- Плазмиды. Структура кольцевых плазмид

Содержание
- 2. Плазмиды С узким спектром хозяев Могут существовать только в очень ограниченном числе видов бактерий. Чаще всего
- 3. Структура кольцевых плазмид Типичная плазмида – это кольцевая двуцепочечная молекула ДНК, каждая цепочка которой ковалентно замкнута
- 4. Репликация плазмид Может происходить по двум механизмам. Ориджин называется oriV. 1. Тета-репликация (однонаправленная или двунаправленная). Вообще
- 5. Репликация плазмид 2. Репликация по механизму катящегося кольца (Rolling Circle Replication, RC) В плазмидах, реплицирующихся по
- 6. RC-репликация 3’-конец двуцепочечного разрыва – затравка для ДНК-полимеразы III, которая и начинает синтез цепочки ДНК, вытесняя
- 7. RC-репликация Другая цепочка материнской ДНК тем временем реплицируется более или менее стандартным образом. Механизм терминации такой
- 8. RC-репликация PolIII начинает синтез второй цепочки. Репликация терминирует стандартно, в результате чего образуется вторая копия ссс
- 9. Интересные подробности о репликации плазмид Белок Rep после своего участия в репликации «приходит в негодность» (это
- 10. Репликация линейных плазмид В линейных плазмидах почти всегда на концах имеются шпилечные структуры. Репликация начинается в
- 11. Репликация линейных плазмид В результате получается «двойная» молекула, в которой концевые шпильки ковалентно сцеплены друг с
- 12. Несовместимость плазмид Несовместимость (inc, incompatibility) двух плазмид – их неспособность существовать в одной бактериальной клетке. Иногда
- 13. Регуляция репликации плазмид и числа их копий Способов такой регуляции очень много, остановимся на основных и
- 14. Регуляция репликации плазмиды ColEI РНК I – короткая РНК, комплементарная РНК II, которая является затравкой для
- 15. Регуляция репликации плазмиды ColEI Если РНК I не синтезируется, то начинается нормальный процесс репликации плазмидной ДНК.
- 16. Регуляция репликации плазмиды ColEI Если РНК I синтезируется, то она немедленно образует прочный комплекс с РНК
