- Введение в организацию белковой структуры и молекулярное моделирование

Содержание
- 2. Проблематика самоорганизации структуры белка “Предсказание трехмерной структуры белка по его аминокислотной последовательности.” “Каким образом белковая молекула
- 3. Белки могут сворачиваться в свои третичные структуры спонтанно В растворах белковые молекулы самостоятельно организуются в трехмерные
- 4. Парадокс Левинтала Можем предположить, что для каждой аминокислоты существует три варианта ее состояния в белке (α-спираль,
- 5. Почему “Самоорганизация структуры белка” так важна? Белки играют важные роли в живых организмах. Некоторые белки напрямую
- 6. Почему проблема “Самоорганизации структуры белка” так сложна? С точки зрения компьютерного моделирования, Сложно моделировать весь процесс
- 7. Молекулярная Динамика (MD) В модели молекулярной динамики, мы имитируем движение атомов как функцию от времени на
- 8. Интегрирование с использованием метода конечной разности Положения в моменты времени (t + Δt ) и (t
- 9. Силы, участвующие в процессе самоорганизации белковой молекулы Электростатические взаимодействия Силы Ван дер Ваальса Водородные связи Гидрофобные
- 10. Функции от составляющих сил, рассчитываемые в Молекулярном Моделировании Электростатическая сила Водородная связь Сила Ван дер Ваальса
- 11. Система для Моделирования с помощью Молекулярной Динамики Без молекул воды С молекулами воды Кол-во атомов: 304
- 12. MD требует колоссальных вычислительных мощностей Квант времени в методе MD (Δt) ограничен 1 фсек (10-15 сек).
- 13. Шкала времени изменения состояний белка и MD Время Сжатие связи Прохождение иона по ионному каналу Упругие
- 14. Гораздо быстрее, Гораздо больше! Специальные компьютеры Расчет свободных взаимодействий (не химических связей) обеспечивается с использованием специальных
- 15. Броуновская Динамика (BD) Динамический вклад растворителя представлен в виде рассеянного случайного воздействия (Открытие Эйнштейна в 1905).
- 16. Система для BD Моделирования Без молекул воды С молекулами воды Кол-во атомов: 304 Кол-во атомов: 304
- 17. Алгоритм BD Уравнение Ланжевина может быть выражено так: Здесь, ri и mi отражают соответственно положение и
- 18. Вычислительное время BD †MTS(Multiple time step) алгоритм: Этот метод позволяет уменьшить необходимость расчетов самой тяжелой части
- 19. Моделирование образования α-спирали с помощью BD Доля возможных в природе связей Время расчета (нсек) 0 300
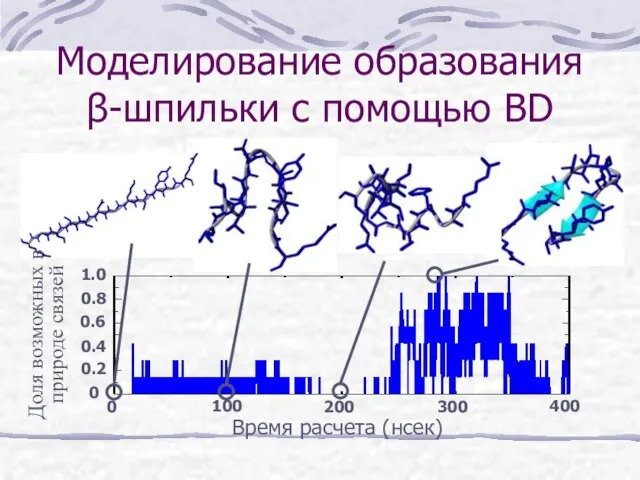
- 20. Моделирование образования β-шпильки с помощью BD Доля возможных в природе связей Время расчета (нсек) 0 300
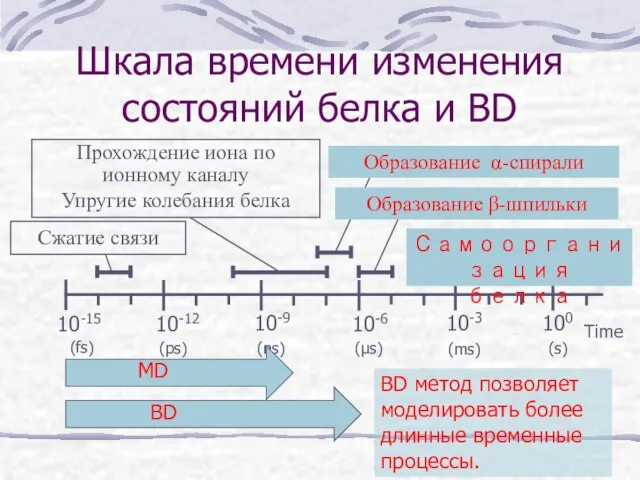
- 21. Шкала времени изменения состояний белка и BD Time BD метод позволяет моделировать более длинные временные процессы.
- 23. Скачать презентацию
Слайд 2Проблематика самоорганизации структуры белка
“Предсказание трехмерной структуры белка по его аминокислотной последовательности.”
“Каким образом
Проблематика самоорганизации структуры белка
“Предсказание трехмерной структуры белка по его аминокислотной последовательности.”
“Каким образом

