокупность реакций матери на антигены плода, так как он отличается
- 13. Выживаемость плода обеспечивают: особая организация тканей между матерью и плодом (трофобласт и др.) защитное влияние АТ
- 14. Противоопухолевый иммунитет направлен против антигенов опухолевых клеток и включает: 1. естественный неспецифический иммунитет естественные киллеры активированные
- 15. приобретенный специфический противоопухолевый иммунитет лизис комплементом опухолевых клеток, покрытых антителами (в основном в крови) лизис лейкоцитами
- 16. Факторы неэффективности противоопухолевого иммунитета скорость размножения опухолевых клеток выше, чем формирование эффекторов иммунитета лейкоциты, связавшие противоопухолевые
- 17. Опухоль выделяет цитокины, подавляющие иммунный ответ, экспрессию HLA молекул, блокирующие активность цитотоксических лимфоцитов постоянно секретируeт толерогенные
- 18. Аутоиммунитет («аутоаллергия») – реакции системы иммунитета на собственные антигены (белки, липопротеиды, гликопротеиды). Это обусловлено нарушением распознавания
- 19. Система иммунитета (СИ) – совокупность молекул, клеток, тканей и органов, осуществляющих иммунные реакции Лимфоидная система включает
- 20. Система гранулоцитов объединяет нейтрофильные лейкоциты, базофильные лейкоциты/тучные клетки, эозинофильные лейкоциты Система мононуклеарных фагоцитов (моноциты, макрофаги тканей
- 21. К центральным органам системы иммунитета относятся красный костный мозг и тимус К периферическим – циркулирующие лимфоциты
- 22. Цитокины и интерлейкины Дифференцировка и взаимодействие клеток системы иммунитета между собой, а также с клетками других
- 23. Общие свойства цитокинов Гликопротеины с молекулярной массой 15-25 кД. Действуют ауто- и паракринно (т.е. на саму
- 24. Классификация цитокинов Интерлейкины (ИЛ) Интерфероны Группа факторов некроза опухоли (ФНО) Группа колониестимулирующих факторов (например, гранулоцитарно-макрофагальный колониестимулирующий
- 25. Интерлейкины Цитокины, выделяемые преимущественно клетками системы иммунитета, получили название интерлейкинов (ИЛ) – факторов межлейкоцитарного взаимодействия ИЛ-1
- 26. ИЛ-2 выделяется Т-хелперами (преимущественно Тх1) и стимулирует пролиферацию и дифференцировку Т- и В-лимфоцитов, ЕКК, моноцитов ИЛ-3
- 27. , ИЛ-5 стимулирует созревание эозинофилов, базофилов и синтез иммуноглобулинов В-лимфоцитами, вырабатывается Т-лимфоцитами под влиянием антигенов ИЛ-6
- 28. ИЛ-10 – выделяется Т-лимфоцитами ( Тх2 и регуляторными Т-хелперами – Tr). Подавляет выделение провоспалительных цитокинов (ИЛ-1,
- 29. Провоспалительные цитокины (ИЛ-1α, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ИНФ-γ, ФНОα и др.) Противоспалительные цитокины (ИЛ4, ИЛ-10, ИЛ-13,
- 30. Молекулы дифференцировки клеток системы иммунитета – CD-антигены В процессе дифференцировки на мембранах клеток системы иммунитета появляются
- 31. CD1 – является общим антигеном тимоцитов CD2 – общий маркер всех Т-клеток, обладает способностью связывать эритроциты
- 32. CD8 – маркер Т-цитотоксических лимфоцитов (Т-киллеров)/Т-супрессоров, участвует в распознавании антигенов, ассоциированных с молекулами HLA I класса
- 33. CD16 – несут нейтрофилы, ЕК, моноциты CD19-22 – маркеры В-лимфоцитов CD34 – имеют все предшественники гемопоэза
- 34. Toll-like рецепторы Распознают типовые структурные компоненты или «Молекулярные образы патогенов», которые сходны у больших групп патогенных
- 35. TLR-1 связывает липопептиды различных групп бактерий TLR-2 взаимодействует с липотейхоевыми кислотами большинства грамположительных бактерий, липопротеинами боррелий,
- 36. TLR-3 связывается с двухцепочечной РНК, что является важным для эффективного противовирусного иммунитета TLR-4 реагирует с ЛПС
- 37. Функции системы TLR Усиление экспрессии костимуляторных молекул на АПК Без костимуляции Т-лимфоциты переходят в состояние неотвечаемости
- 38. Разный цитокиновый профиль стимулирует превращение Тх0 либо в Тх1, либо в Тх2 Активация Тх1 приводит к
- 39. Дифференцировка Т-лимфоцитов ГСК → ЛСК → про Тл (CD34, CD7) Антигеннезависимая дифференцировка тимус → пре Тл
- 40. Антигензависимая дифференцировка Т-лимфоцитов Тх0 → Тх1 (ИЛ2, γ-интерферон), клеточный иммунитет Тх0 → Тх2 (ИЛ4, ИЛ5, ИЛ10,
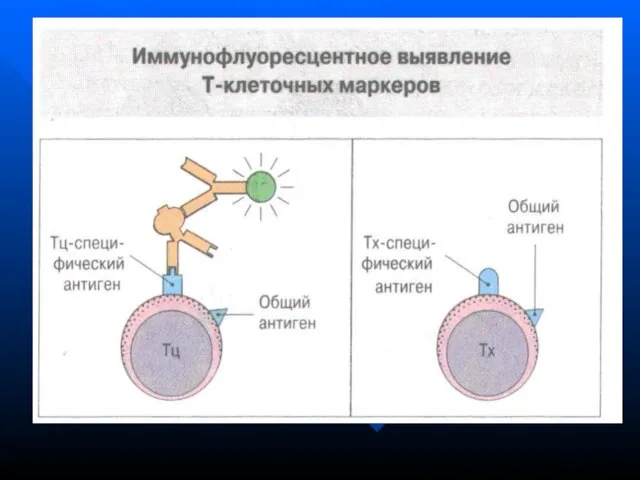
- 41. Основные субпопуляции: Т0 (нулевые, тимические, «наивные», незрелые) Т-цитотоксические Т-хелперы (Тх1, Тх2, Тх3) Т-памяти В норме у
- 42. Т-клеточный рецептор
- 43. ТКР у Тх и Тц одинаковы по строению Тх распознают комплексы АГ+HLAII, причем АГ в виде
- 45. Дифференцировка В-лимфоцитов Процесс созревания стимулируется цитокинами: ИЛ3, ИЛ7, ИЛ4 Антигеннезависимая дифференцировка ГСК (CD34) → ЛСК →
- 46. пIgM может взаимодействовать с АГ Igα и Igβ - передают сигнал в клетку Зрелый Вл (ВКР,
- 48. На поверхности В-лимфоцитов имеются дифференцировочные антигены: СD19, 20, 22, 40, 72, рецепторы к эритроцитам мыши, к
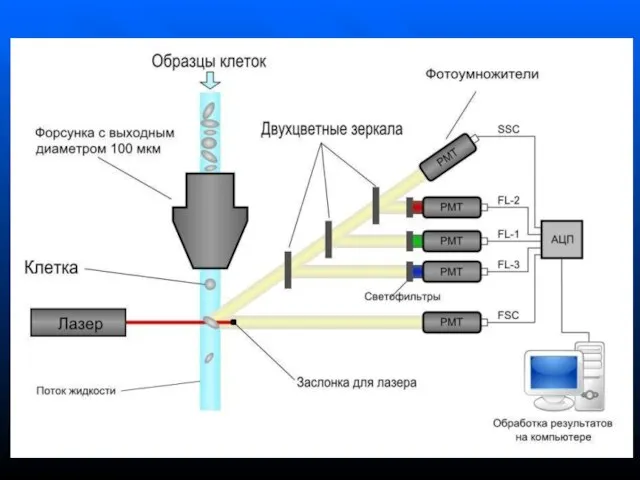
- 49. Проточная цитометрия Метод оптического измерения параметров клетки, ее органелл и происходящих в ней процессов Выявляют рассеяние
- 50. Клеточная суспензия, предварительно меченная флюоресцирующими моноклональными АТ или флуоресцентными красителями, попадает в поток жидкости, проходящий через
- 51. В момент пересечения клеткой лазерного луча детекторы фиксируют: рассеяние света под малыми углами (от 1° до
- 52. Применение Для выявления определённых клеток в исследуемых образцах (бактерий, грибов, собственных клеток организма человека) Определения чувствительности