- 17. Регуляция репликации плазмиды ColEI Чем больше в клетке копий РНК I, тем слабее репликация, а значит,
- 18. Регуляция репликации плазмиды R1 Регуляция также основана на действии короткой РНК, только она образует комплекс не
- 19. Регуляция репликации плазмиды R1 Но тут вступает в дело РНК СорА, синтезируемая с соответствующего промотора. Она
- 20. Регуляция репликации плазмиды R1 Это очень не по нраву клеточной РНКазе III, которая разрезает этот дуплекс.
- 21. Регуляция репликации плазмиды ColIb-P9 мРНК, кодирующая белок Rep (в данном случае RepZ), кодирует также и лидерный
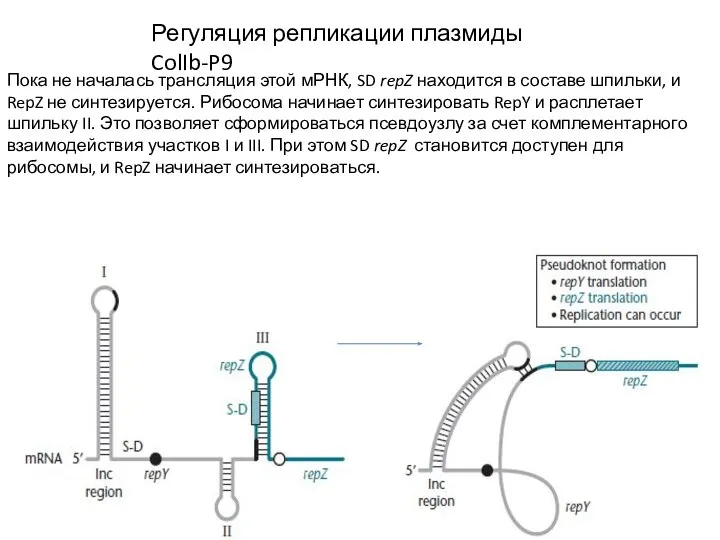
- 22. Регуляция репликации плазмиды ColIb-P9 Пока не началась трансляция этой мРНК, SD repZ находится в составе шпильки,
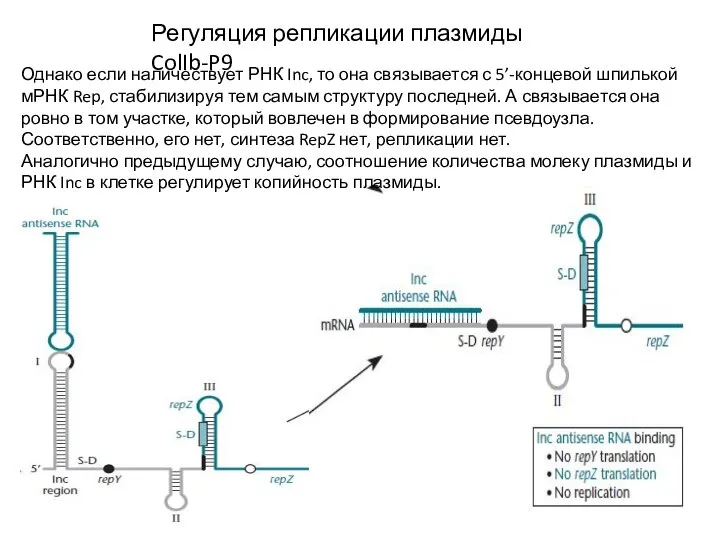
- 23. Регуляция репликации плазмиды ColIb-P9 Однако если наличествует РНК Inc, то она связывается с 5’-концевой шпилькой мРНК
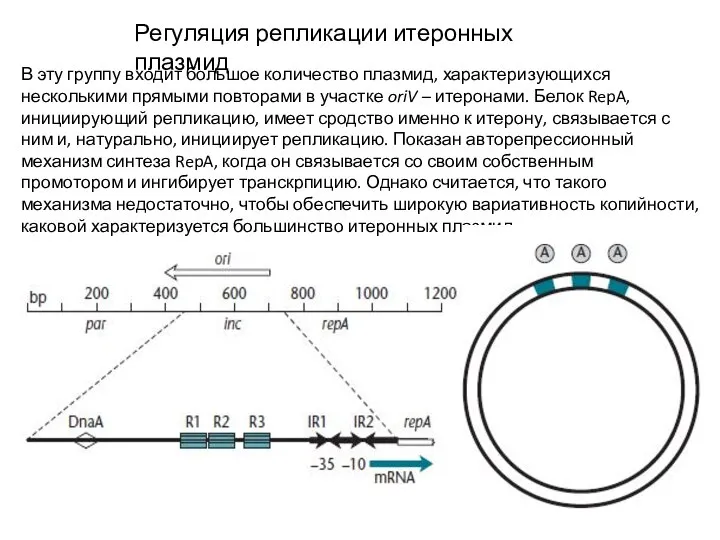
- 24. Регуляция репликации итеронных плазмид В эту группу входит большое количество плазмид, характеризующихся несколькими прямыми повторами в
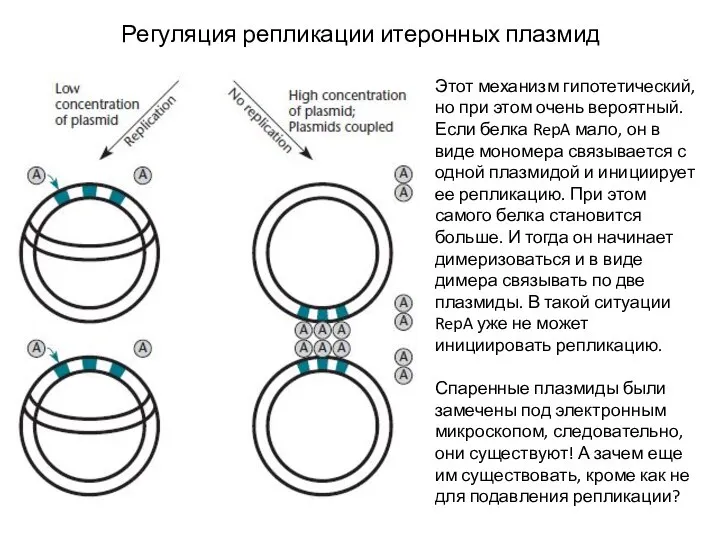
- 25. Регуляция репликации итеронных плазмид Этот механизм гипотетический, но при этом очень вероятный. Если белка RepA мало,
- 26. Потеря плазмиды Потеря плазмиды есть ситуация, в которой после клеточного деления в одну из дочерних клеток
- 27. Распределение плазмиды Разделение плазмиды (partitioning) - совокупность механизмов, по которым увеличивается вероятность попадания данной плазмиды в
- 28. Распределение плазмиды R1 После репликации две плазмиды часто находятся в непосредственной близости друг от друга. Белок
- 29. Распределение плазмиды R1 Когда удлинение филаментов ParM заканчивается (то есть когда плазмиды достигли полюсов и удлиняться
- 30. Распределение плазмид Р1 и F Par-система этих плазмид состоит из генов parA и parB, а также
- 32. Скачать презентацию
Слайд 2Плазмиды
С узким спектром хозяев
Могут существовать только в очень ограниченном числе видов бактерий.
Чаще
Плазмиды
С узким спектром хозяев
Могут существовать только в очень ограниченном числе видов бактерий.
