- Ферменты

Содержание
- 2. ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ФЕРМЕНТАХ Ферменты, или энзимы, – это биологические катализаторы, ускоряющие химические реакции. Общее число
- 3. Ферменты ускоряют химические реакции в 108 – 1020 раз. Слово фермент произошло от латинского fermentum –
- 4. Большинство ферментов по своей природе являются белками. У некоторых РНК обнаружена способность осуществлять катализ; такие РНК
- 5. Ферменты имеют ряд общих свойств с химическими небелковыми катализаторами: не расходуются в процессе катализа и не
- 7. Ферменты от химических катализаторов отличаются по ряду параметров: ферменты обладают более высокой эффективностью действия; ферменты обладают
- 8. ферменты эффективно действуют в мягких условиях: при температуре 0 – 40 оС, при атмосферном давлении, при
- 9. Структура ферментов Относительная молекулярная масса ферментов может колебаться от 104 до 106 и более. Ферменты –
- 10. В качестве кофакторов могут выступать ионы металлов Fe2+, Mn2+, Mg2+, Zn2+ или сложные органические вещества, которые
- 11. Если кофермент прочно связанный с ферментом называется простетической группой сложного белка. Кофактор легко диссоциируюет из комплекса
- 12. Каталитически активный комплекс фермент – кофактор называют холоферментом. Отделение кофактора от холофермента приводит к образованию неактивного
- 13. В молекуле фермента присутствует активный центр. Активный центр – это область молекулы фермента, в которой происходит
- 14. Фермент может иметь аллостерический центр, служащий для контакта с регуляторной молекулой.
- 15. Активный центр образуют аминокислотные остатки полипептидной цепи. В состав активного центра может входить и небелковый компонент.
- 16. Аминокислотные остатки, образующие активный центр, в полипептидной цепи находятся на значительном расстоянии и оказываются сближенными при
- 17. Некоторые ферменты синтезируются в неактивной форме в виде проферментов, которые затем под действием определенных факторов активируются.
- 18. Изоферменты – это группа ферментов, выполняющих идентичную каталитическую функцию у представителей одного биологического вида, но отличающихся
- 19. Номенклатура и классификация ферментов Тривиальная номенклатура. Например, пепсин (от греч. “пепсис” – “пищеварение”), трипсин (от греч.
- 20. Рациональная номенклатура. Название фермента составляется из названия субстрата и характерного окончания “-аза”. Например, амилаза - катализирует
- 21. Существуют и систематические названия ферментов, включающие названия субстратов и отражающие характер катализируемой реакции: АТФ + D-глюкоза
- 22. В соответствии с катализируемой реакцией все ферменты делятся на 6 классов. Оксидоредуктазы. Катализируют окислительно-восстановительные реакции Трансферазы.
- 23. Лиазы. Катализируют реакции присоединения групп по двойным связям и обратные реакции. Изомеразы. Катализируют реакции изомеризации (внутримолекулярный