- 56. Иммуноглобулины (Ig) – большое семейство белков, которые синтезируются В-л и плазмоцитами при электрофорезе образуют фракцию γ-глобулинов
- 57. Структура Ig 2 тяжелые полипептидные цепи (H – heavy) 2 легкие (L – light) полипептидные цепи
- 58. В цепях Ig различают константные (c) и вариабельные (v) участки. Участки Ig цепей, замкнутые в виде
- 59. Между СН1 и СН2 доменами тяжелой цепи локализуется подвижный – "шарнирный" участок Ig, чувствительный к протеолитическим
- 60. Функции Fc-фрагмента CН2 домен Fc-фрагмента активирует комплемент по классическому пути (после образования комплекса АГ+АТ) СH3 домен
- 62. Свойства иммуноглобулинов IgG составляют основную массу иммуноглобулинов сыворотки крови (75-85%) – 10 г/л (8-12 г/л) Передаются
- 63. IgМ – пентамер, в сыворотке крови в среднем – 1 г/л (0.8-1.5 г/л) АТ IgM синтезируются
- 64. IgА в крови присутствуют в виде мономеров (от 1,5 до 3 г/л), а в секретах в
- 66. IgD содержатся в сыворотке крови в концентрации 0,03-0,04 г/л Служат рецепторами созревающих В-лимфоцитов Увеличиваются при некоторых
- 67. IgЕ присутствуют в сыворотке крови в концентрации около 0,00005 г/л или от 0 до 100 МЕ/мл
- 68. Антитела Это Ig любого из 5 классов, специфично взаимодействующие с АГ Естественные АТ находятся в организме
- 69. Механизмы действия антител нейтрализация активных центров токсинов (токсиннейтрализующий эффект) образование комплекса АГ-АТ, который активирует комплемент с
- 70. опсонизация объектов фагоцитоза (усиление фагоцитоза) связывание с Fc-рецепторами лейкоцитов, которые приобретают способность специфично взаимодействовать с антигенами
- 71. антирецепторныеАТ, связываясь с соответствующим рецептором, блокируют или стимулируют функцию клетки АТ обладают собственной медленной ферментативной активностью
- 72. Бивалентные полные АТ (обычно IgG) имеют 2 активных центра Моновалентные неполные АТ, у которых один активный
- 73. Сила связывания (сродство) одного активного центра АТ с эпитопом АГ- аффинность Прочность связывания всей молекулы Ig
- 74. Поликлональные АТ сыворотки представляют собой смесь Ig различных классов Моноклональные АТ разработаны на основе гибридомной технологии,
- 75. Получение моноклональных АТ Иммунизация мышей антигеном Из селезенки иммунизированных мышей получают суспензию клеток, среди которых есть
- 76. Проводят слияние этих антителообразующих В-клеток с В-клетками мышиной опухоли – плазмоцитомы («бессмертные» клетки) Образуются гибридные клетки,
- 77. Их культивируют в специальной среде, в которой не растут обычные негибридные клетки Из смеси гибридных клеток
- 78. Применение моноклональных АТ Выявление АГ бактерий и вирусов Выявление клеточных маркеров (CD) Выявление гормонов, медиаторов и
- 79. Генетические основы разнообразия АТ и ТКР H и L цепи Ig кодируются несколькими генными сегментами, расположенными
- 80. Каждый такой сегмент формируется из нескольких участков ДНК При объединении фрагментов генома в единую последовательность ДНК
- 81. Антигены (АГ) - любые простые или сложные вещества, которые при попадании в организм тем или иным
- 82. Основные свойства антигенов: Иммуногенность – способность АГ индуцировать в организме иммунную реакцию Специфичность – способность взаимодействовать
- 83. Иммуногенность обусловлена сложностью пространственной структуры молекулы (боковые радикалы, разветвленные цепи и т.п.) Эпитоп (антигенная детерминанта )
- 85. Гаптены - низкомолекулярные вещества, в обычных условиях не вызывают иммунную реакцию Способны запускать иммунный ответ после
- 86. Аллергены - АГили гаптены, которые при повторном попадании в организм вызывают аллергическую реакцию. Все АГ и
- 87. Т-зависимые АГ- запускают иммунный ответ с участием Т-лимфоцитов (белки) Т-независимые АГ- запускают иммунный ответ и синтез
- 88. Экзогенные и эндогенные АГ Экзогенные антигены попадают в организм из внешней среды. Среди них различают инфекционные
- 89. Инфекционные АГ- антигены бактерий, вирусов, грибов, простейших, токсины, ферменты ⇒ группоспецифические (встречаются у разных видов одного
- 90. По локализации: О-АГ Н-АГ К-АГ О-АГ - полисахарид, входит в состав к л е т о
- 91. Строение ЛПС Центральная часть ЛПС – полисахарид, состоит из остатков 2-кето-3-дезоксиоктоната, галактозы, глюкозы, гептозы и N-ацетилглюкозамина
- 92. Липид А - неспецифический иммуностимулятор, токсичен ЛПС – эндотоксин и пироген В небольших дозах активирует макрофаги
- 93. Н-АГ входит в состав жгутиков, состоит из белка флагеллина, термолабилен К-АГ - гетерогенная группа поверхностных капсульных
- 94. Протективные антигены Это совокупность антигенных детерминант (эпитопов), которые вызывают наиболее сильный иммунный ответ, что предохраняет организм
- 95. Антигенная мимикрия- сходство в строении АГ бактерий, человека, животных Гемолитические стрептококки группы А содержат АГ, общие
- 96. У возбудителя сифилиса есть фосфолипиды, сходные по строению с кардиолипидным АГ сердца животных и человека Поэтому
- 97. Суперантигены - в небольших дозах вызывают поликлональную активацию и пролиферацию большого числа Т-лимфоцитов (более 20%, обычные
- 98. НЕИНФЕКЦИОННЫЕ АНТИГЕНЫ АГ растений, лекарственные препараты, химические, природные и синтетические вещества, антигены животных и человека
- 99. Антигены животных по отношению к человеку являются ксеногенными Антигены, отличающие одного индивидуума от другого, называют аллогенными
- 100. Антигены эритроцитов На поверхности эритроцитов имеется более 100 антигенов, относящихся к 14 системам Наиболее важными являются
- 101. Антигены лейкоцитов HLA (Human Leucocyte Antigens) контролируются генами 6 хромосомы (главным комплексом гистосовместимости) - Обусловливают несовместимость