Слайд 3Белки могут сворачиваться в свои третичные структуры спонтанно
В растворах белковые молекулы самостоятельно
Белки могут сворачиваться в свои третичные структуры спонтанно
В растворах белковые молекулы самостоятельно

Слайд 4Парадокс Левинтала
Можем предположить, что для каждой аминокислоты существует три варианта ее состояния
Парадокс Левинтала
Можем предположить, что для каждой аминокислоты существует три варианта ее состояния

Слайд 5Почему “Самоорганизация структуры белка” так важна?
Белки играют важные роли в живых организмах.
Некоторые
Почему “Самоорганизация структуры белка” так важна?
Белки играют важные роли в живых организмах.
Некоторые

Слайд 6Почему проблема “Самоорганизации структуры белка” так сложна?
С точки зрения компьютерного моделирования,
Сложно моделировать
Почему проблема “Самоорганизации структуры белка” так сложна?
С точки зрения компьютерного моделирования,
Сложно моделировать

Слайд 7Молекулярная Динамика (MD)
В модели молекулярной динамики, мы имитируем движение атомов как функцию
Молекулярная Динамика (MD)
В модели молекулярной динамики, мы имитируем движение атомов как функцию

Слайд 8Интегрирование с использованием метода конечной разности
Положения в моменты времени (t + Δt
Интегрирование с использованием метода конечной разности
Положения в моменты времени (t + Δt

Слайд 9Силы, участвующие в процессе самоорганизации белковой молекулы
Электростатические взаимодействия
Силы Ван дер Ваальса
Водородные связи
Гидрофобные
Силы, участвующие в процессе самоорганизации белковой молекулы
Электростатические взаимодействия
Силы Ван дер Ваальса
Водородные связи
Гидрофобные

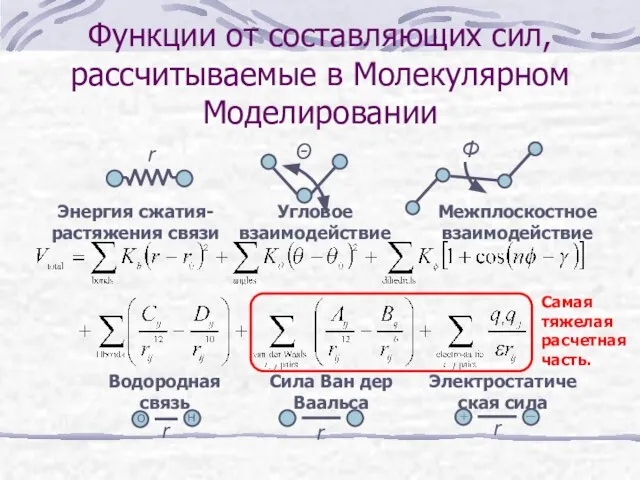
Слайд 10Функции от составляющих сил, рассчитываемые в Молекулярном Моделировании
Электростатическая сила
Водородная связь
Сила Ван дер
Функции от составляющих сил, рассчитываемые в Молекулярном Моделировании
Электростатическая сила
Водородная связь
Сила Ван дер
Слайд 11Система для Моделирования с помощью Молекулярной Динамики
Без молекул воды
С молекулами воды
Кол-во атомов:
Система для Моделирования с помощью Молекулярной Динамики
Без молекул воды
С молекулами воды
Кол-во атомов:

Слайд 12MD требует колоссальных вычислительных мощностей
Квант времени в методе MD (Δt) ограничен 1
MD требует колоссальных вычислительных мощностей
Квант времени в методе MD (Δt) ограничен 1

