- Эукариоттардағы РНҚ процессинг

Содержание
- 2. Бактериялық мРНҚ молекуласы геномның белгілі бір жерлерінде РНҚ-полимеразаның тоқтауымен немесе қайта іске қосылуы арқылы синтезделеді. Эукариоттарда
- 3. Эукариоттар мен бактериядағы «геннен белокқа дейінгі» жол сатыларының қысқаша сипаты
- 4. Эукариоттарда мРНҚ молекуласының екі ұшы да модификацияланады: 5‘-ұшы кэпирленеді, ал 3‘-ұшы полиаденилденеді. Бұндай арнайы ұштар бұл
- 5. Прокариот және эукариоттардағы мРНҚ молекуласының құрылымын салыстыру
- 6. РНҚ полимераза II арқылы транскрипция инициациясының маңызды сатысы – CTD (С-ұшы домені) деп аталған РНҚ полимераза
- 7. Эукариоттардағы РНҚ полимераза II «РНҚ зауыты» сияқты CTD 7-амин қышқылдық тізбектің 52 тандемді қайталамасынан тұрады; Әр
- 8. РНҚ полимераза II шамамен РНҚ-ның 25 нуклеотидін түзгеннен кейін РНҚ-ның жаңа молекуласының 5‘-ұшында гуаниннің модификацияланған нуклеотиді
- 9. РНҚ полимераза II арқылы синтезделетін әр РНҚ молекуласының 5´-ұшында кэп құрылымның түзілуіне әкелетін реакциялар
- 10. Эукариот гендері ұсақ бөлшектерге бөлінген: кодтаушы тізбектер – экспрессиялаушы тізбектер немесе экзондар, - ұзындау тізбектер -қосындылар
- 11. Экзон мен интронның өзара орналасуын көрсететін адамның екі генінің құрылымы
- 12. Пре-мРНҚ-ның сплайсинг реакциясы
- 13. Егеуқұйрықтың α-тропомиозин генінің альтернативті сплайсингі Көптеген гендердің сплайсингі бір емес, бірнеше әдістер арқылы жүреді, ол бір
- 14. Сплайсингтік кешен РНҚ-ізашарлары молекуласының үш бөлігін таниды: сплайсингтің 5‘-сайтын, сплайсинтің 3‘-сайтын және кесілген арқанның негізін қалыптастыратын
- 15. Адам геномындағы көптеген интрондардың «басталуы» мен «аяқталуын» сигнализациялайтын РНҚ молекуласындағы нуклеотидтердің консенсустық тізбектері
- 16. РНҚ сплайсингінің негізгі сатыларын белоктар емес РНҚ молекулалары атқарады. Маманданған РНҚ молекулалары қай жерде сплайсинг жүру
- 17. Пре-мРНҚ сплайсингінің механизмі
- 18. Сплайсинг реакциясының басталуы кезінде сплайсингтің 5 ‘ –шекарасын тану, тармақталу нүктесінің участогын тану және сплайсингтің 3
- 19. Пре-мРНҚ сплайсингі кезінде болатын кейбір қайта түзілулер
- 20. Транскрипция басталарда РНҚ полимеразаның фосфорланған ұшы сплайсоманың бірнеше компоненттерін алып жүреді және бұл компоненттер тікелей полимеразадан
- 21. Сплайсинг қателігінің екі түрі
- 22. Адам, шыбын, құрт геномындағы экзон және интрон ұзындығының ауытқуы
- 23. Экзонды анықтау концепциясы
- 24. Экзон және интрон шекарасын таңбалау, сонымен қатар сплайсоманы жинақтау РНҚ молекуласында, яғни РНҚ полимераза әлі де
- 25. Ашытқы сияқты қарапайым эукариоттар тек snРНП –дің бір жинағынан тұрады, алайда шыбын, сүтқоректілер, өсімдік типіндегі күрделі
- 26. Трипаносомдарда ( адамдарда африкалық ұйқы ауруын тудыратын қарапайым жануарлар) және нематодтарда сплайсингтің ерекше түрі анықталған, оны
- 27. РНҚ сплайсингтің үш типінде қолданылатын механизмнің жалпы схемасы
- 28. β-талассемиямен ауыратын адамдардағы β-глобиннің біріншілік РНҚ транскриптінің дұрыс емес процессингі
- 29. Эволюция кезінде жасушалар басты катализатор ретінде белоктарды емес, РНҚ молекуласын пайдалануы мүмкін және өзінің генетикалық ақпаратын
- 30. Интронның өздігінен сплайсингтелетін тізбегінің екі танымал классы
- 31. мРНҚ-ның әр молекуласындағы 3‘-ұшының позициясы геномда кодталған сигнал арқылы анықталады. Бұндай сигналдар РНҚ-да транскрипцияланады және РНҚ
- 32. Эукариоттық аРНҚ-ның 3 ‘ –ұшының түзілуі кезіндегі ыдырау мен полиаденилдеуді бағыттайтын консенсустық нуклеотидтік тізбектер
- 33. Эукариоттардың мРНҚ 3 ‘ –ұшының түзілуі кезіндегі басты кезеңдерінің кейібіреулері
- 34. Эукариоттың пре-мРНҚ-сының 3 ‘ –ұшы ыдырағаннан кейін РНҚ полимеразаII транскрипцияны бастайды, бірақ біраздан соң матрицаны қоршап
- 35. Сәтті өңделген мРНҚ молекуласы нуклеоплазма мен цитозольды байланыстыратын ядролық кеуекті кешен арқылы өтеді. Макромолекулалар ядролық немесе
- 36. Ірі мРНҚ молекуласының ядролық кеуектік кешен арқылы тасымалдануы
- 37. Экспортталуға дайын мРНҚ молекуласының және оның ядролық кеуек арқылы тасымалының схемалық суреті
- 38. Эукариоттарда РРНҚ-ның төрт типі бар, олардың әрқайсысы бір рибосомаға ұқсас көшірме түрінде кездеседі. Төрт типтің үшеуі
- 39. Электрондық микроскоп арқылы бақыланған рРНҚ –ның тандемді орналасқан генінің транскрипциясы
- 40. Кең ауқымды химиялық модификациялар 13000-нуклеотидтік рРНҚ ізашарында жүреді,яғни одан рРНҚ молекулалары кесіліп тасталмағанға және рибосомаға жинақталғанға
- 41. Үш рибосомалық РНҚ-ның түзілуі арқылы эукариоттың рРНҚ-сының 45 S ізашары молекуласының нуклеотидтік процессингі және химиялық модификациясы.
- 42. Бағыттаушы РНҚ арқылы рРНҚ ізашарының модификациясы
- 43. Адам фибробластындағы ядрошықтың жұқа кесіндісінің электрондық микрофотографиясы; ядрошықта үш әртүрлі зона көрінген
- 44. Адам жасушасында ядрошықтың сыртқы түрі жасушалық циклдың әр сатысында өзгеріске ұшырап отырады
- 45. Ядрошықтың бірігуі
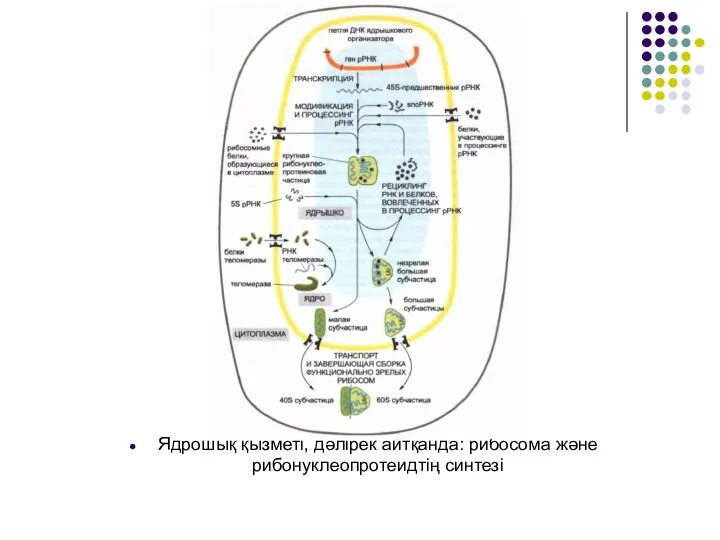
- 46. Ядрошық қызметі, дәлірек айтқанда: рибосома және рибонуклеопротеидтің синтезі
- 48. Скачать презентацию
Слайд 2Бактериялық мРНҚ молекуласы геномның белгілі бір жерлерінде РНҚ-полимеразаның тоқтауымен немесе қайта іске
Бактериялық мРНҚ молекуласы геномның белгілі бір жерлерінде РНҚ-полимеразаның тоқтауымен немесе қайта іске