- 102. Молекулы HLA I класса – гетеродимеры, состоят из двух цепей Легкая цепь - β2-микроглобулин Тяжелая цепь
- 104. Молекулы HLA II класса состоят из двух полипептидных цепей: α и β. Обе цепи имеют по
- 106. Эндогенные антигены - собственные аутологичные молекулы (аутоантигены), вызывающие активацию системы иммунитета - естественные первичные (нормальная ткань
- 107. По тканевой и клеточной принадлежности - Стромальные (АГ эластических, коллагеновых и других волокон) - Клеточные (мембранные,
- 108. Система мононуклеарных фагоцитов Моноциты крови Оседлые макрофаги печени (клетки Купфера) Мезангиальные клетки Альвеолярные макрофаги Глиальные макрофаги
- 109. Молекулярные маркеры моноцитов и макрофагов CD 14 – рецептор для ЛПС связывающего белка рецепторы для Fc-фрагмента
- 110. Функции макрофагов: фагоцитоз распознавание и представление (презентация) антигенов секреция медиаторов системы □иммунитета (монокинов)
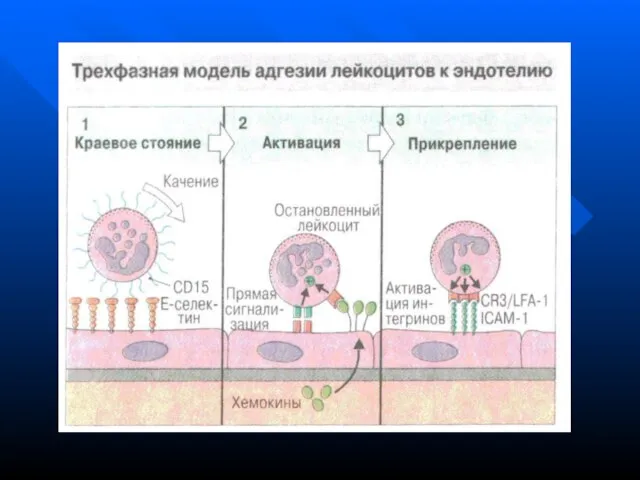
- 111. Стадии фагоцитоза хемотаксис адгезия эндоцитоз (поглощение) переваривание удаление продуктов фагоцитоза
- 112. Хемотаксис - движение макрофагов к объекту фагоцитоза по градиенту концентрации хемотаксических факторов (бактериальных компонентов, интерлейкинов (IL-8)
- 113. Адгезия реализуется двумя механизмами: иммунным и неиммунным Неиммунный фагоцитоз осуществляется за счет связывания антигена на поверхности
- 114. В связывании могут участвовать самые разнообразные молекулы и рецепторы Лектины микробных клеток Лектины фагоцитарных клеток (рецепторы
- 116. Эндоцитоз (поглощение) инвагинация мембраны фагоцита обволакивание объекта фагоцитоза образование фагосомы слияние фагосомы с лизосомами и образование
- 117. Переваривание Кислородзависимые механизмы (образование активных форм кислорода: Н2О2, оксидаза, миелопероксидаза и др.) Кислороднезависимые механизмы
- 118. Кислородзависимые бактерицидные механизмы - образование активных форм кислорода (АФК) через гексозомонофосфатный шунт - «дыхательный взрыв»
- 119. Кислороднезависимые бактерицидные механизмы: Ферменты-гидролазы - катепсины, лизоцим, глюкуронидаза и т.д. Катионные антибиотикоподобные белки - дефензимы вызывают
- 120. Завершенный фагоцитоз - полное переваривание и гибель бактериальной клетки Незавершенный фагоцитоз - микробные клетки остаются жизнеспособными.
- 121. Представление (презентация) антигенов макрофагами В результате фагоцитоза и переваривания антигенов образуется большое количество низкомолекулярных антигенных фрагментов
- 122. Экзоантигены - пептиды длиной 12-25 аминокислот связываются с HLA 2 класса (HLA-DR, HLA-DP, HLA-DQ). Только после
- 123. Секреция медиаторов иммунной системы (монокинов). Второй сигнал для активации Т-хелперов - выделение макрофагами ИЛ1 - монокина
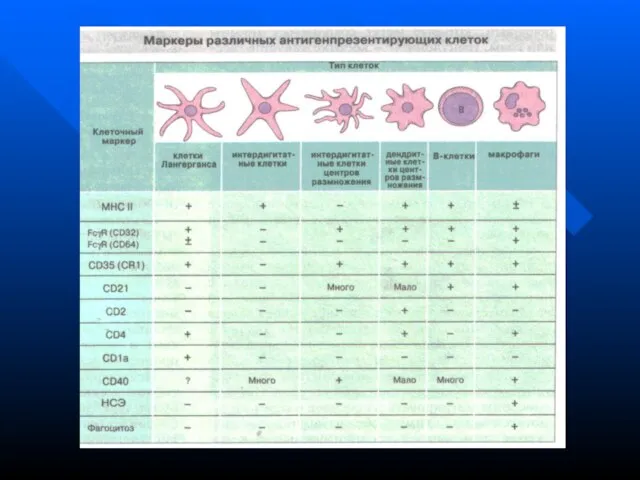
- 124. ДРУГИЕ АНТИГЕН-ПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ (АПК) Клетки Лангерганса из кожи мигрируют в паракортикальные области регионарных лимфоузлов. Там они
- 126. СИСТЕМА ГРАНУЛОЦИТОВ В нее входят нейтрофильные, базофильные и эозинофильные гранулоциты (микрофаги). Все они происходят из ГСК
- 128. Нейтрофилы составляют 47-72% всех лейкоцитов. Их содержится 2.5 - 4.5х109/л. Средний срок жизни нейтрофилов - 7-9
- 129. Азурофильные гранулы содержат β-глюкуронидазу, катепсины, кислые и нейтральные протеиназы, миелопероксидазу. В специфических гранулах находятся коллагеназа, лизоцим,
- 131. Основные молекулы-маркеры на нейтрофилах : CD13,СД 14, СD64 , рецепторы к С1q, С3b, С5а компонентам комплемента,CD32(FcγRII)
- 132. Базофилы участвуют в аллергических реакциях. На поверхности базофилов имеются Fcε-рецепторы, связывающие IgE. В гранулах базофилов содержится
- 133. Эозинофилы играют большую роль в противопаразитарном иммунитете и аллергии. В крови 1-6%, созревают под действием ИЛ-5.
- 134. Характеристика системы макрофагов и гранулоцитов 1. Определяют количество лейкоцитов в крови. 2. Оценивают поглотительную и переваривающую
- 135. Определяют фагоцитарный индекс и фагоцитарное число Фагоцитарный индекс - это среднее количество частиц или микроорганизмов в
- 136. Оценка показателей фагоцитоза В норме через 90 мин фагоцитарный индекс должен быть ниже, чем через 30
- 137. НСТ-тест Метаболическую активность - «дыхательный взрыв» определяют после окраски лейкоцитов раствором нитросинего тетразолия. Под влиянием активных
- 138. С помощью моноклональных антител определяют антигены фагоцитов (СD14, СD11, СD18, HLA-DR и др.). Выявляют рецепторы к
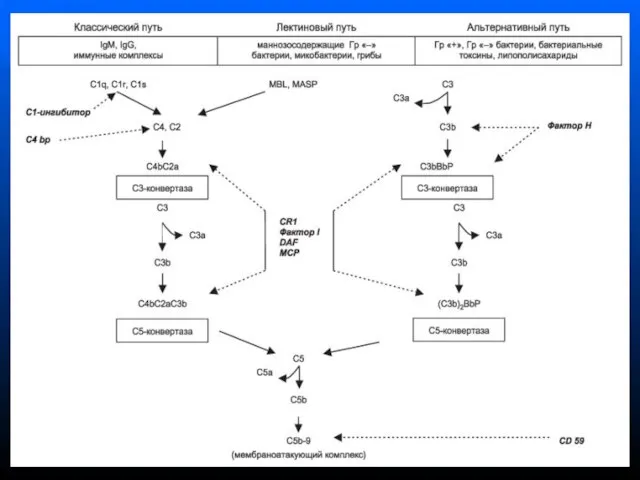
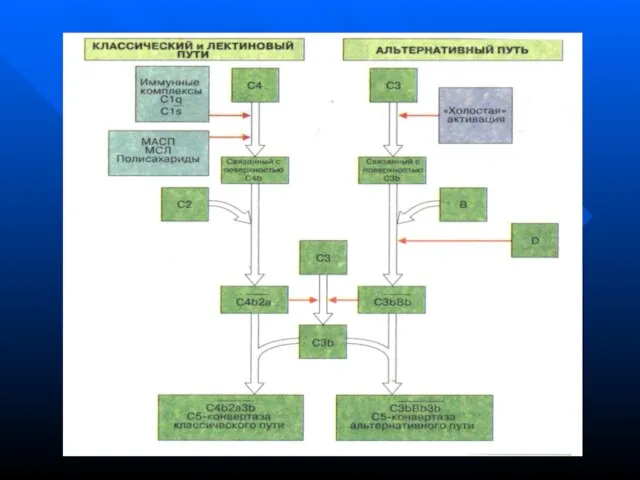
- 139. Система комплемента сложная система ферментативных и рецепторных белков сыворотки крови (более 20) Активация системы комплемента -
- 140. Белки классического пути активации системы комплемента называются компонентами и обозначаются буквой C (C1, С2, С3 и
- 141. При активации компонентов комплемента обычно образуется 2 фрагмента Больший фрагмент обозначается малой латинской буквой «b» (напр.,
- 142. Среди регуляторных белков различают естественный С1-ингибитор, который тормозит спонтанную активацию C1q компонента При дефиците С1 ингибитора
- 143. Пути активации системы комплемента Классический (запускается комплексом АГ-АТ в присутствии катионов Ca и Mg обычно на
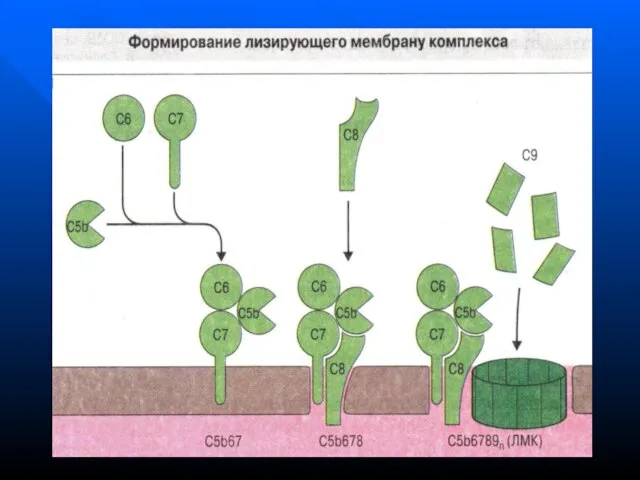
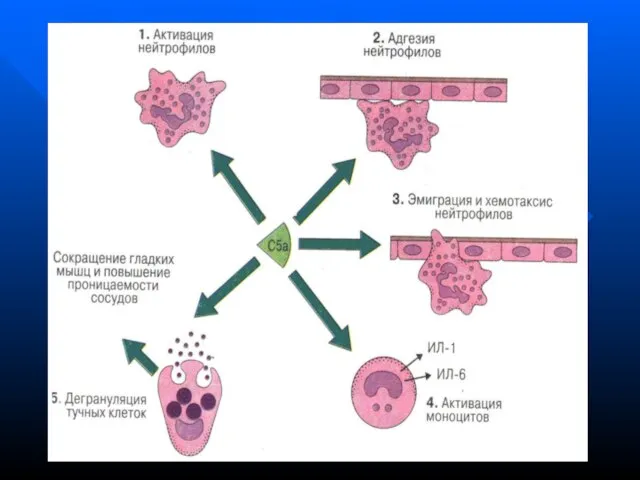
- 148. Функции системы комплемента Лизис клеток-мишеней (бактериальных, зараженных вирусом, опухолевых) Опсонизация, т.е. усиление фагоцитоза через рецепторы к
- 150. Скачать презентацию