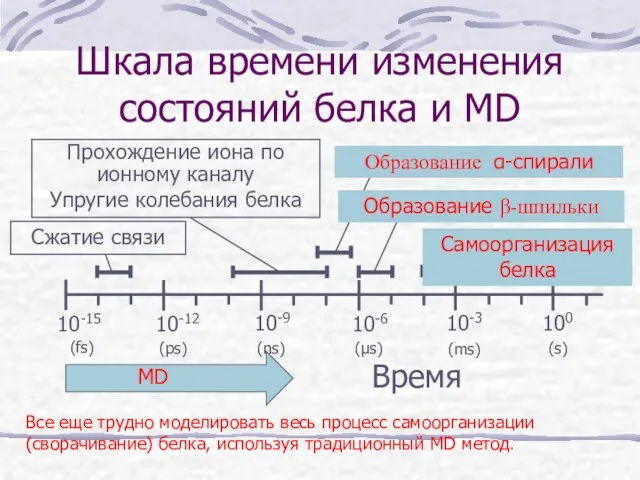
Слайд 13Шкала времени изменения состояний белка и MD
Время
Сжатие связи
Прохождение иона по ионному каналу
Упругие
Шкала времени изменения состояний белка и MD
Время
Сжатие связи
Прохождение иона по ионному каналу
Упругие
Слайд 14Гораздо быстрее, Гораздо больше!
Специальные компьютеры
Расчет свободных взаимодействий (не химических связей) обеспечивается с
Гораздо быстрее, Гораздо больше!
Специальные компьютеры
Расчет свободных взаимодействий (не химических связей) обеспечивается с

Слайд 15Броуновская Динамика (BD)
Динамический вклад растворителя представлен в виде рассеянного случайного воздействия (Открытие
Броуновская Динамика (BD)
Динамический вклад растворителя представлен в виде рассеянного случайного воздействия (Открытие

Слайд 16Система для BD Моделирования
Без молекул воды
С молекулами воды
Кол-во атомов: 304
Кол-во атомов: 304 +
Система для BD Моделирования
Без молекул воды
С молекулами воды
Кол-во атомов: 304
Кол-во атомов: 304 +

Слайд 17Алгоритм BD
Уравнение Ланжевина может быть выражено так:
Здесь, ri и mi отражают соответственно
Алгоритм BD
Уравнение Ланжевина может быть выражено так: Здесь, ri и mi отражают соответственно