Чаще
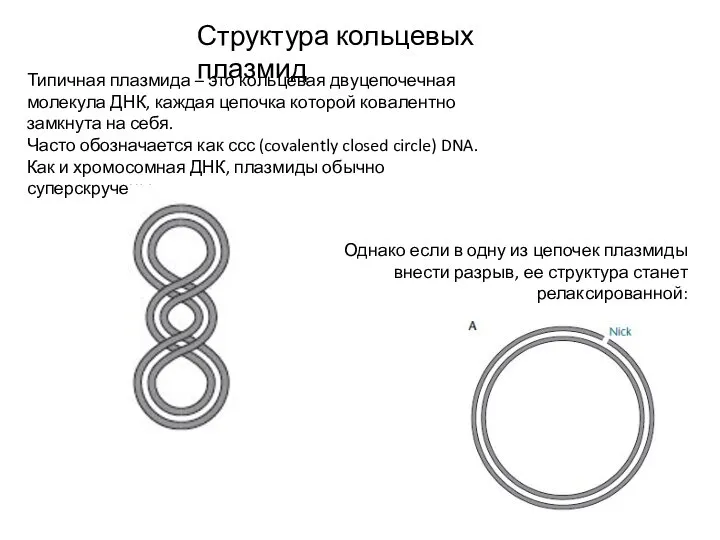
Слайд 3Структура кольцевых плазмид
Типичная плазмида – это кольцевая двуцепочечная молекула ДНК, каждая цепочка
Структура кольцевых плазмид
Типичная плазмида – это кольцевая двуцепочечная молекула ДНК, каждая цепочка
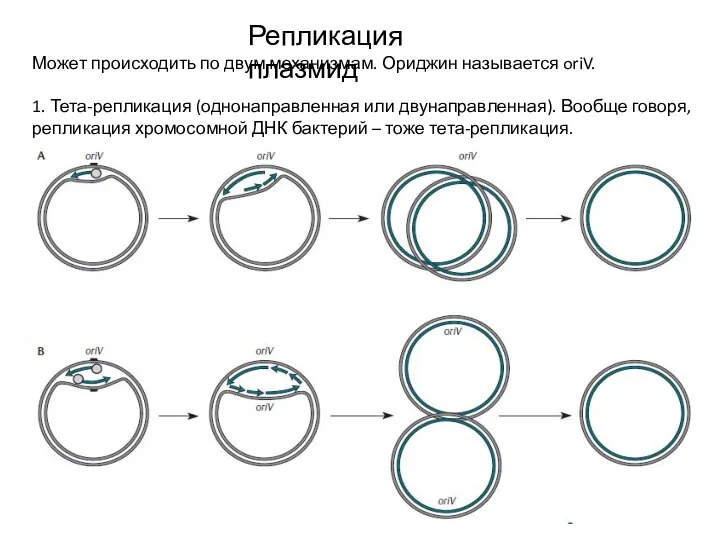
Слайд 4Репликация плазмид
Может происходить по двум механизмам. Ориджин называется oriV.
1. Тета-репликация (однонаправленная или
Репликация плазмид
Может происходить по двум механизмам. Ориджин называется oriV.
1. Тета-репликация (однонаправленная или
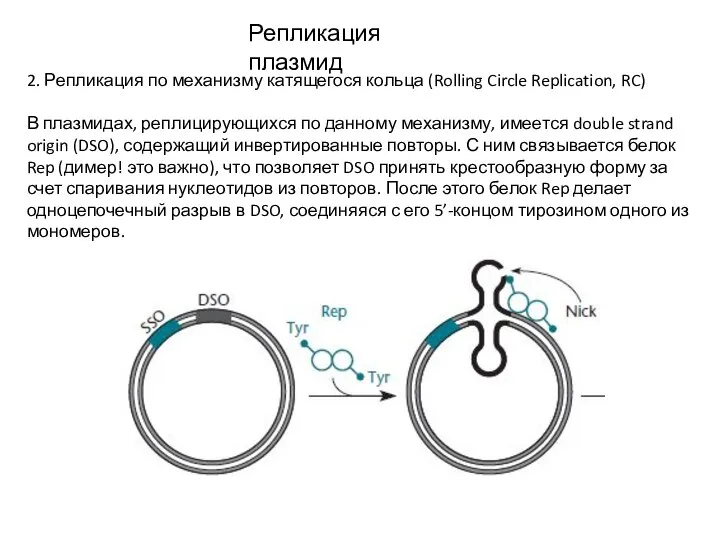
Слайд 5Репликация плазмид
2. Репликация по механизму катящегося кольца (Rolling Circle Replication, RC)
В плазмидах,
Репликация плазмид
2. Репликация по механизму катящегося кольца (Rolling Circle Replication, RC)
В плазмидах,
Слайд 6RC-репликация
3’-конец двуцепочечного разрыва – затравка для ДНК-полимеразы III, которая и начинает синтез
RC-репликация
3’-конец двуцепочечного разрыва – затравка для ДНК-полимеразы III, которая и начинает синтез
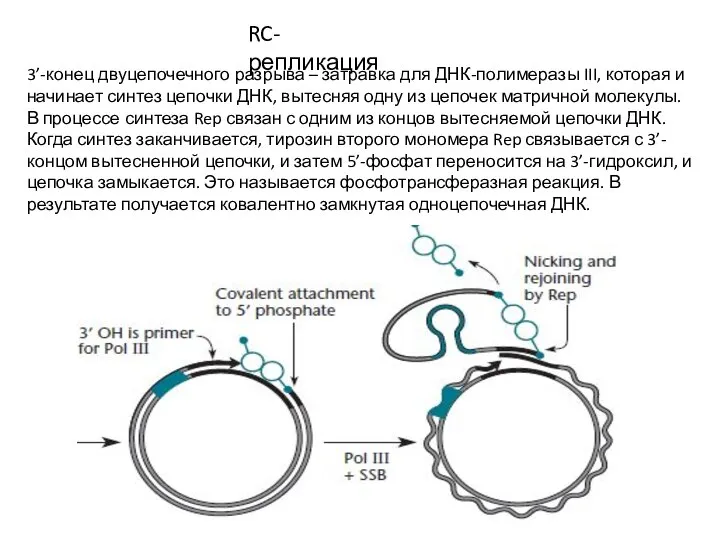
Слайд 7RC-репликация
Другая цепочка материнской ДНК тем временем реплицируется более или менее стандартным образом.
RC-репликация
Другая цепочка материнской ДНК тем временем реплицируется более или менее стандартным образом.