Слайд 3Рецепторы – макромолекулы на клетках, специфически связывающие различные биологически активные вещества (лиганды)
Цитокины
Рецепторы – макромолекулы на клетках, специфически связывающие различные биологически активные вещества (лиганды)
Цитокины

Слайд 4Иммунитет – эволюционно обусловленная совокупность реакций взаимодействия между системой иммунитета и биологически
Иммунитет – эволюционно обусловленная совокупность реакций взаимодействия между системой иммунитета и биологически

Слайд 5
Полезные феномены:
Противоинфекционный иммунитет –невосприимчивость организма к конкретным инфекционным агентам
Полезные феномены:
Противоинфекционный иммунитет –невосприимчивость организма к конкретным инфекционным агентам

Слайд 6 Патологические феномены:
Гиперчувствительность – повышенная реакция СИ на антигены-аллергены – приводит
Патологические феномены:
Гиперчувствительность – повышенная реакция СИ на антигены-аллергены – приводит

Слайд 7Виды иммунитета
Видовой иммунитет (конституциональный, наследственный) – вариант неспецифической резистентности организма, генетически
Виды иммунитета
Видовой иммунитет (конституциональный, наследственный) – вариант неспецифической резистентности организма, генетически

Слайд 8Естественный врожденный иммунитет (неспецифические естественные факторы иммунитета):
Гуморальные факторы:
система комплемента, С-реактивный белок, фермент
Естественный врожденный иммунитет (неспецифические естественные факторы иммунитета):
Гуморальные факторы:
система комплемента, С-реактивный белок, фермент

Слайд 9Приобретенный (адаптивный) иммунитет возникает в течение жизни каждого индивида, может быть противоинфекционный
Приобретенный (адаптивный) иммунитет возникает в течение жизни каждого индивида, может быть противоинфекционный

Слайд 10Искусственный активный иммунитет возникает после введения в организм вакцин и анатоксинов, которые

Слайд 11Виды неинфекционного иммунитета:
Трансплантационный иммунитет возникает при пересадке органов и тканей от донора
Трансплантационный иммунитет возникает при пересадке органов и тканей от донора

Слайд 12Репродуктивный иммунитет в системе "мать-плод". Это совокупность реакций матери на антигены плода,

Слайд 13Выживаемость плода обеспечивают:
особая организация тканей между матерью и плодом (трофобласт и
Выживаемость плода обеспечивают:
особая организация тканей между матерью и плодом (трофобласт и

Слайд 14Противоопухолевый иммунитет направлен против антигенов опухолевых клеток и включает:
1. естественный неспецифический иммунитет
естественные
Противоопухолевый иммунитет направлен против антигенов опухолевых клеток и включает:
1. естественный неспецифический иммунитет
естественные

Слайд 15 приобретенный специфический противоопухолевый иммунитет
лизис комплементом опухолевых клеток, покрытых антителами (в основном
приобретенный специфический противоопухолевый иммунитет
лизис комплементом опухолевых клеток, покрытых антителами (в основном

Слайд 16Факторы неэффективности противоопухолевого иммунитета
скорость размножения опухолевых клеток выше, чем формирование эффекторов
Факторы неэффективности противоопухолевого иммунитета
скорость размножения опухолевых клеток выше, чем формирование эффекторов

Слайд 17 Опухоль выделяет цитокины, подавляющие иммунный ответ, экспрессию HLA молекул, блокирующие активность
Опухоль выделяет цитокины, подавляющие иммунный ответ, экспрессию HLA молекул, блокирующие активность

Слайд 18Аутоиммунитет («аутоаллергия») – реакции системы иммунитета на собственные антигены (белки, липопротеиды, гликопротеиды).
Аутоиммунитет («аутоаллергия») – реакции системы иммунитета на собственные антигены (белки, липопротеиды, гликопротеиды).

Слайд 19 Система иммунитета (СИ) – совокупность молекул, клеток, тканей и органов, осуществляющих
Система иммунитета (СИ) – совокупность молекул, клеток, тканей и органов, осуществляющих

Слайд 20Система гранулоцитов объединяет нейтрофильные лейкоциты, базофильные лейкоциты/тучные клетки, эозинофильные лейкоциты
Система мононуклеарных фагоцитов
Система гранулоцитов объединяет нейтрофильные лейкоциты, базофильные лейкоциты/тучные клетки, эозинофильные лейкоциты
Система мононуклеарных фагоцитов

Слайд 21К центральным органам системы иммунитета относятся красный костный мозг и тимус
К периферическим
К центральным органам системы иммунитета относятся красный костный мозг и тимус
К периферическим

Слайд 22Цитокины и интерлейкины
Дифференцировка и взаимодействие клеток системы иммунитета между собой, а
Цитокины и интерлейкины
Дифференцировка и взаимодействие клеток системы иммунитета между собой, а

Слайд 23Общие свойства цитокинов
Гликопротеины с молекулярной массой 15-25 кД.
Действуют ауто- и паракринно (т.е.
Общие свойства цитокинов
Гликопротеины с молекулярной массой 15-25 кД.
Действуют ауто- и паракринно (т.е.

Слайд 24Классификация цитокинов
Интерлейкины (ИЛ)
Интерфероны
Группа факторов некроза опухоли (ФНО)
Группа колониестимулирующих факторов (например, гранулоцитарно-макрофагальный
Классификация цитокинов
Интерлейкины (ИЛ)
Интерфероны
Группа факторов некроза опухоли (ФНО)
Группа колониестимулирующих факторов (например, гранулоцитарно-макрофагальный

Слайд 25Интерлейкины
Цитокины, выделяемые преимущественно клетками системы иммунитета, получили название интерлейкинов (ИЛ) –
Интерлейкины
Цитокины, выделяемые преимущественно клетками системы иммунитета, получили название интерлейкинов (ИЛ) –

Слайд 26ИЛ-2 выделяется Т-хелперами (преимущественно Тх1) и стимулирует пролиферацию и дифференцировку Т- и
ИЛ-2 выделяется Т-хелперами (преимущественно Тх1) и стимулирует пролиферацию и дифференцировку Т- и

Слайд 27,
ИЛ-5 стимулирует созревание эозинофилов, базофилов и синтез иммуноглобулинов В-лимфоцитами, вырабатывается Т-лимфоцитами под
,
ИЛ-5 стимулирует созревание эозинофилов, базофилов и синтез иммуноглобулинов В-лимфоцитами, вырабатывается Т-лимфоцитами под

Слайд 28ИЛ-10 – выделяется Т-лимфоцитами ( Тх2 и регуляторными Т-хелперами – Tr). Подавляет
ИЛ-10 – выделяется Т-лимфоцитами ( Тх2 и регуляторными Т-хелперами – Tr). Подавляет

Слайд 29Провоспалительные цитокины (ИЛ-1α, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ИНФ-γ, ФНОα и др.)
Противоспалительные цитокины
Провоспалительные цитокины (ИЛ-1α, ИЛ-6, ИЛ-8, ИЛ-12, ИЛ-18, ИНФ-γ, ФНОα и др.)
Противоспалительные цитокины

Слайд 30Молекулы дифференцировки клеток системы иммунитета – CD-антигены
В процессе дифференцировки на мембранах
Молекулы дифференцировки клеток системы иммунитета – CD-антигены
В процессе дифференцировки на мембранах

Слайд 31CD1 – является общим антигеном тимоцитов
CD2 – общий маркер всех Т-клеток, обладает
CD2 – общий маркер всех Т-клеток, обладает

Слайд 32CD8 – маркер Т-цитотоксических лимфоцитов (Т-киллеров)/Т-супрессоров, участвует в распознавании антигенов, ассоциированных с

Слайд 33CD16 – несут нейтрофилы, ЕК, моноциты
CD19-22 – маркеры В-лимфоцитов
CD34 – имеют все
CD19-22 – маркеры В-лимфоцитов
CD34 – имеют все

Слайд 34Toll-like рецепторы
Распознают типовые структурные компоненты или «Молекулярные образы патогенов», которые
Toll-like рецепторы
Распознают типовые структурные компоненты или «Молекулярные образы патогенов», которые