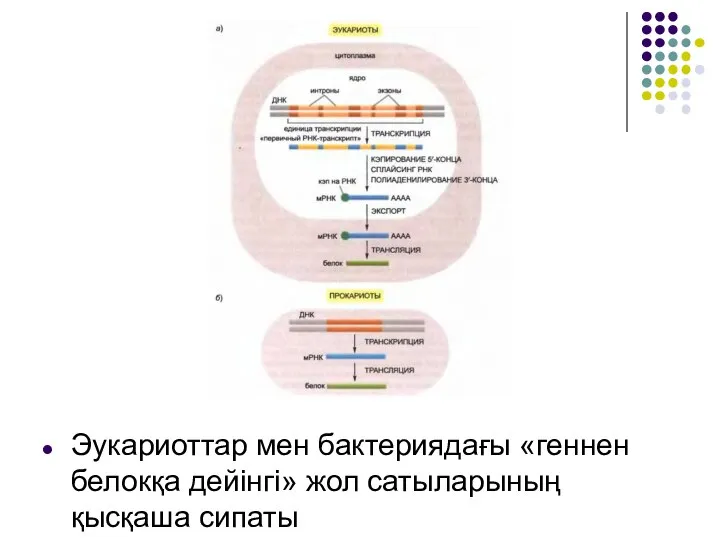
Слайд 3Эукариоттар мен бактериядағы «геннен белокқа дейінгі» жол сатыларының қысқаша сипаты
Эукариоттар мен бактериядағы «геннен белокқа дейінгі» жол сатыларының қысқаша сипаты
Слайд 4Эукариоттарда мРНҚ молекуласының екі ұшы да модификацияланады: 5‘-ұшы кэпирленеді, ал 3‘-ұшы полиаденилденеді.
Бұндай
Эукариоттарда мРНҚ молекуласының екі ұшы да модификацияланады: 5‘-ұшы кэпирленеді, ал 3‘-ұшы полиаденилденеді.
Бұндай

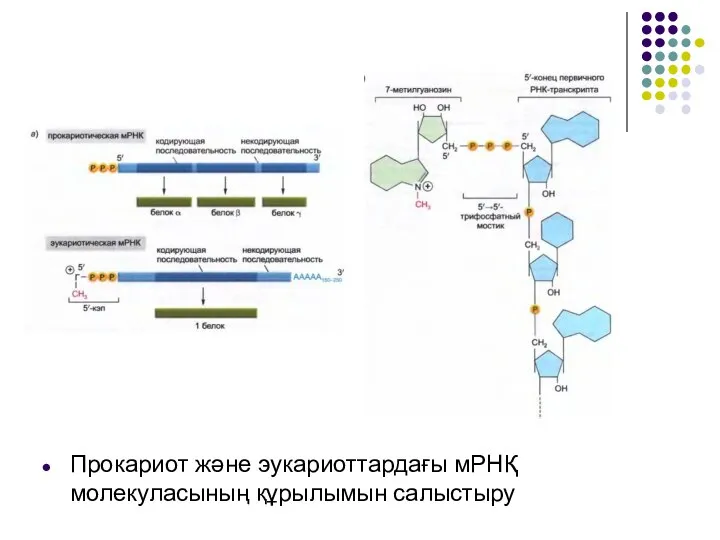
Слайд 5Прокариот және эукариоттардағы мРНҚ молекуласының құрылымын салыстыру
Прокариот және эукариоттардағы мРНҚ молекуласының құрылымын салыстыру
Слайд 6РНҚ полимераза II арқылы транскрипция инициациясының маңызды сатысы – CTD (С-ұшы домені)
РНҚ полимераза II арқылы транскрипция инициациясының маңызды сатысы – CTD (С-ұшы домені)