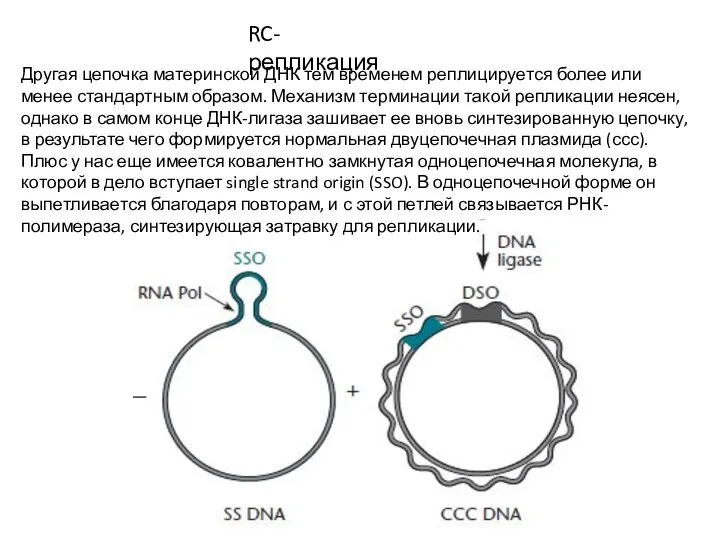
Слайд 8RC-репликация
PolIII начинает синтез второй цепочки.
Репликация терминирует стандартно, в результате чего образуется вторая
RC-репликация
PolIII начинает синтез второй цепочки.
Репликация терминирует стандартно, в результате чего образуется вторая
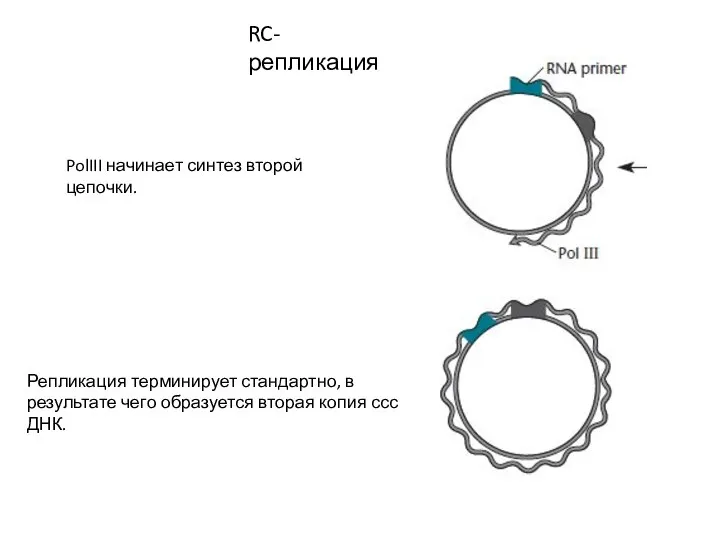
Слайд 9Интересные подробности о репликации плазмид
Белок Rep после своего участия в репликации «приходит
Интересные подробности о репликации плазмид
Белок Rep после своего участия в репликации «приходит

Слайд 10Репликация линейных плазмид
В линейных плазмидах почти всегда на концах имеются шпилечные структуры.
Репликация линейных плазмид
В линейных плазмидах почти всегда на концах имеются шпилечные структуры.
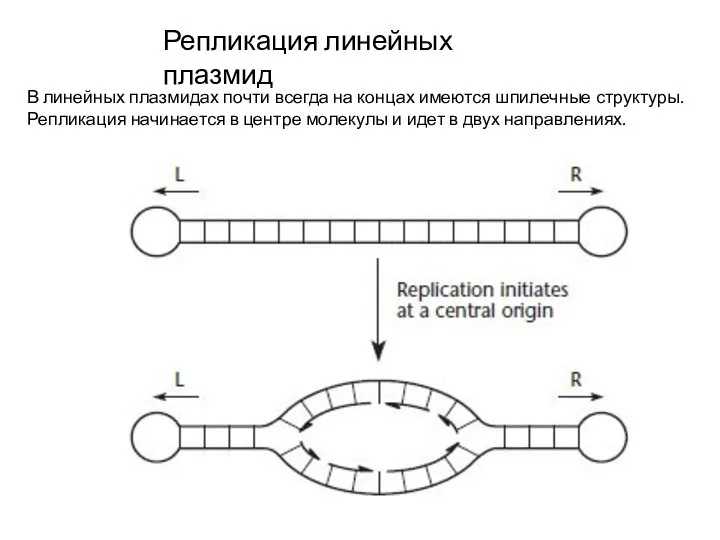
Слайд 11Репликация линейных плазмид
В результате получается «двойная» молекула, в которой концевые шпильки ковалентно
Репликация линейных плазмид
В результате получается «двойная» молекула, в которой концевые шпильки ковалентно
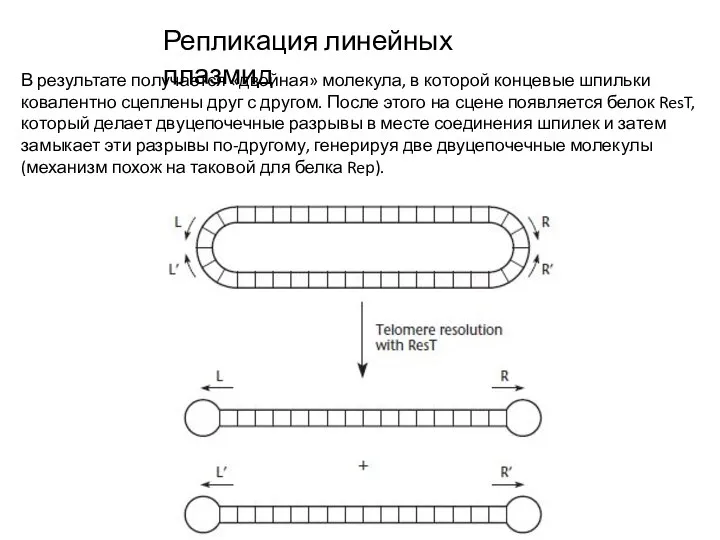
Слайд 12Несовместимость плазмид
Несовместимость (inc, incompatibility) двух плазмид – их неспособность существовать в одной
Несовместимость плазмид
Несовместимость (inc, incompatibility) двух плазмид – их неспособность существовать в одной

Слайд 13Регуляция репликации плазмид и числа их копий
Способов такой регуляции очень много, остановимся
Регуляция репликации плазмид и числа их копий
Способов такой регуляции очень много, остановимся