Слайд 35TLR-1 связывает липопептиды различных групп бактерий
TLR-2 взаимодействует с липотейхоевыми кислотами большинства грамположительных
TLR-2 взаимодействует с липотейхоевыми кислотами большинства грамположительных

Слайд 36TLR-3 связывается с двухцепочечной РНК, что является важным для эффективного противовирусного иммунитета
TLR-4
TLR-4

Слайд 37Функции системы TLR
Усиление экспрессии костимуляторных молекул на АПК
Без костимуляции Т-лимфоциты
Функции системы TLR
Усиление экспрессии костимуляторных молекул на АПК
Без костимуляции Т-лимфоциты

Слайд 38Разный цитокиновый профиль стимулирует превращение Тх0 либо в Тх1, либо в Тх2

Слайд 39Дифференцировка Т-лимфоцитов
ГСК → ЛСК → про Тл (CD34, CD7)
Антигеннезависимая дифференцировка
тимус
Дифференцировка Т-лимфоцитов
ГСК → ЛСК → про Тл (CD34, CD7)
Антигеннезависимая дифференцировка
тимус

Слайд 40Антигензависимая дифференцировка
Т-лимфоцитов
Тх0 → Тх1 (ИЛ2, γ-интерферон), клеточный иммунитет
Тх0 → Тх2 (ИЛ4,
Т-лимфоцитов
Тх0 → Тх1 (ИЛ2, γ-интерферон), клеточный иммунитет
Тх0 → Тх2 (ИЛ4,

Слайд 41 Основные субпопуляции:
Т0 (нулевые, тимические, «наивные», незрелые)
Т-цитотоксические
Основные субпопуляции:
Т0 (нулевые, тимические, «наивные», незрелые)
Т-цитотоксические

Слайд 42Т-клеточный рецептор
Т-клеточный рецептор
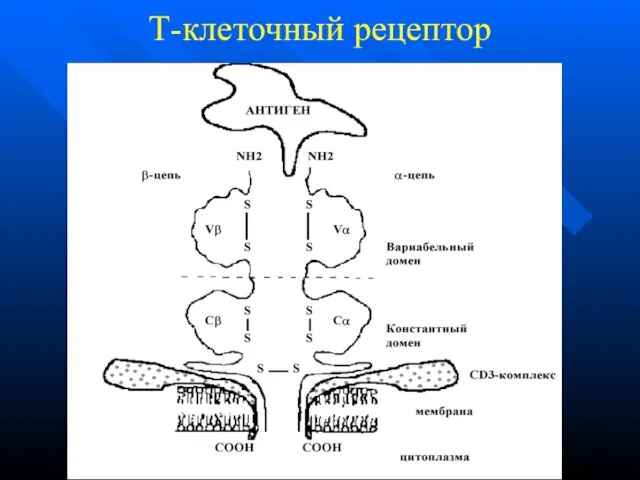
Слайд 43 ТКР у Тх и Тц одинаковы по строению
Тх распознают комплексы
ТКР у Тх и Тц одинаковы по строению
Тх распознают комплексы

Слайд 45Дифференцировка В-лимфоцитов
Процесс созревания стимулируется цитокинами: ИЛ3, ИЛ7, ИЛ4
Антигеннезависимая дифференцировка
ГСК (CD34) → ЛСК
Дифференцировка В-лимфоцитов
Процесс созревания стимулируется цитокинами: ИЛ3, ИЛ7, ИЛ4
Антигеннезависимая дифференцировка
ГСК (CD34) → ЛСК

Слайд 46пIgM может взаимодействовать с АГ
Igα и Igβ - передают сигнал
Igα и Igβ - передают сигнал

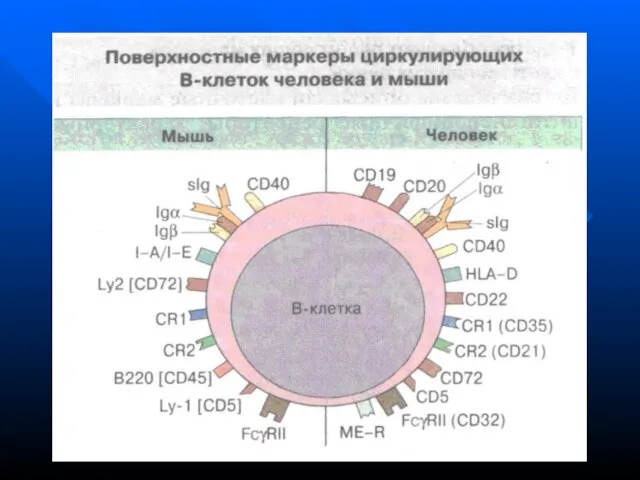
Слайд 48На поверхности В-лимфоцитов имеются дифференцировочные антигены: СD19, 20, 22, 40, 72, рецепторы

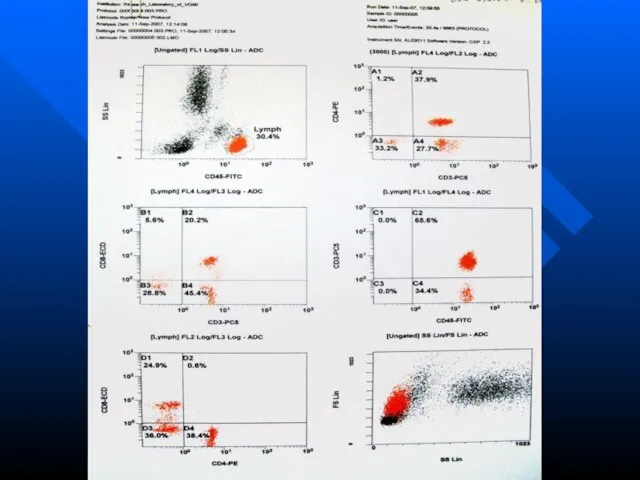
Слайд 49Проточная цитометрия
Метод оптического измерения параметров клетки, ее органелл и происходящих в ней
Проточная цитометрия
Метод оптического измерения параметров клетки, ее органелл и происходящих в ней

Слайд 50Клеточная суспензия, предварительно меченная флюоресцирующими моноклональными АТ или флуоресцентными красителями, попадает в
Клеточная суспензия, предварительно меченная флюоресцирующими моноклональными АТ или флуоресцентными красителями, попадает в

Слайд 51В момент пересечения клеткой лазерного луча детекторы фиксируют:
рассеяние света под малыми углами
рассеяние света под малыми углами

Слайд 52Применение
Для выявления определённых клеток в исследуемых образцах (бактерий, грибов, собственных клеток организма
Применение
Для выявления определённых клеток в исследуемых образцах (бактерий, грибов, собственных клеток организма


Слайд 56Иммуноглобулины (Ig) – большое семейство белков, которые синтезируются В-л и плазмоцитами

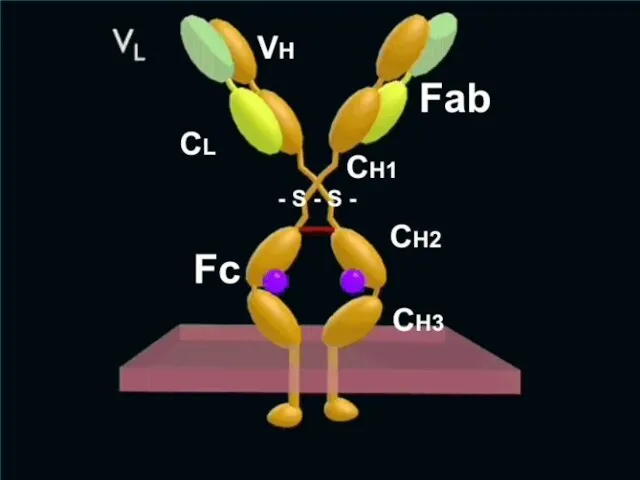
Слайд 57Структура Ig
2 тяжелые полипептидные цепи (H – heavy)
2 легкие (L
2 тяжелые полипептидные цепи (H – heavy)
2 легкие (L

Слайд 58В цепях Ig различают константные (c) и вариабельные (v) участки.
Участки Ig
Участки Ig

Слайд 59Между СН1 и СН2 доменами тяжелой цепи локализуется подвижный – "шарнирный" участок

Слайд 60Функции Fc-фрагмента
CН2 домен Fc-фрагмента активирует комплемент по классическому пути (после образования
Функции Fc-фрагмента
CН2 домен Fc-фрагмента активирует комплемент по классическому пути (после образования

Слайд 62Свойства иммуноглобулинов
IgG составляют основную массу иммуноглобулинов сыворотки крови (75-85%) – 10
Свойства иммуноглобулинов
IgG составляют основную массу иммуноглобулинов сыворотки крови (75-85%) – 10