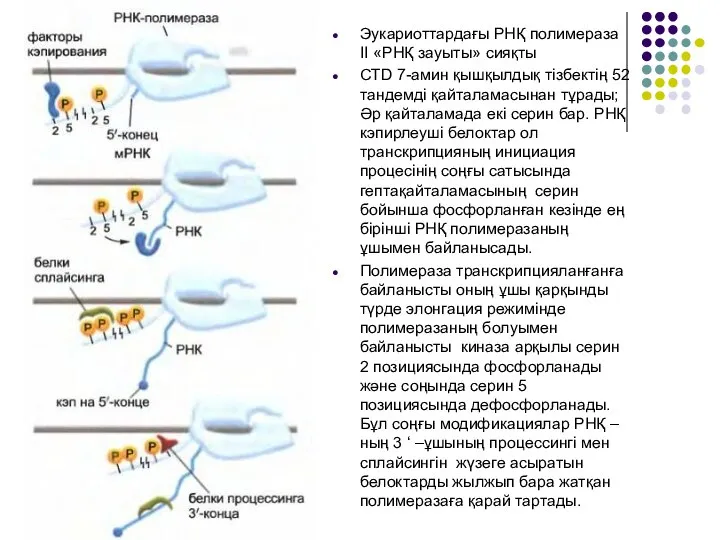
Слайд 7Эукариоттардағы РНҚ полимераза II «РНҚ зауыты» сияқты
CTD 7-амин қышқылдық тізбектің 52 тандемді
Эукариоттардағы РНҚ полимераза II «РНҚ зауыты» сияқты
CTD 7-амин қышқылдық тізбектің 52 тандемді
Слайд 8РНҚ полимераза II шамамен РНҚ-ның 25 нуклеотидін түзгеннен кейін РНҚ-ның жаңа молекуласының
РНҚ полимераза II шамамен РНҚ-ның 25 нуклеотидін түзгеннен кейін РНҚ-ның жаңа молекуласының

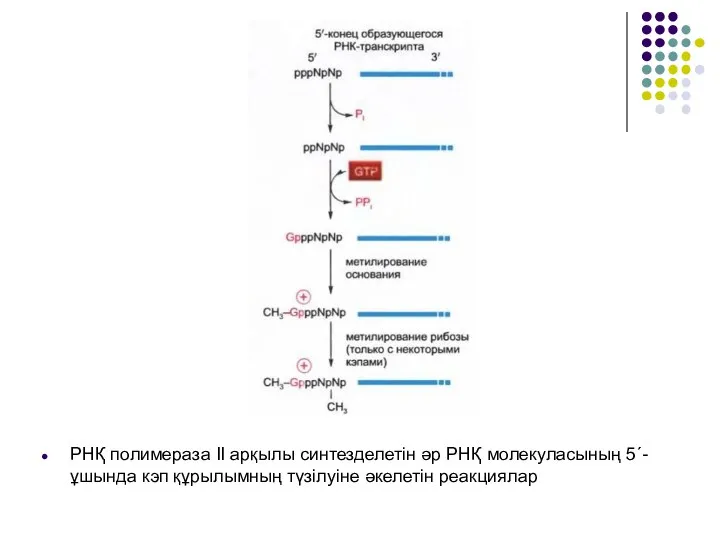
Слайд 9РНҚ полимераза II арқылы синтезделетін әр РНҚ молекуласының 5´-ұшында кэп құрылымның түзілуіне
РНҚ полимераза II арқылы синтезделетін әр РНҚ молекуласының 5´-ұшында кэп құрылымның түзілуіне
Слайд 10Эукариот гендері ұсақ бөлшектерге бөлінген: кодтаушы тізбектер – экспрессиялаушы тізбектер немесе экзондар,
Эукариот гендері ұсақ бөлшектерге бөлінген: кодтаушы тізбектер – экспрессиялаушы тізбектер немесе экзондар,

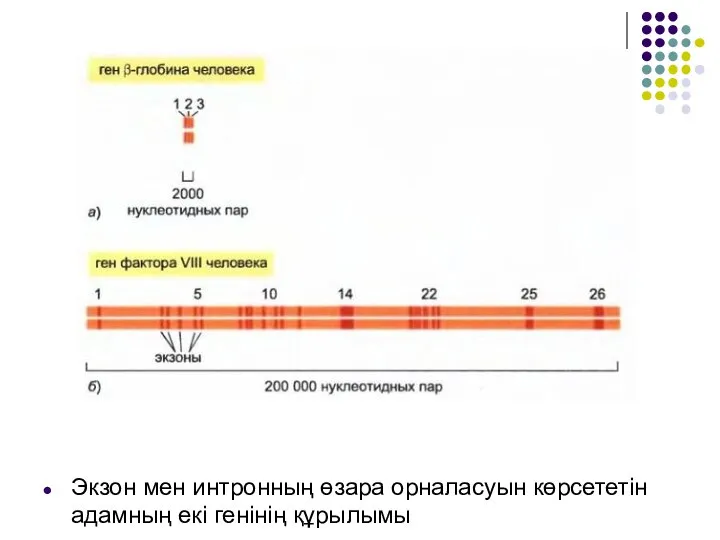
Слайд 11Экзон мен интронның өзара орналасуын көрсететін адамның екі генінің құрылымы
Экзон мен интронның өзара орналасуын көрсететін адамның екі генінің құрылымы
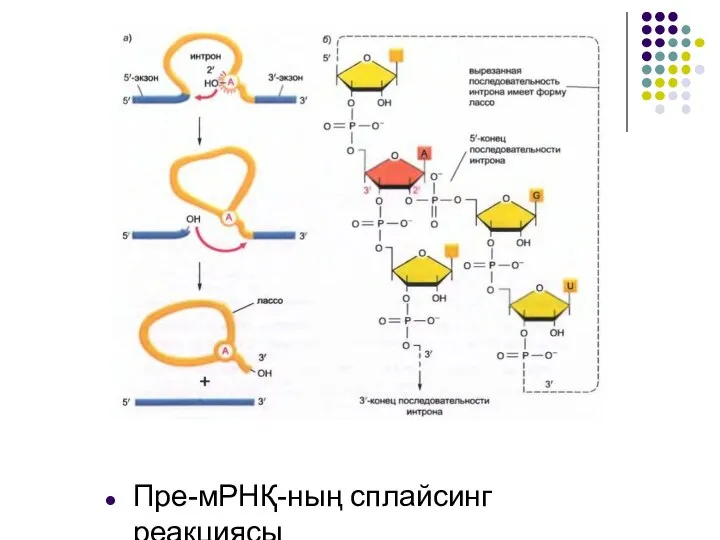
Слайд 12Пре-мРНҚ-ның сплайсинг реакциясы
Пре-мРНҚ-ның сплайсинг реакциясы
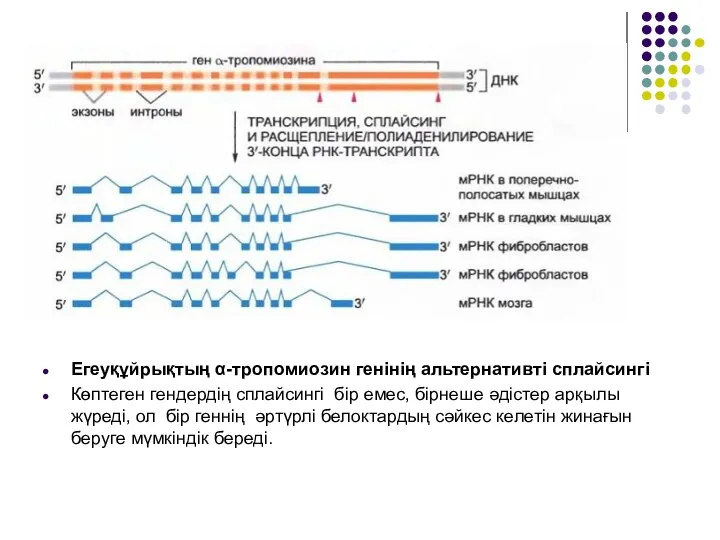
Слайд 13Егеуқұйрықтың α-тропомиозин генінің альтернативті сплайсингі
Көптеген гендердің сплайсингі бір емес, бірнеше әдістер арқылы
Егеуқұйрықтың α-тропомиозин генінің альтернативті сплайсингі
Көптеген гендердің сплайсингі бір емес, бірнеше әдістер арқылы
Слайд 14Сплайсингтік кешен РНҚ-ізашарлары молекуласының үш бөлігін таниды: сплайсингтің 5‘-сайтын, сплайсинтің 3‘-сайтын және
Сплайсингтік кешен РНҚ-ізашарлары молекуласының үш бөлігін таниды: сплайсингтің 5‘-сайтын, сплайсинтің 3‘-сайтын және