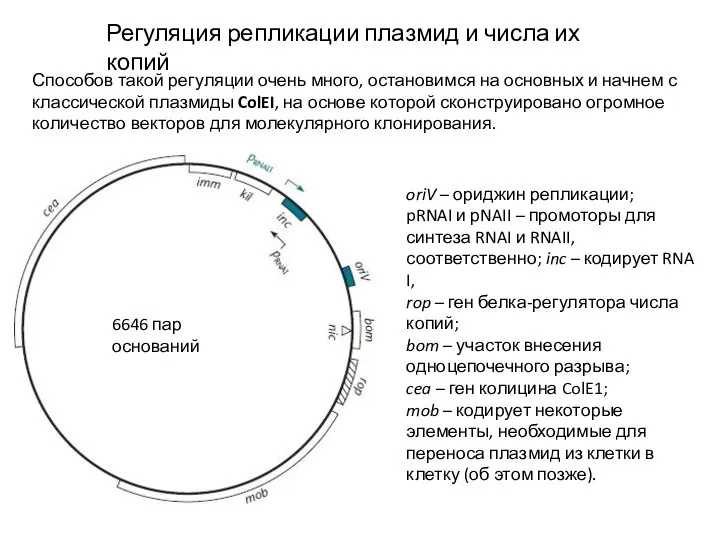
Слайд 14Регуляция репликации плазмиды ColEI
РНК I – короткая РНК, комплементарная РНК II, которая
Регуляция репликации плазмиды ColEI
РНК I – короткая РНК, комплементарная РНК II, которая
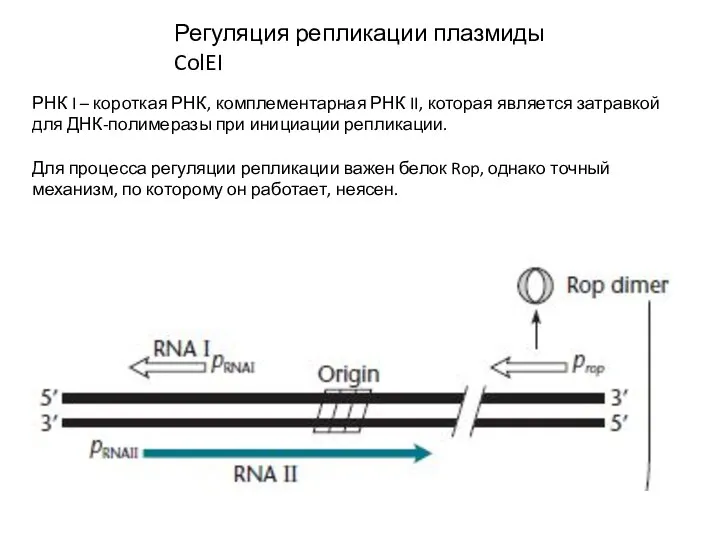
Слайд 15Регуляция репликации плазмиды ColEI
Если РНК I не синтезируется, то начинается нормальный процесс
Регуляция репликации плазмиды ColEI
Если РНК I не синтезируется, то начинается нормальный процесс
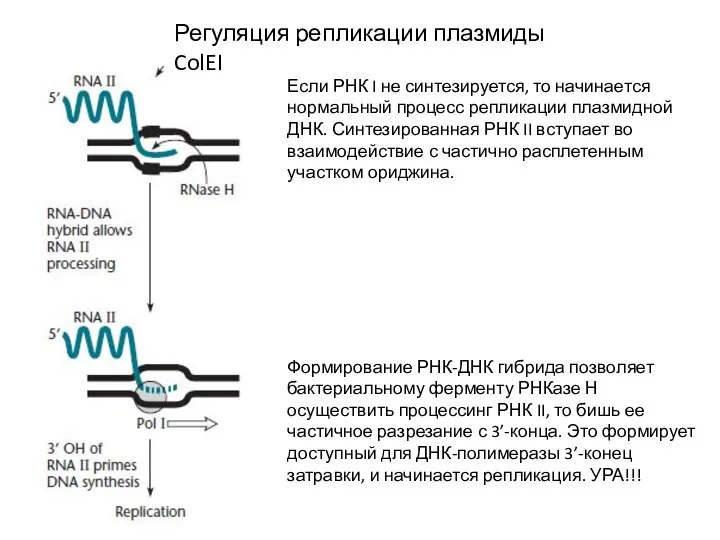
Слайд 16Регуляция репликации плазмиды ColEI
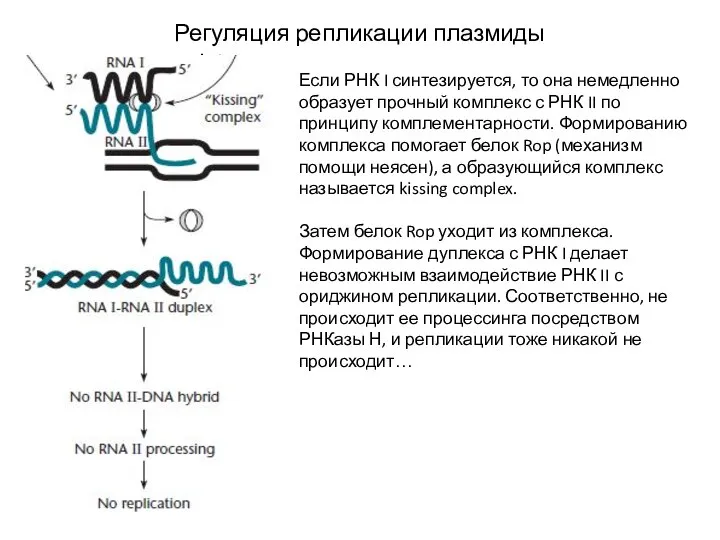
Если РНК I синтезируется, то она немедленно образует прочный
Регуляция репликации плазмиды ColEI
Если РНК I синтезируется, то она немедленно образует прочный
Слайд 17Регуляция репликации плазмиды ColEI
Чем больше в клетке копий РНК I, тем слабее
Регуляция репликации плазмиды ColEI
Чем больше в клетке копий РНК I, тем слабее

Слайд 18Регуляция репликации плазмиды R1
Регуляция также основана на действии короткой РНК, только она
Регуляция репликации плазмиды R1
Регуляция также основана на действии короткой РНК, только она

Слайд 19Регуляция репликации плазмиды R1
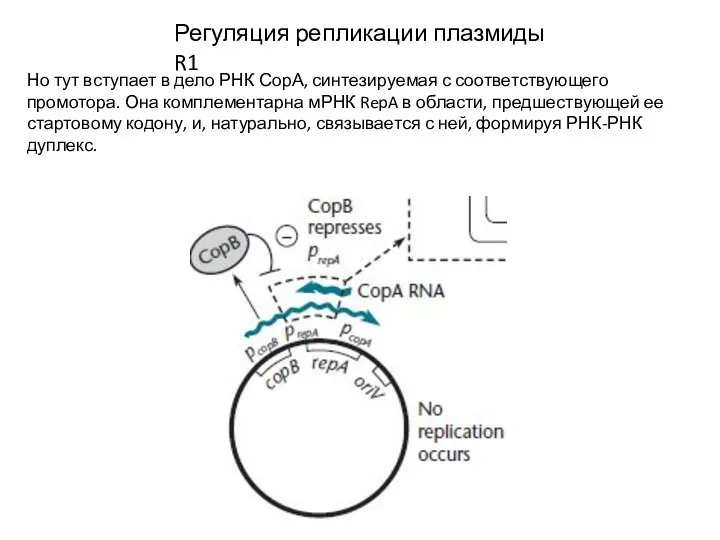
Но тут вступает в дело РНК СорА, синтезируемая с
Регуляция репликации плазмиды R1
Но тут вступает в дело РНК СорА, синтезируемая с
Слайд 20Регуляция репликации плазмиды R1
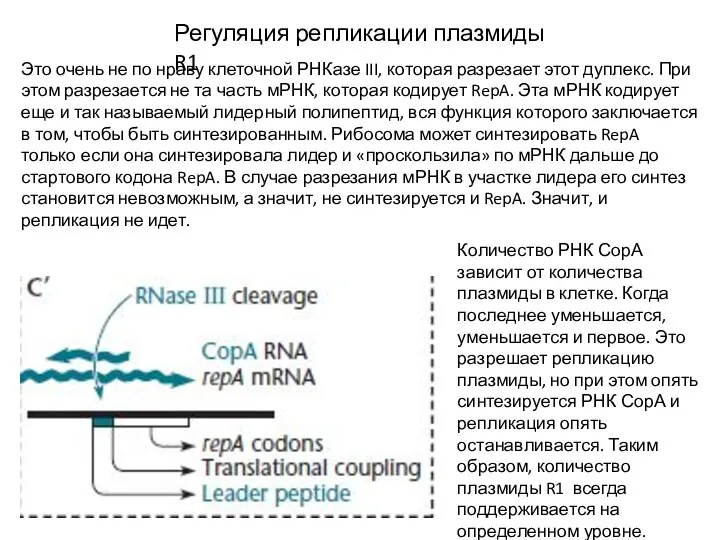
Это очень не по нраву клеточной РНКазе III, которая
Регуляция репликации плазмиды R1
Это очень не по нраву клеточной РНКазе III, которая