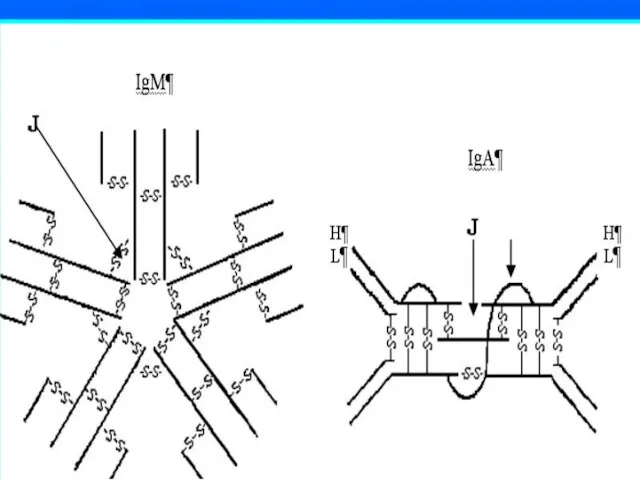
Слайд 63IgМ – пентамер, в сыворотке крови в среднем – 1 г/л (0.8-1.5

Слайд 64IgА в крови присутствуют в виде мономеров (от 1,5 до 3 г/л),

Слайд 66
IgD содержатся в сыворотке крови в концентрации 0,03-0,04 г/л
Служат
IgD содержатся в сыворотке крови в концентрации 0,03-0,04 г/л
Служат

Слайд 67IgЕ присутствуют в сыворотке крови в концентрации около 0,00005 г/л или от

Слайд 68Антитела
Это Ig любого из 5 классов, специфично взаимодействующие с АГ
Естественные АТ
Антитела
Это Ig любого из 5 классов, специфично взаимодействующие с АГ
Естественные АТ

Слайд 69Механизмы действия антител
нейтрализация активных центров токсинов (токсиннейтрализующий эффект)
образование комплекса АГ-АТ,
Механизмы действия антител
нейтрализация активных центров токсинов (токсиннейтрализующий эффект)
образование комплекса АГ-АТ,

Слайд 70опсонизация объектов фагоцитоза (усиление фагоцитоза)
связывание с Fc-рецепторами лейкоцитов, которые приобретают способность специфично
связывание с Fc-рецепторами лейкоцитов, которые приобретают способность специфично

Слайд 71антирецепторныеАТ, связываясь с соответствующим рецептором, блокируют или стимулируют функцию клетки
АТ обладают
АТ обладают

Слайд 72Бивалентные полные АТ (обычно IgG) имеют 2 активных центра
Моновалентные неполные АТ,
Моновалентные неполные АТ,

Слайд 73Сила связывания (сродство) одного активного центра АТ с эпитопом АГ- аффинность
Прочность связывания
Прочность связывания

Слайд 74Поликлональные АТ сыворотки представляют собой смесь Ig различных классов
Моноклональные АТ разработаны
Моноклональные АТ разработаны

Слайд 75Получение моноклональных АТ
Иммунизация мышей антигеном
Из селезенки иммунизированных мышей получают суспензию клеток,
Получение моноклональных АТ
Иммунизация мышей антигеном
Из селезенки иммунизированных мышей получают суспензию клеток,

Слайд 76Проводят слияние этих антителообразующих В-клеток с В-клетками мышиной опухоли – плазмоцитомы («бессмертные»

Слайд 77Их культивируют в специальной среде, в которой не растут обычные негибридные клетки

Слайд 78Применение моноклональных АТ
Выявление АГ бактерий и вирусов
Выявление клеточных маркеров (CD)
Выявление гормонов, медиаторов
Применение моноклональных АТ
Выявление АГ бактерий и вирусов
Выявление клеточных маркеров (CD)
Выявление гормонов, медиаторов

Слайд 79Генетические основы разнообразия АТ и ТКР
H и L цепи Ig кодируются несколькими
Генетические основы разнообразия АТ и ТКР
H и L цепи Ig кодируются несколькими

Слайд 80Каждый такой сегмент формируется из нескольких участков ДНК
При объединении фрагментов генома в
При объединении фрагментов генома в

Слайд 81Антигены (АГ) - любые простые или сложные вещества, которые при попадании в
Антигены (АГ) - любые простые или сложные вещества, которые при попадании в

Слайд 82 Основные свойства антигенов:
Иммуногенность – способность АГ индуцировать в организме иммунную
Основные свойства антигенов: Иммуногенность – способность АГ индуцировать в организме иммунную

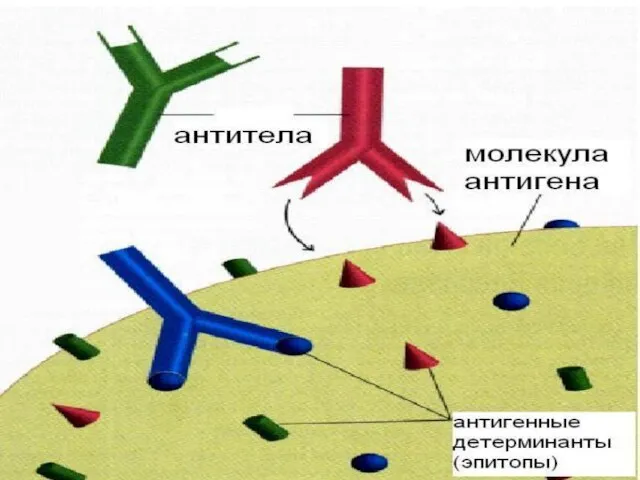
Слайд 83 Иммуногенность обусловлена сложностью пространственной структуры молекулы (боковые радикалы, разветвленные цепи и
Иммуногенность обусловлена сложностью пространственной структуры молекулы (боковые радикалы, разветвленные цепи и

Слайд 85 Гаптены - низкомолекулярные вещества, в обычных условиях не вызывают иммунную реакцию
Гаптены - низкомолекулярные вещества, в обычных условиях не вызывают иммунную реакцию

Слайд 86Аллергены - АГили гаптены, которые при повторном попадании в организм вызывают аллергическую
Аллергены - АГили гаптены, которые при повторном попадании в организм вызывают аллергическую

Слайд 87Т-зависимые АГ- запускают иммунный ответ с участием Т-лимфоцитов (белки)
Т-независимые АГ- запускают
Т-зависимые АГ- запускают иммунный ответ с участием Т-лимфоцитов (белки) Т-независимые АГ- запускают

Слайд 88 Экзогенные и эндогенные АГ
Экзогенные антигены попадают в организм из внешней
Экзогенные и эндогенные АГ Экзогенные антигены попадают в организм из внешней

Слайд 89Инфекционные АГ- антигены бактерий, вирусов, грибов, простейших, токсины, ферменты
⇒ группоспецифические (встречаются
Инфекционные АГ- антигены бактерий, вирусов, грибов, простейших, токсины, ферменты ⇒ группоспецифические (встречаются

Слайд 90По локализации: О-АГ Н-АГ К-АГ
О-АГ - полисахарид, входит в состав к
По локализации: О-АГ Н-АГ К-АГ О-АГ - полисахарид, входит в состав к

Слайд 91 Строение ЛПС
Центральная часть ЛПС – полисахарид, состоит из остатков 2-кето-3-дезоксиоктоната, галактозы,
Строение ЛПС Центральная часть ЛПС – полисахарид, состоит из остатков 2-кето-3-дезоксиоктоната, галактозы,

Слайд 92Липид А - неспецифический иммуностимулятор, токсичен
ЛПС – эндотоксин и пироген
В
Липид А - неспецифический иммуностимулятор, токсичен ЛПС – эндотоксин и пироген В

Слайд 93Н-АГ входит в состав жгутиков, состоит из белка флагеллина, термолабилен
К-АГ -
Н-АГ входит в состав жгутиков, состоит из белка флагеллина, термолабилен К-АГ -

Слайд 94Протективные антигены
Это совокупность антигенных детерминант (эпитопов), которые вызывают наиболее сильный иммунный
Протективные антигены Это совокупность антигенных детерминант (эпитопов), которые вызывают наиболее сильный иммунный

Слайд 95Антигенная мимикрия- сходство в строении АГ бактерий, человека, животных
Гемолитические стрептококки группы
Антигенная мимикрия- сходство в строении АГ бактерий, человека, животных Гемолитические стрептококки группы

Слайд 96У возбудителя сифилиса есть фосфолипиды, сходные по строению с кардиолипидным АГ сердца
У возбудителя сифилиса есть фосфолипиды, сходные по строению с кардиолипидным АГ сердца

Слайд 97Суперантигены - в небольших дозах вызывают поликлональную активацию и пролиферацию большого числа
Суперантигены - в небольших дозах вызывают поликлональную активацию и пролиферацию большого числа