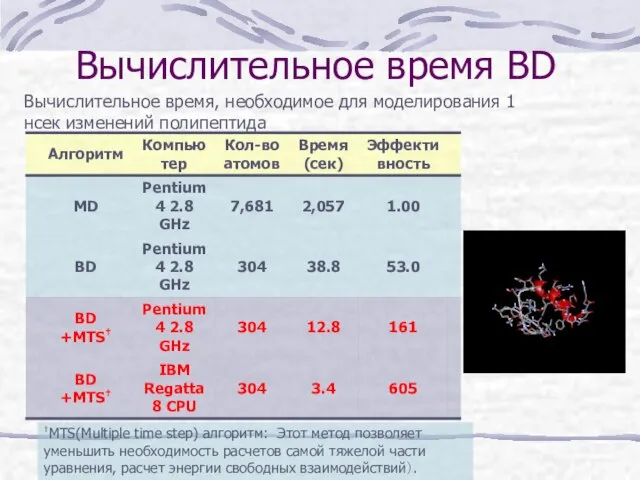
Слайд 18Вычислительное время BD
†MTS(Multiple time step) алгоритм: Этот метод позволяет уменьшить необходимость расчетов самой
Вычислительное время BD
†MTS(Multiple time step) алгоритм: Этот метод позволяет уменьшить необходимость расчетов самой
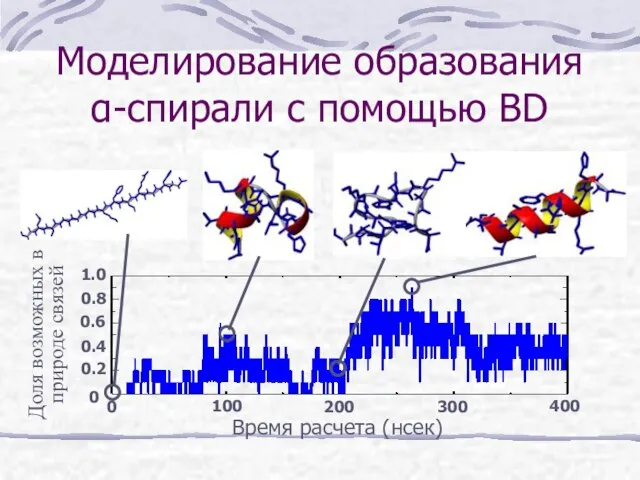
Слайд 19Моделирование образования α-спирали с помощью BD
Доля возможных в природе связей
Время расчета (нсек)
0
300
200
100
400
0
0.2
0.4
0.6
0.8
1.0
Моделирование образования α-спирали с помощью BD
Доля возможных в природе связей
Время расчета (нсек)
0
300
200
100
400
0
0.2
0.4
0.6
0.8
1.0
Слайд 20Моделирование образования
β-шпильки с помощью BD
Доля возможных в природе связей
Время расчета (нсек)
0
300
200
100
400
0
0.2
0.4
0.6
0.8
1.0
Моделирование образования
β-шпильки с помощью BD
Доля возможных в природе связей
Время расчета (нсек)
0
300
200
100
400
0
0.2
0.4
0.6
0.8
1.0
Слайд 21Шкала времени изменения состояний белка и BD
Time
BD метод позволяет моделировать более длинные
Шкала времени изменения состояний белка и BD
Time
BD метод позволяет моделировать более длинные
 Элементы росписи оятской керамики. Часть 2
Элементы росписи оятской керамики. Часть 2 КРАТКАЯ ИСТОРИЯ РАЗВИТИЯВЫЧИСЛИТЕЛЬНОЙ ТЕХНИКИ
КРАТКАЯ ИСТОРИЯ РАЗВИТИЯВЫЧИСЛИТЕЛЬНОЙ ТЕХНИКИ Отделка вышиванием по низу прямой юбки используя мотивы горных мари.
Отделка вышиванием по низу прямой юбки используя мотивы горных мари. The Amazing world of English - Speaking Countries
The Amazing world of English - Speaking Countries Предложение по оптимизации печатной ифраструктуры
Предложение по оптимизации печатной ифраструктуры Digital-маркетинг.Клиенты из интернетаот стартадо повторных продаж
Digital-маркетинг.Клиенты из интернетаот стартадо повторных продаж Тайна как инструмент изучения бессознательного в культурно-историческом подходе
Тайна как инструмент изучения бессознательного в культурно-историческом подходе Опыт Казахстанав обработке дефолтовлистинговых компанийАндрей ЦалюкВице-президентАО "Казахстанская фондовая биржа"специал
Опыт Казахстанав обработке дефолтовлистинговых компанийАндрей ЦалюкВице-президентАО "Казахстанская фондовая биржа"специал Газові види пального їх сьогодення й майбутнє
Газові види пального їх сьогодення й майбутнє Автоперевозки
Автоперевозки  Кто хочет стать миллионеромСтихотворение «Железная дорога»
Кто хочет стать миллионеромСтихотворение «Железная дорога» Хорошая Карма Предстоящее чтение будет приятным, но непродолжительным. Насладись им! Это то, что сказал Далай Лама в преддверии 2009
Хорошая Карма Предстоящее чтение будет приятным, но непродолжительным. Насладись им! Это то, что сказал Далай Лама в преддверии 2009 Презентация на тему Средняя линия трапеции
Презентация на тему Средняя линия трапеции Презентация на тему Александр Исаевич Солженицин политик и патриот
Презентация на тему Александр Исаевич Солженицин политик и патриот Светская этика
Светская этика Активные и пассивные потребители
Активные и пассивные потребители Экономические школы
Экономические школы техногенна небезпека
техногенна небезпека Показательная функция
Показательная функция Презентація ІІУС_3
Презентація ІІУС_3 Ф О Т О Э Ф Ф Е К Т
Ф О Т О Э Ф Ф Е К Т Э-74 Против курения
Э-74 Против курения Какие бывают животные? 2 класс
Какие бывают животные? 2 класс Нефть – не топливо, топить можно и ассигнациями…
Нефть – не топливо, топить можно и ассигнациями… Мировые религии
Мировые религии New York City
New York City Cправочная Cлужба«Нужные вещи 393000» www.393000.ru
Cправочная Cлужба«Нужные вещи 393000» www.393000.ru Культура славян
Культура славян