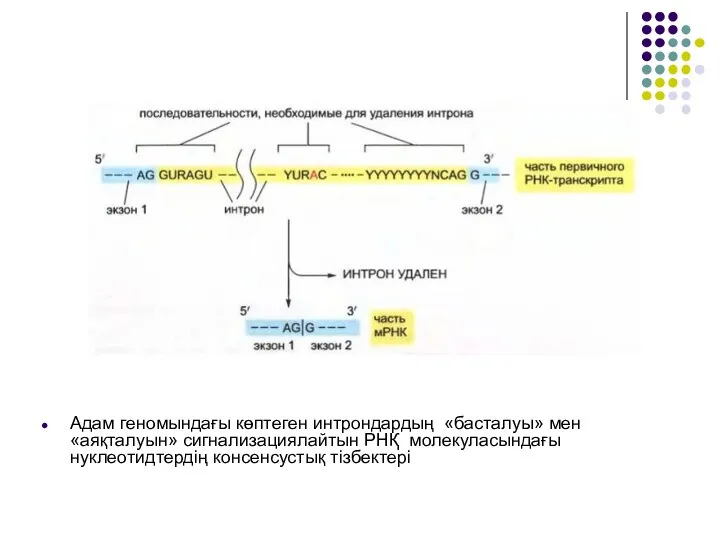
Слайд 15Адам геномындағы көптеген интрондардың «басталуы» мен «аяқталуын» сигнализациялайтын РНҚ молекуласындағы нуклеотидтердің консенсустық
Адам геномындағы көптеген интрондардың «басталуы» мен «аяқталуын» сигнализациялайтын РНҚ молекуласындағы нуклеотидтердің консенсустық
Слайд 16РНҚ сплайсингінің негізгі сатыларын белоктар емес РНҚ молекулалары атқарады. Маманданған РНҚ молекулалары
РНҚ сплайсингінің негізгі сатыларын белоктар емес РНҚ молекулалары атқарады. Маманданған РНҚ молекулалары

Слайд 17Пре-мРНҚ сплайсингінің механизмі
Пре-мРНҚ сплайсингінің механизмі
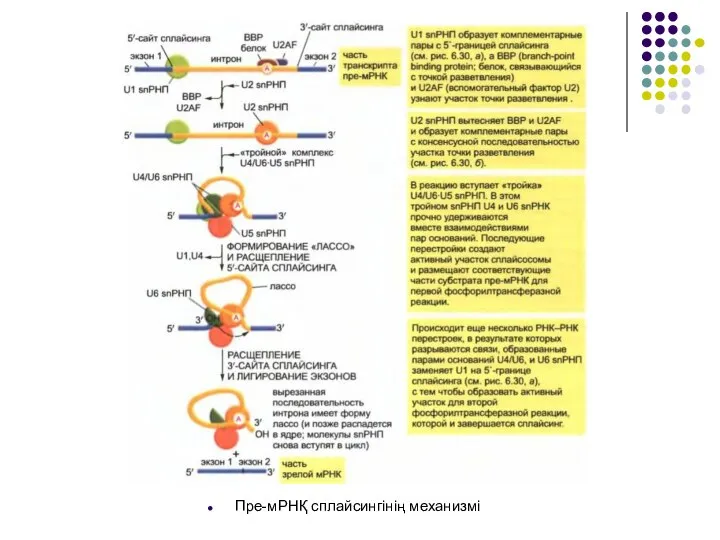
Слайд 18Сплайсинг реакциясының басталуы кезінде сплайсингтің 5 ‘ –шекарасын тану, тармақталу нүктесінің участогын
Сплайсинг реакциясының басталуы кезінде сплайсингтің 5 ‘ –шекарасын тану, тармақталу нүктесінің участогын

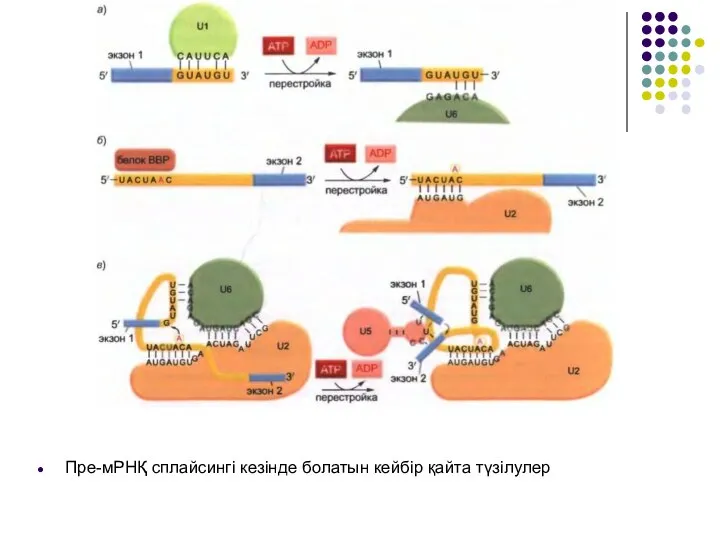
Слайд 19Пре-мРНҚ сплайсингі кезінде болатын кейбір қайта түзілулер
Пре-мРНҚ сплайсингі кезінде болатын кейбір қайта түзілулер
Слайд 20Транскрипция басталарда РНҚ полимеразаның фосфорланған ұшы сплайсоманың бірнеше компоненттерін алып жүреді және
Транскрипция басталарда РНҚ полимеразаның фосфорланған ұшы сплайсоманың бірнеше компоненттерін алып жүреді және