Слайд 21Регуляция репликации плазмиды ColIb-P9
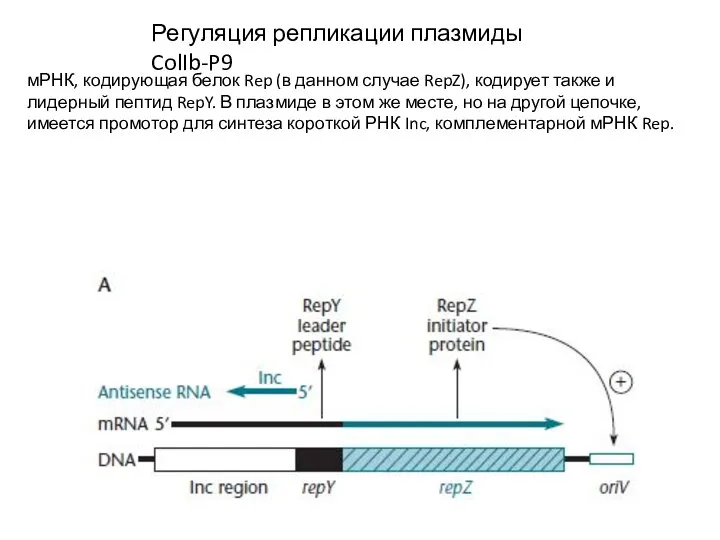
мРНК, кодирующая белок Rep (в данном случае RepZ), кодирует
Регуляция репликации плазмиды ColIb-P9
мРНК, кодирующая белок Rep (в данном случае RepZ), кодирует
Слайд 22Регуляция репликации плазмиды ColIb-P9
Пока не началась трансляция этой мРНК, SD repZ находится
Регуляция репликации плазмиды ColIb-P9
Пока не началась трансляция этой мРНК, SD repZ находится
Слайд 23Регуляция репликации плазмиды ColIb-P9
Однако если наличествует РНК Inc, то она связывается с
Регуляция репликации плазмиды ColIb-P9
Однако если наличествует РНК Inc, то она связывается с
Слайд 24Регуляция репликации итеронных плазмид
В эту группу входит большое количество плазмид, характеризующихся несколькими
Регуляция репликации итеронных плазмид
В эту группу входит большое количество плазмид, характеризующихся несколькими
Слайд 25Регуляция репликации итеронных плазмид
Этот механизм гипотетический, но при этом очень вероятный. Если
Регуляция репликации итеронных плазмид
Этот механизм гипотетический, но при этом очень вероятный. Если
Слайд 26Потеря плазмиды
Потеря плазмиды есть ситуация, в которой после клеточного деления в одну
Потеря плазмиды
Потеря плазмиды есть ситуация, в которой после клеточного деления в одну

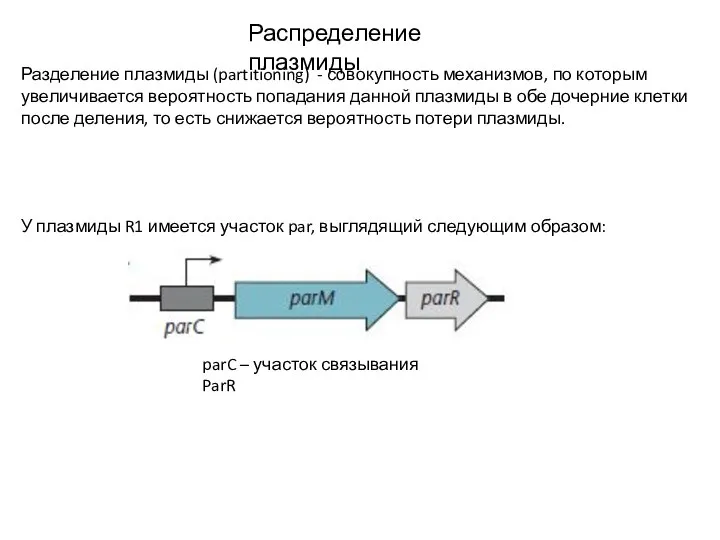
Слайд 27Распределение плазмиды
Разделение плазмиды (partitioning) - совокупность механизмов, по которым увеличивается вероятность попадания
Распределение плазмиды