Слайд 98НЕИНФЕКЦИОННЫЕ АНТИГЕНЫ
АГ растений, лекарственные препараты, химические, природные и синтетические вещества, антигены
НЕИНФЕКЦИОННЫЕ АНТИГЕНЫ АГ растений, лекарственные препараты, химические, природные и синтетические вещества, антигены

Слайд 99Антигены животных по отношению к человеку являются ксеногенными
Антигены, отличающие одного индивидуума
Антигены животных по отношению к человеку являются ксеногенными Антигены, отличающие одного индивидуума

Слайд 100 Антигены эритроцитов
На поверхности эритроцитов имеется более 100 антигенов, относящихся к
Антигены эритроцитов На поверхности эритроцитов имеется более 100 антигенов, относящихся к

Слайд 101 Антигены лейкоцитов
HLA (Human Leucocyte Antigens) контролируются генами 6 хромосомы (главным
Антигены лейкоцитов HLA (Human Leucocyte Antigens) контролируются генами 6 хромосомы (главным

Слайд 102 Молекулы HLA I класса – гетеродимеры, состоят из двух цепей Легкая
Молекулы HLA I класса – гетеродимеры, состоят из двух цепей Легкая

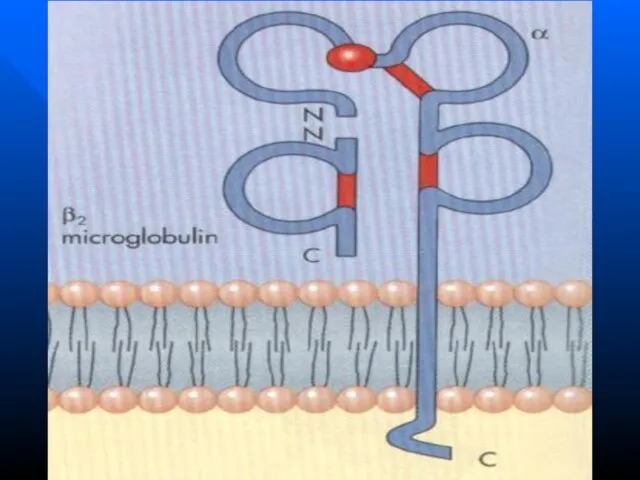
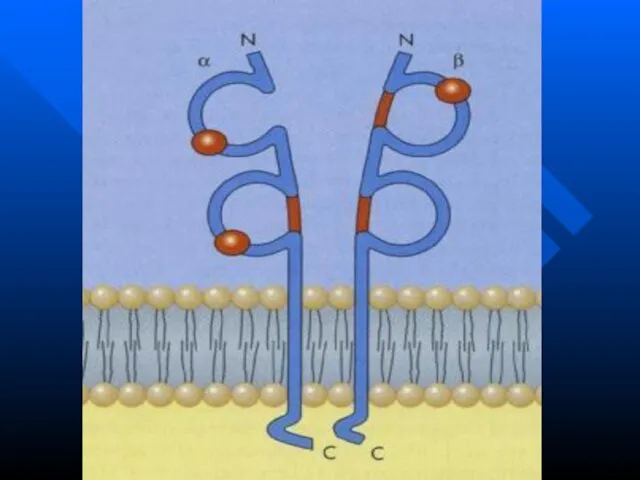
Слайд 104Молекулы HLA II класса состоят из двух полипептидных цепей: α и β.
Молекулы HLA II класса состоят из двух полипептидных цепей: α и β.

Слайд 106 Эндогенные антигены - собственные аутологичные молекулы (аутоантигены), вызывающие активацию системы иммунитета
Эндогенные антигены - собственные аутологичные молекулы (аутоантигены), вызывающие активацию системы иммунитета

Слайд 107
По тканевой и клеточной принадлежности - Стромальные (АГ эластических, коллагеновых и
По тканевой и клеточной принадлежности - Стромальные (АГ эластических, коллагеновых и

Слайд 108Система мононуклеарных фагоцитов
Моноциты крови
Оседлые макрофаги печени (клетки Купфера)
Система мононуклеарных фагоцитов
Моноциты крови
Оседлые макрофаги печени (клетки Купфера)

Слайд 109Молекулярные маркеры моноцитов и макрофагов
CD 14 – рецептор для ЛПС
Молекулярные маркеры моноцитов и макрофагов
CD 14 – рецептор для ЛПС

Слайд 110Функции макрофагов:
фагоцитоз распознавание и представление (презентация) антигенов секреция медиаторов системы □иммунитета
Функции макрофагов: фагоцитоз распознавание и представление (презентация) антигенов секреция медиаторов системы □иммунитета

Слайд 111Стадии фагоцитоза
хемотаксис
адгезия
эндоцитоз (поглощение)
переваривание
удаление продуктов фагоцитоза
Стадии фагоцитоза
хемотаксис
адгезия
эндоцитоз (поглощение)
переваривание
удаление продуктов фагоцитоза

Слайд 112Хемотаксис - движение макрофагов к объекту фагоцитоза по градиенту концентрации хемотаксических факторов
Хемотаксис - движение макрофагов к объекту фагоцитоза по градиенту концентрации хемотаксических факторов

Слайд 113Адгезия реализуется двумя механизмами: иммунным и неиммунным
Неиммунный фагоцитоз осуществляется за счет
Адгезия реализуется двумя механизмами: иммунным и неиммунным Неиммунный фагоцитоз осуществляется за счет

Слайд 114
В связывании могут участвовать самые разнообразные молекулы и рецепторы
Лектины микробных клеток
Лектины фагоцитарных
В связывании могут участвовать самые разнообразные молекулы и рецепторы Лектины микробных клеток Лектины фагоцитарных

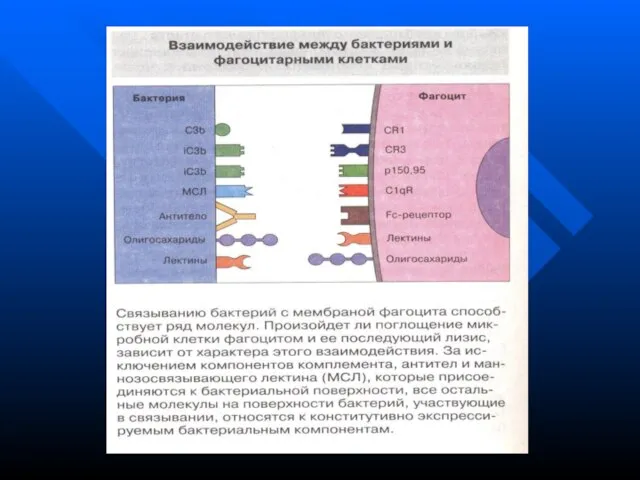
Слайд 116Эндоцитоз (поглощение)
инвагинация мембраны фагоцита
обволакивание объекта фагоцитоза
образование фагосомы
слияние фагосомы с
Эндоцитоз (поглощение) инвагинация мембраны фагоцита обволакивание объекта фагоцитоза образование фагосомы слияние фагосомы с

Слайд 117Переваривание
Кислородзависимые механизмы (образование активных форм кислорода: Н2О2, оксидаза, миелопероксидаза и др.)
Кислороднезависимые
Переваривание Кислородзависимые механизмы (образование активных форм кислорода: Н2О2, оксидаза, миелопероксидаза и др.) Кислороднезависимые

Слайд 118Кислородзависимые бактерицидные механизмы -
образование активных форм кислорода
(АФК) через гексозомонофосфатный шунт -
Кислородзависимые бактерицидные механизмы - образование активных форм кислорода (АФК) через гексозомонофосфатный шунт -
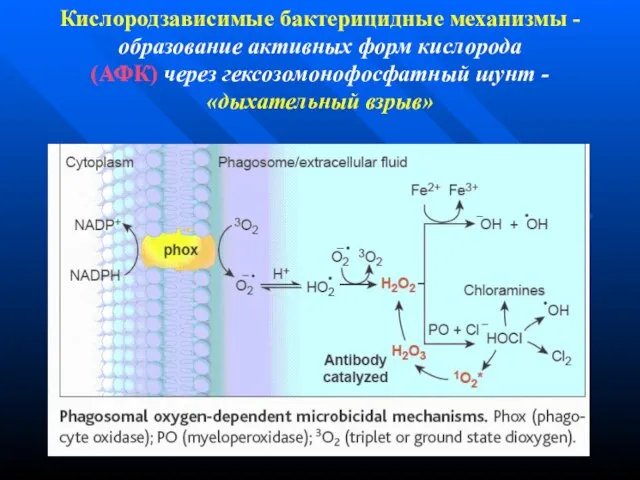
Слайд 119
Кислороднезависимые бактерицидные механизмы:
Ферменты-гидролазы - катепсины, лизоцим, глюкуронидаза и т.д.
Катионные антибиотикоподобные
Кислороднезависимые бактерицидные механизмы: Ферменты-гидролазы - катепсины, лизоцим, глюкуронидаза и т.д. Катионные антибиотикоподобные