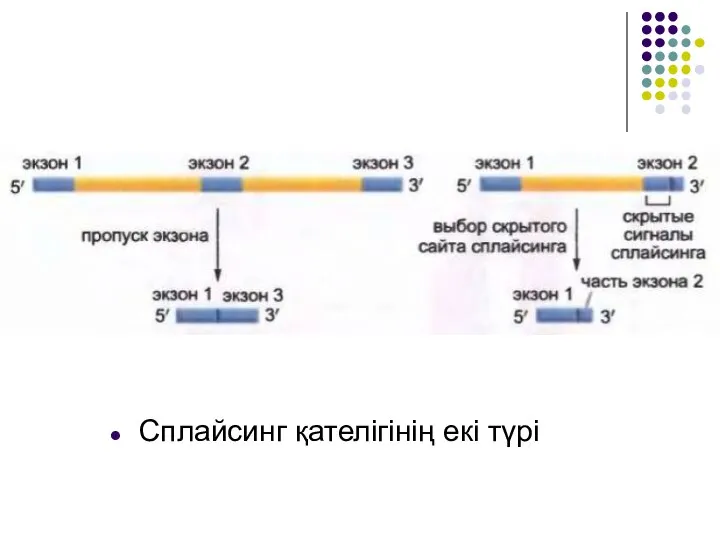
Слайд 21Сплайсинг қателігінің екі түрі
Сплайсинг қателігінің екі түрі
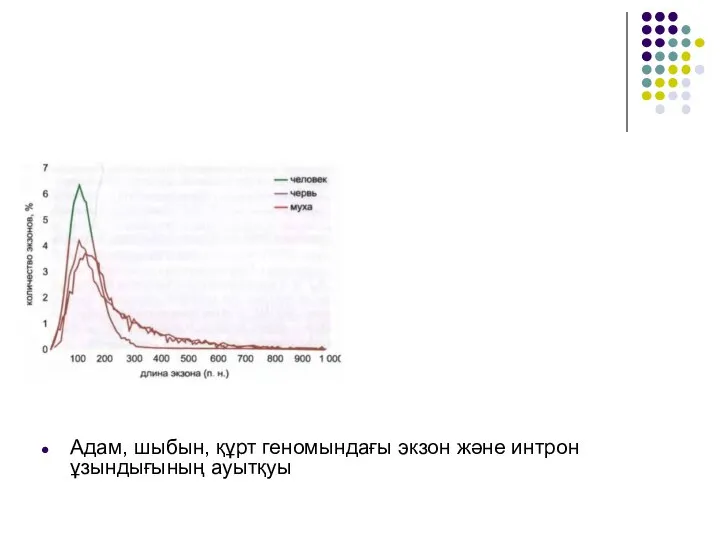
Слайд 22Адам, шыбын, құрт геномындағы экзон және интрон ұзындығының ауытқуы
Адам, шыбын, құрт геномындағы экзон және интрон ұзындығының ауытқуы
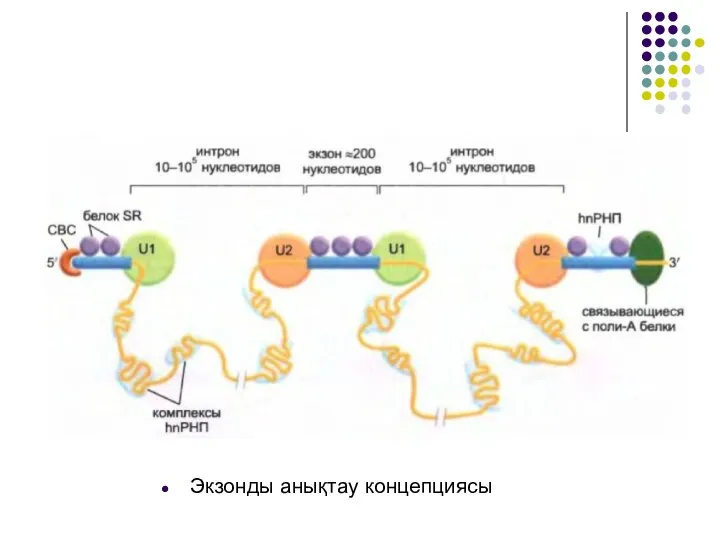
Слайд 23Экзонды анықтау концепциясы
Экзонды анықтау концепциясы
Слайд 24Экзон және интрон шекарасын таңбалау, сонымен қатар сплайсоманы жинақтау РНҚ молекуласында, яғни
Экзон және интрон шекарасын таңбалау, сонымен қатар сплайсоманы жинақтау РНҚ молекуласында, яғни

Слайд 25Ашытқы сияқты қарапайым эукариоттар тек snРНП –дің бір жинағынан тұрады, алайда шыбын,
Ашытқы сияқты қарапайым эукариоттар тек snРНП –дің бір жинағынан тұрады, алайда шыбын,

Слайд 26Трипаносомдарда ( адамдарда африкалық ұйқы ауруын тудыратын қарапайым жануарлар) және нематодтарда сплайсингтің
Трипаносомдарда ( адамдарда африкалық ұйқы ауруын тудыратын қарапайым жануарлар) және нематодтарда сплайсингтің