Разделение плазмиды (partitioning) - совокупность механизмов, по которым увеличивается вероятность попадания
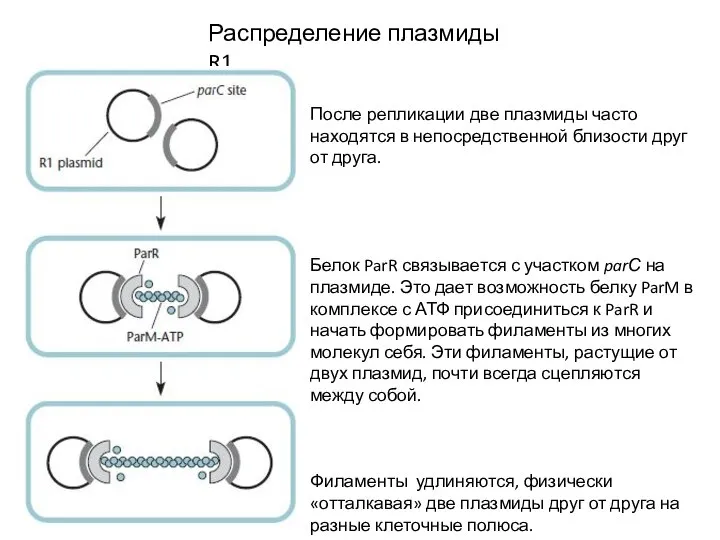
Слайд 28Распределение плазмиды R1
После репликации две плазмиды часто находятся в непосредственной близости друг
Распределение плазмиды R1
После репликации две плазмиды часто находятся в непосредственной близости друг
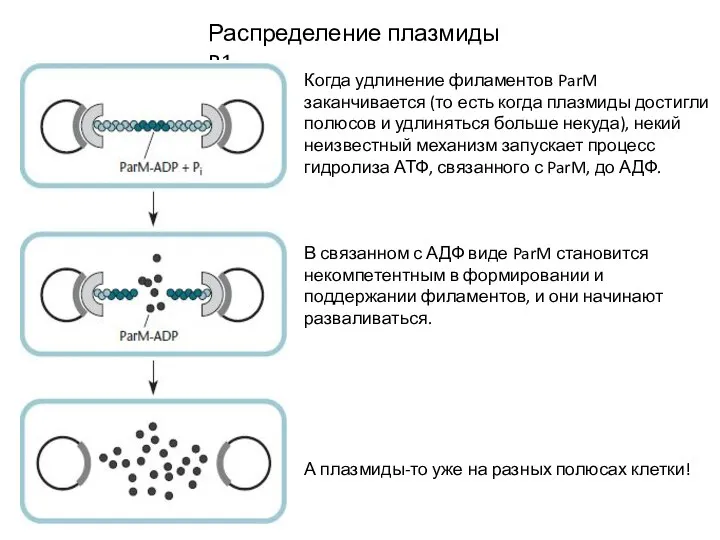
Слайд 29Распределение плазмиды R1
Когда удлинение филаментов ParM заканчивается (то есть когда плазмиды достигли
Распределение плазмиды R1
Когда удлинение филаментов ParM заканчивается (то есть когда плазмиды достигли
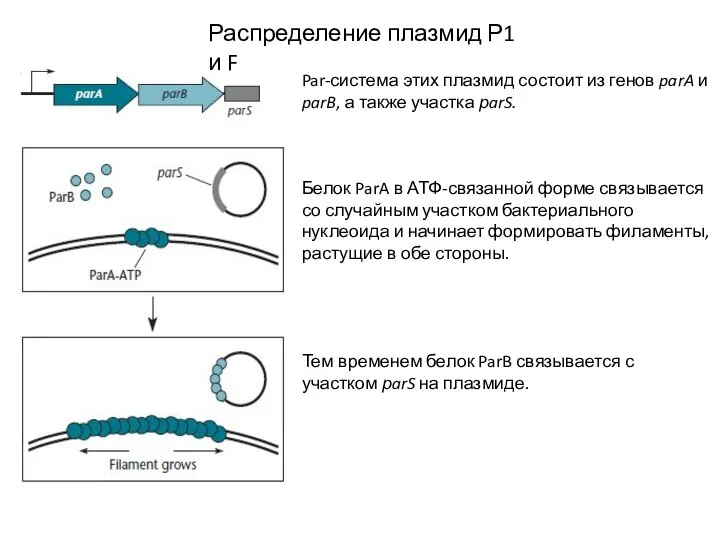
Слайд 30Распределение плазмид Р1 и F
Par-система этих плазмид состоит из генов parA и
Распределение плазмид Р1 и F
Par-система этих плазмид состоит из генов parA и
 Популяция – как основная единица биомониторинга
Популяция – как основная единица биомониторинга Листерии. Морфология
Листерии. Морфология Ткани
Ткани Класс : Моллюски. Тип: Головоногие
Класс : Моллюски. Тип: Головоногие Индивидуальный проект: Путешествие с молекулой кислорода по организму. 10 класс
Индивидуальный проект: Путешествие с молекулой кислорода по организму. 10 класс Презентация на тему ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Презентация на тему ПРОИСХОЖДЕНИЕ И ЭВОЛЮЦИЯ ЧЕЛОВЕКА  Автономная (вегетативная) нервная система. Строение и функции. Иерархия нервных связей внутренностей. Парасимпатическая часть