Слайд 120Завершенный фагоцитоз - полное переваривание и гибель бактериальной клетки
Незавершенный фагоцитоз -
Завершенный фагоцитоз - полное переваривание и гибель бактериальной клетки Незавершенный фагоцитоз -

Слайд 121Представление (презентация) антигенов макрофагами
В результате фагоцитоза и переваривания антигенов образуется большое
Представление (презентация) антигенов макрофагами В результате фагоцитоза и переваривания антигенов образуется большое

Слайд 122Экзоантигены - пептиды длиной 12-25 аминокислот связываются с HLA 2 класса (HLA-DR,
Экзоантигены - пептиды длиной 12-25 аминокислот связываются с HLA 2 класса (HLA-DR,

Слайд 123Секреция медиаторов иммунной системы (монокинов).
Второй сигнал для активации Т-хелперов - выделение
Секреция медиаторов иммунной системы (монокинов). Второй сигнал для активации Т-хелперов - выделение

Слайд 124ДРУГИЕ АНТИГЕН-ПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
(АПК)
Клетки Лангерганса из кожи мигрируют в паракортикальные области регионарных лимфоузлов.
ДРУГИЕ АНТИГЕН-ПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ
(АПК)
Клетки Лангерганса из кожи мигрируют в паракортикальные области регионарных лимфоузлов.

Слайд 126СИСТЕМА ГРАНУЛОЦИТОВ
В нее входят нейтрофильные, базофильные и эозинофильные гранулоциты (микрофаги). Все
СИСТЕМА ГРАНУЛОЦИТОВ В нее входят нейтрофильные, базофильные и эозинофильные гранулоциты (микрофаги). Все

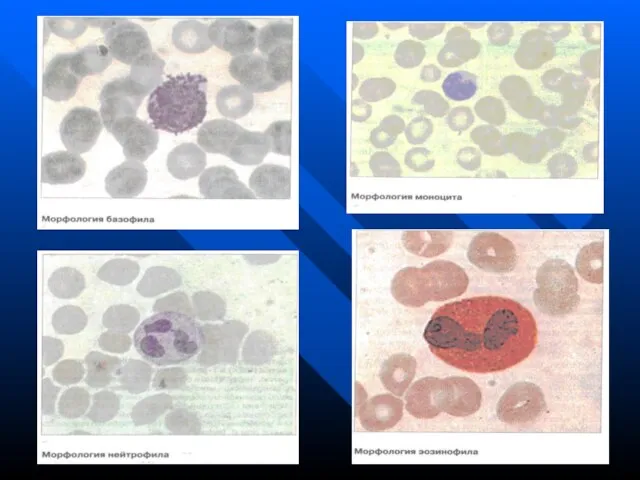
Слайд 128Нейтрофилы
составляют 47-72% всех лейкоцитов. Их содержится 2.5 - 4.5х109/л. Средний срок
Нейтрофилы составляют 47-72% всех лейкоцитов. Их содержится 2.5 - 4.5х109/л. Средний срок

Слайд 129Азурофильные гранулы содержат β-глюкуронидазу, катепсины, кислые и нейтральные протеиназы, миелопероксидазу.
В специфических
Азурофильные гранулы содержат β-глюкуронидазу, катепсины, кислые и нейтральные протеиназы, миелопероксидазу. В специфических

Слайд 131Основные молекулы-маркеры на нейтрофилах : CD13,СД 14, СD64 , рецепторы к С1q,
Основные молекулы-маркеры на нейтрофилах : CD13,СД 14, СD64 , рецепторы к С1q,

Слайд 132Базофилы
участвуют в аллергических реакциях. На поверхности базофилов имеются Fcε-рецепторы, связывающие IgE.
Базофилы участвуют в аллергических реакциях. На поверхности базофилов имеются Fcε-рецепторы, связывающие IgE.

Слайд 133Эозинофилы
играют большую роль в противопаразитарном иммунитете и аллергии. В крови 1-6%,
Эозинофилы играют большую роль в противопаразитарном иммунитете и аллергии. В крови 1-6%,

Слайд 134Характеристика системы макрофагов и гранулоцитов
1. Определяют количество лейкоцитов в крови.
2. Оценивают поглотительную
Характеристика системы макрофагов и гранулоцитов 1. Определяют количество лейкоцитов в крови. 2. Оценивают поглотительную

Слайд 135Определяют фагоцитарный индекс и фагоцитарное число
Фагоцитарный индекс - это среднее количество
Определяют фагоцитарный индекс и фагоцитарное число Фагоцитарный индекс - это среднее количество

Слайд 136Оценка показателей фагоцитоза
В норме через 90 мин фагоцитарный индекс должен быть ниже,
Оценка показателей фагоцитоза В норме через 90 мин фагоцитарный индекс должен быть ниже,

Слайд 137НСТ-тест
Метаболическую активность - «дыхательный взрыв» определяют после окраски лейкоцитов раствором нитросинего
НСТ-тест Метаболическую активность - «дыхательный взрыв» определяют после окраски лейкоцитов раствором нитросинего

Слайд 138
С помощью моноклональных антител определяют антигены фагоцитов (СD14, СD11, СD18, HLA-DR и
С помощью моноклональных антител определяют антигены фагоцитов (СD14, СD11, СD18, HLA-DR и

Слайд 139Система комплемента
сложная система ферментативных и рецепторных белков сыворотки крови (более 20)
Активация
Система комплемента
сложная система ферментативных и рецепторных белков сыворотки крови (более 20)
Активация

Слайд 140Белки классического пути активации системы комплемента называются компонентами и обозначаются буквой C

Слайд 141При активации компонентов комплемента обычно образуется 2 фрагмента
Больший фрагмент обозначается малой
Больший фрагмент обозначается малой

Слайд 142Среди регуляторных белков различают естественный С1-ингибитор, который тормозит спонтанную активацию C1q компонента

Слайд 143Пути активации системы комплемента
Классический (запускается комплексом АГ-АТ в присутствии катионов Ca и
Пути активации системы комплемента
Классический (запускается комплексом АГ-АТ в присутствии катионов Ca и

Слайд 148Функции системы комплемента
Лизис клеток-мишеней (бактериальных, зараженных вирусом, опухолевых)
Опсонизация, т.е. усиление фагоцитоза через
Функции системы комплемента
Лизис клеток-мишеней (бактериальных, зараженных вирусом, опухолевых)
Опсонизация, т.е. усиление фагоцитоза через

 Атомно-абсорбционная спектроскопия
Атомно-абсорбционная спектроскопия История возникновения числа
История возникновения числа НДФЛ. Заработная плата (оплата труда работника)
НДФЛ. Заработная плата (оплата труда работника) Презентация на тему Русский интуитивизм
Презентация на тему Русский интуитивизм Лекция 3
Лекция 3 Системный блок
Системный блок Вклад ученых-физиков в дело Великой Победы
Вклад ученых-физиков в дело Великой Победы От детства к зрелости. Воспитание ребенка в семье
От детства к зрелости. Воспитание ребенка в семье Классификация цепей поставок и их краткая характеристика Ненадышина Т., Губанова М.
Классификация цепей поставок и их краткая характеристика Ненадышина Т., Губанова М. Изучение готовности первоклассников к обучению в школе
Изучение готовности первоклассников к обучению в школе Is it good to be young
Is it good to be young  О едином подходе к формированию ОК-компетенций в области Техники и технологии
О едином подходе к формированию ОК-компетенций в области Техники и технологии Презентация на тему Грибы - двойники
Презентация на тему Грибы - двойники безопасность
безопасность Счастливые отношения в семье. Вводный вебинар
Счастливые отношения в семье. Вводный вебинар ОГП тема 3
ОГП тема 3 Презентация на тему Тип Кольчатые черви
Презентация на тему Тип Кольчатые черви Информационный проект нового поколения
Информационный проект нового поколения «Национальное бесконечно богато…» Д.С. Лихачёв
«Национальное бесконечно богато…» Д.С. Лихачёв Школьная ассоциация нового самоуправления
Школьная ассоциация нового самоуправления Тест по теме: «Столетняя война»
Тест по теме: «Столетняя война» Продаем 3-комнатную квартиру в Омске
Продаем 3-комнатную квартиру в Омске Печать автомобильных деталей на 3D-принтере
Печать автомобильных деталей на 3D-принтере Привлечение финансирования через ЗПИФ
Привлечение финансирования через ЗПИФ Проблемные кредиты – итальянский опыт
Проблемные кредиты – итальянский опыт Как выбрать инструмент для решения проблемы
Как выбрать инструмент для решения проблемы Презентация на тему Развивающее обучение что это такое
Презентация на тему Развивающее обучение что это такое  Предпринимательство и предприниматель
Предпринимательство и предприниматель