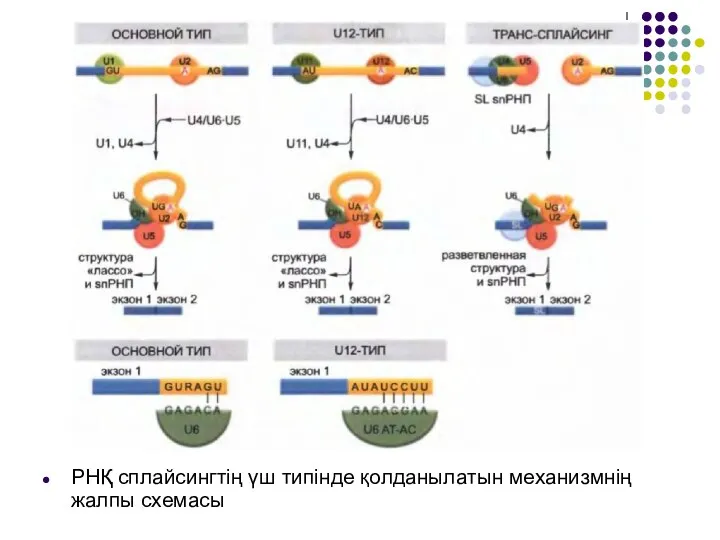
Слайд 27РНҚ сплайсингтің үш типінде қолданылатын механизмнің жалпы схемасы
РНҚ сплайсингтің үш типінде қолданылатын механизмнің жалпы схемасы
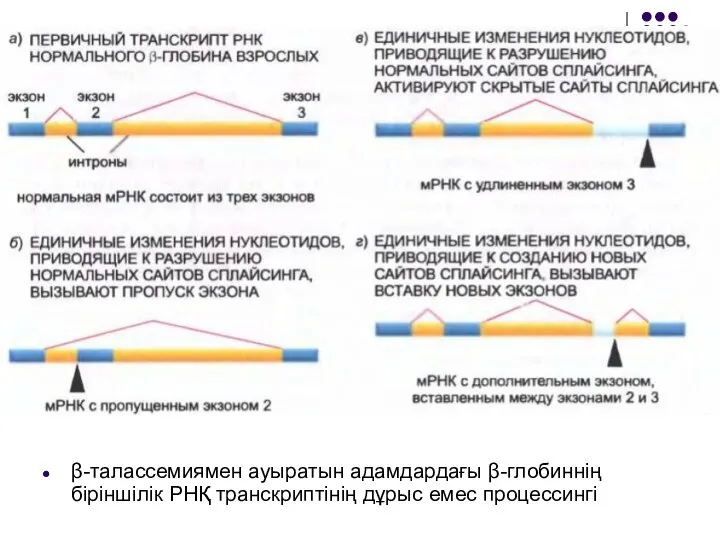
Слайд 28β-талассемиямен ауыратын адамдардағы β-глобиннің біріншілік РНҚ транскриптінің дұрыс емес процессингі
β-талассемиямен ауыратын адамдардағы β-глобиннің біріншілік РНҚ транскриптінің дұрыс емес процессингі
Слайд 29Эволюция кезінде жасушалар басты катализатор ретінде белоктарды емес, РНҚ молекуласын пайдалануы мүмкін
Эволюция кезінде жасушалар басты катализатор ретінде белоктарды емес, РНҚ молекуласын пайдалануы мүмкін

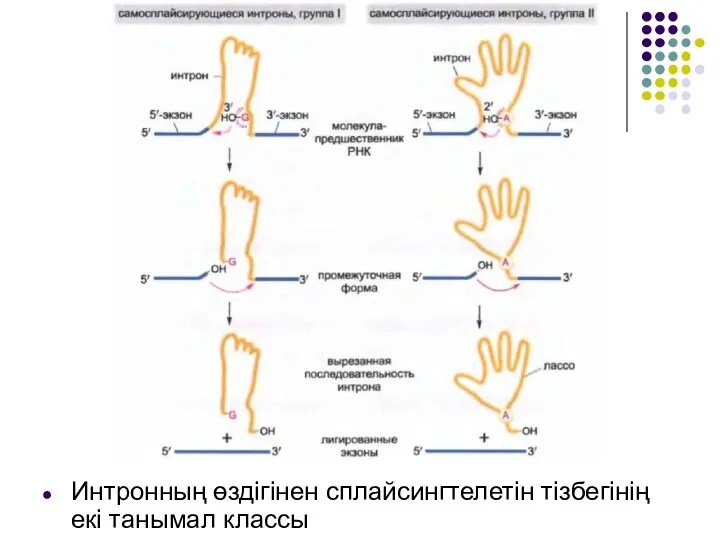
Слайд 30Интронның өздігінен сплайсингтелетін тізбегінің екі танымал классы
Интронның өздігінен сплайсингтелетін тізбегінің екі танымал классы
Слайд 31мРНҚ-ның әр молекуласындағы 3‘-ұшының позициясы геномда кодталған сигнал арқылы анықталады.
Бұндай сигналдар РНҚ-да
мРНҚ-ның әр молекуласындағы 3‘-ұшының позициясы геномда кодталған сигнал арқылы анықталады.
Бұндай сигналдар РНҚ-да

Слайд 32Эукариоттық аРНҚ-ның 3 ‘ –ұшының түзілуі кезіндегі ыдырау мен полиаденилдеуді бағыттайтын консенсустық
Эукариоттық аРНҚ-ның 3 ‘ –ұшының түзілуі кезіндегі ыдырау мен полиаденилдеуді бағыттайтын консенсустық
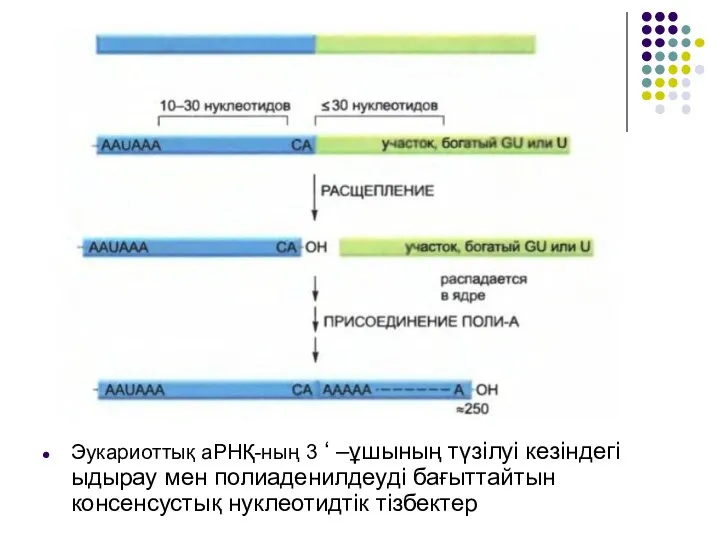
Слайд 33Эукариоттардың мРНҚ 3 ‘ –ұшының түзілуі кезіндегі басты кезеңдерінің кейібіреулері
Эукариоттардың мРНҚ 3 ‘ –ұшының түзілуі кезіндегі басты кезеңдерінің кейібіреулері
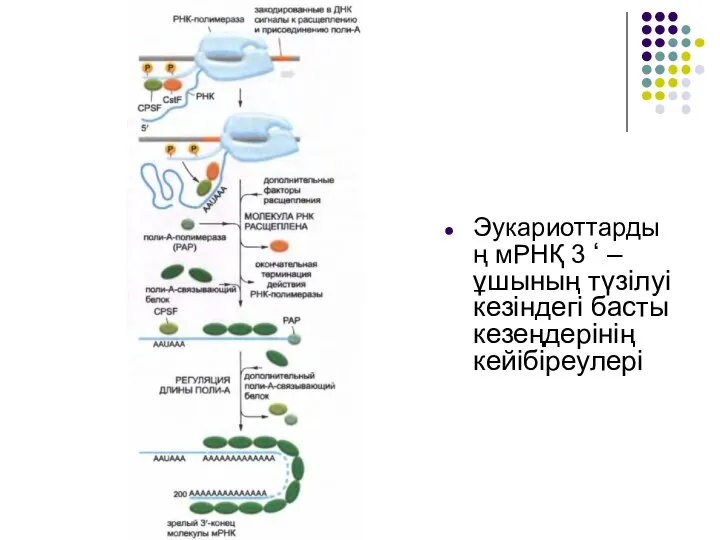
Слайд 34Эукариоттың пре-мРНҚ-сының 3 ‘ –ұшы ыдырағаннан кейін РНҚ полимеразаII транскрипцияны бастайды, бірақ
Эукариоттың пре-мРНҚ-сының 3 ‘ –ұшы ыдырағаннан кейін РНҚ полимеразаII транскрипцияны бастайды, бірақ