Автономная (вегетативная) нервная система. Строение и функции. Иерархия нервных связей внутренностей. Парасимпатическая часть Редкие птицы
Редкие птицы Не губите первоцветы, весны первые приметы
Не губите первоцветы, весны первые приметы Здоровье волос питание, нутрицевтическая поддержка
Здоровье волос питание, нутрицевтическая поддержка Обыкновенный скворец
Обыкновенный скворец Строение зуба
Строение зуба Развитие жизни в криптозое
Развитие жизни в криптозое Mother s Love
Mother s Love Химические реакции в нашем организме
Химические реакции в нашем организме Химический состав клетки
Химический состав клетки Домики, которые построила природа
Домики, которые построила природа Презентация на тему Зона степей (4 класс)
Презентация на тему Зона степей (4 класс)  Большой Арктический государственній природный заповедник
Большой Арктический государственній природный заповедник Круги кровообращения
Круги кровообращения a3793c89_lek.16-17-serdechno-sosudistaya_sistema
a3793c89_lek.16-17-serdechno-sosudistaya_sistema Клетки иммунной системы
Клетки иммунной системы Закономерности передачи наследственной информации. Генетическая организация хромосом
Закономерности передачи наследственной информации. Генетическая организация хромосом Черепные нервы
Черепные нервы Презентация на тему Лес – природное сообщество
Презентация на тему Лес – природное сообщество  Взаимоотношения между организмами
Взаимоотношения между организмами Отдел Покрытосеменные (цветковые) растения
Отдел Покрытосеменные (цветковые) растения Определи класс членистоногих
Определи класс членистоногих