Слайд 35Сәтті өңделген мРНҚ молекуласы нуклеоплазма мен цитозольды байланыстыратын ядролық кеуекті кешен арқылы
Сәтті өңделген мРНҚ молекуласы нуклеоплазма мен цитозольды байланыстыратын ядролық кеуекті кешен арқылы

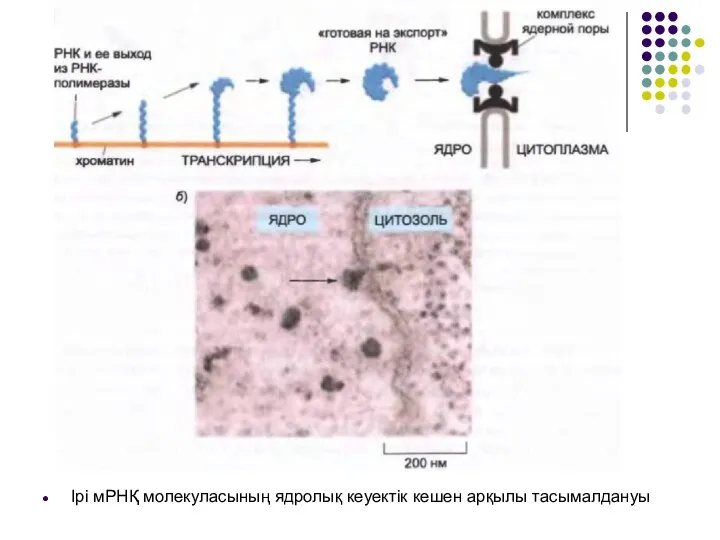
Слайд 36Ірі мРНҚ молекуласының ядролық кеуектік кешен арқылы тасымалдануы
Ірі мРНҚ молекуласының ядролық кеуектік кешен арқылы тасымалдануы
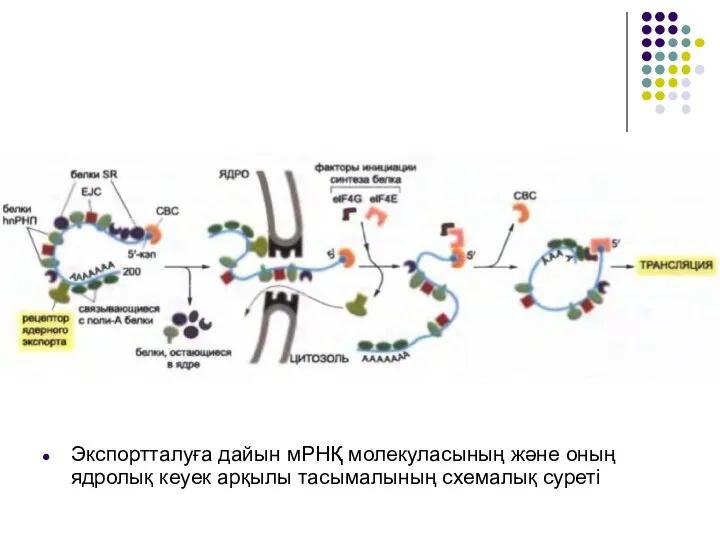
Слайд 37Экспортталуға дайын мРНҚ молекуласының және оның ядролық кеуек арқылы тасымалының схемалық суреті
Экспортталуға дайын мРНҚ молекуласының және оның ядролық кеуек арқылы тасымалының схемалық суреті
Слайд 38Эукариоттарда РРНҚ-ның төрт типі бар, олардың әрқайсысы бір рибосомаға ұқсас көшірме түрінде
Эукариоттарда РРНҚ-ның төрт типі бар, олардың әрқайсысы бір рибосомаға ұқсас көшірме түрінде

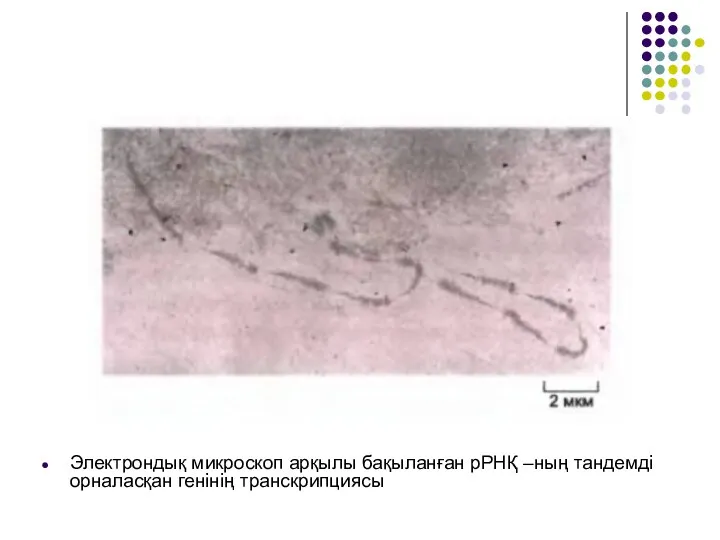
Слайд 39Электрондық микроскоп арқылы бақыланған рРНҚ –ның тандемді орналасқан генінің транскрипциясы
Электрондық микроскоп арқылы бақыланған рРНҚ –ның тандемді орналасқан генінің транскрипциясы
Слайд 40Кең ауқымды химиялық модификациялар 13000-нуклеотидтік рРНҚ ізашарында жүреді,яғни одан рРНҚ молекулалары кесіліп
Кең ауқымды химиялық модификациялар 13000-нуклеотидтік рРНҚ ізашарында жүреді,яғни одан рРНҚ молекулалары кесіліп

Слайд 41Үш рибосомалық РНҚ-ның түзілуі арқылы эукариоттың рРНҚ-сының 45 S ізашары молекуласының нуклеотидтік
Үш рибосомалық РНҚ-ның түзілуі арқылы эукариоттың рРНҚ-сының 45 S ізашары молекуласының нуклеотидтік

Слайд 42Бағыттаушы РНҚ арқылы рРНҚ ізашарының модификациясы
Бағыттаушы РНҚ арқылы рРНҚ ізашарының модификациясы

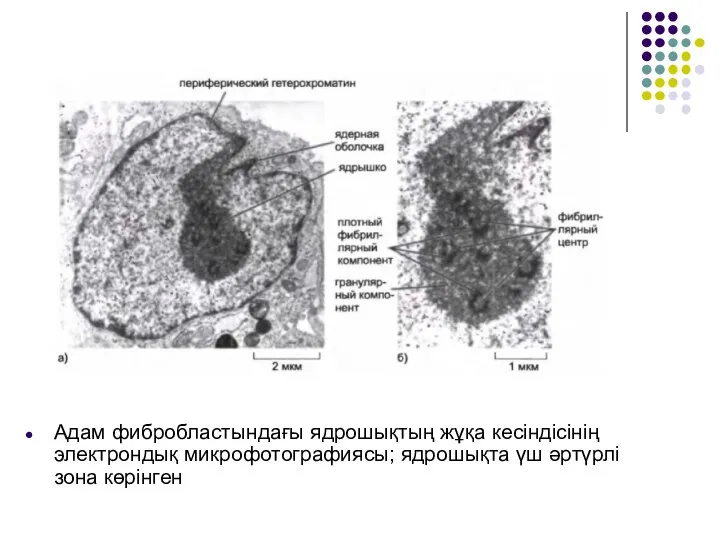
Слайд 43Адам фибробластындағы ядрошықтың жұқа кесіндісінің электрондық микрофотографиясы; ядрошықта үш әртүрлі зона көрінген
Адам фибробластындағы ядрошықтың жұқа кесіндісінің электрондық микрофотографиясы; ядрошықта үш әртүрлі зона көрінген
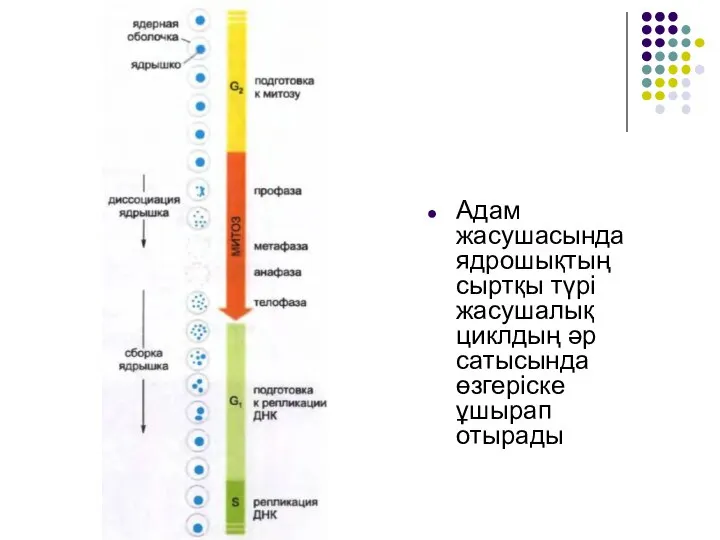
Слайд 44Адам жасушасында ядрошықтың сыртқы түрі жасушалық циклдың әр сатысында өзгеріске ұшырап отырады
Адам жасушасында ядрошықтың сыртқы түрі жасушалық циклдың әр сатысында өзгеріске ұшырап отырады
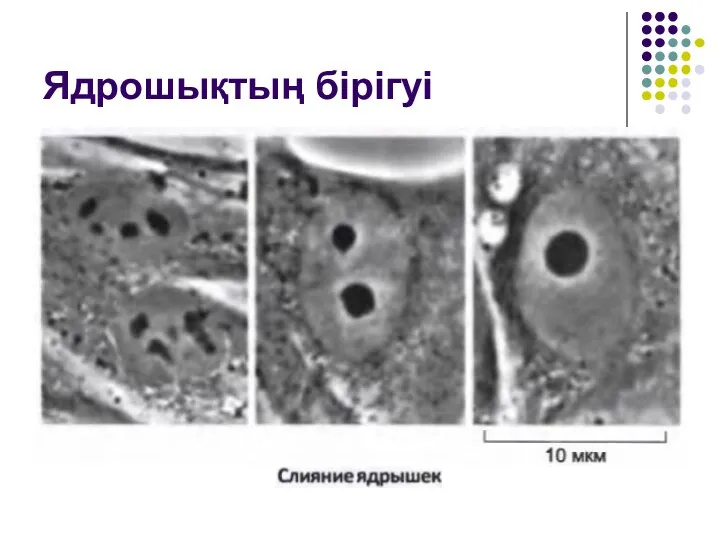
Слайд 45Ядрошықтың бірігуі
Ядрошықтың бірігуі
Слайд 46Ядрошық қызметі, дәлірек айтқанда: рибосома және рибонуклеопротеидтің синтезі
Ядрошық қызметі, дәлірек айтқанда: рибосома және рибонуклеопротеидтің синтезі
 Презентация на тему Класс Земноводные
Презентация на тему Класс Земноводные  Антропогенез. Теория Ч. Дарвина
Антропогенез. Теория Ч. Дарвина Енот - полоскун
Енот - полоскун Презентация на тему ВОЗДУХ И ЕГО ОХРАНА
Презентация на тему ВОЗДУХ И ЕГО ОХРАНА  Фауна открытых биоценозов и агроценозов
Фауна открытых биоценозов и агроценозов Холодний Яр
Холодний Яр Организм как единое целое
Организм как единое целое Перец Скорпион Тринидада
Перец Скорпион Тринидада Птицы Южного Урала
Птицы Южного Урала Центр дрессировки Дог-Профи
Центр дрессировки Дог-Профи Ядро клетки
Ядро клетки Отдел голосеменные
Отдел голосеменные Презентация на тему Доказательства животного происхождения человека
Презентация на тему Доказательства животного происхождения человека  Презентация на тему Железы внутренней секреции
Презентация на тему Железы внутренней секреции  Пока дышу – живу. Модедь Дондерса
Пока дышу – живу. Модедь Дондерса Характеристика и многообразие рыб
Характеристика и многообразие рыб Биосфера без вирусов
Биосфера без вирусов Презентация на тему История собаки динго
Презентация на тему История собаки динго  Животные весной
Животные весной Форма і пластика тварин
Форма і пластика тварин Коровы
Коровы Вирусы – внутриклеточные паразиты
Вирусы – внутриклеточные паразиты Из опыта ландшафтного волонтерства
Из опыта ландшафтного волонтерства obmen_veschestv
obmen_veschestv Nuklein kislatasy
Nuklein kislatasy Взаимосвязь между структурой, свойствами и функциями клеточной мембраны
Взаимосвязь между структурой, свойствами и функциями клеточной мембраны Анализаторы
Анализаторы Происхождение человека
Происхождение человека