- 24. В свою очередь каждый класс подразделяют на подклассы, подклассы – на подподклассы. Ферментам, образующим подподклассы, присваивается
- 25. МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ Последовательность стадий катализа: Взаимодействие субстрата с ферментом в активном центре. Химическое превращение субстрата
- 26. Взаимодействие фермента Е с субстратом S приводит к образованию промежуточного фермент-субстратного комплексаES. При взаимодействии фермента с
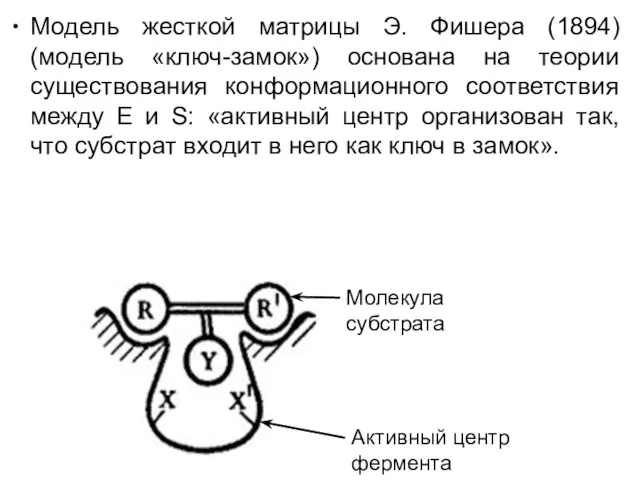
- 27. Модель жесткой матрицы Э. Фишера (1894) (модель «ключ-замок») основана на теории существования конформационного соответствия между Е
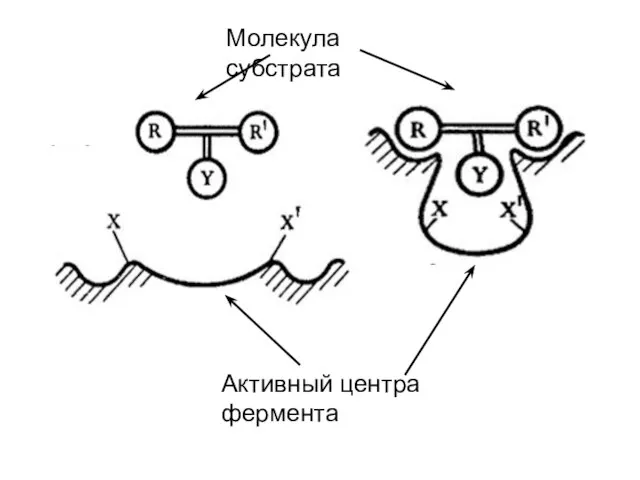
- 28. Модель и теория индуцированного конформационного соответствия между Е и S Д. Кошланда (1958) (модель «рука-перчатка»). Взаимодействие
- 30. В катализе принимают участие: функциональные группы, которые могут быть донорами или акцепторами Н+ (кислотами и основаниями);
- 31. КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их зависимости от различных
- 32. Способы выражения ферментативной активности: Единица активности фермента – количество фермента, которое в стандартных условиях катализирует превращение
- 33. Удельная активность – число единиц ферментативной активности, приходящееся на 1 мг белка (1 мкмоль/мин·мг белка). Активность
- 34. В процессе ферментативной реакции фермент (Е) взаимодействует с субстратом (S) с образованием фермент-субстратный комплекс, который затем
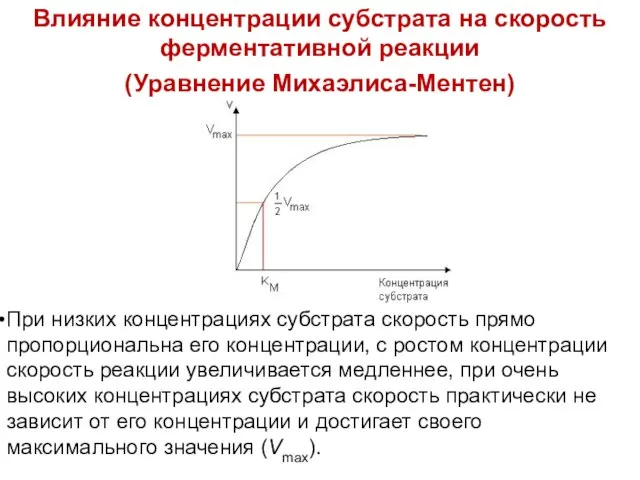
- 35. Влияние концентрации субстрата на скорость ферментативной реакции (Уравнение Михаэлиса-Ментен) При низких концентрациях субстрата скорость прямо пропорциональна
- 36. При высоких концентрациях субстрата все молекулы фермента находятся в составе фермент-субстратного комплекса, и достигается полное насыщение
- 37. График зависимости активности фермента от концентрации субстрата описывается уравнением Михаэлиса – Ментен, которое получило название в
- 38. Физический смысл константы Михаэлиса: При условии, что v = ½ Vmax, получаем KM = [S]. Константа
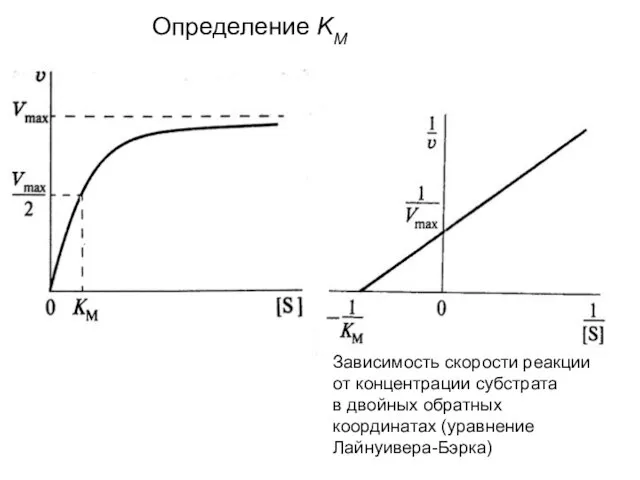
- 39. Зависимость скорости реакции от концентрации субстрата в двойных обратных координатах (уравнение Лайнуивера-Бэрка) Определение KM
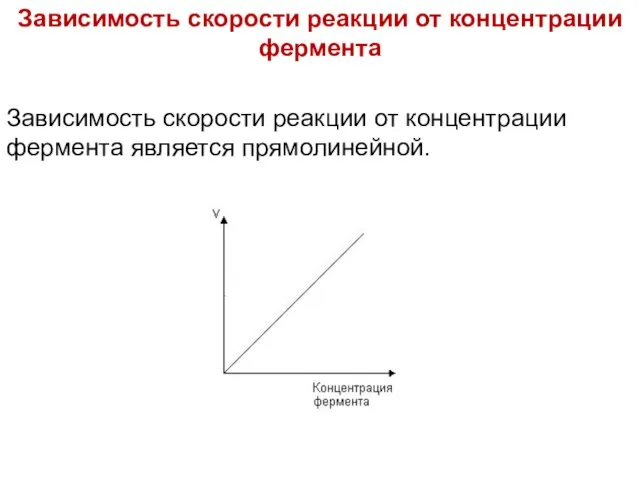
- 40. Зависимость скорости реакции от концентрации фермента Зависимость скорости реакции от концентрации фермента является прямолинейной.
- 41. Не прямолинейная зависимость - при высоких концентрациях фермента наблюдается вследствие нехватки субстрата или агрегации молекул фермента
- 42. Зависимость скорости ферментативной реакции от температуры Зависимость скорости ферментативной реакции от температуры представлена на графике
- 43. При низких температурах (приблизительно до 40 – 50 оС) повышение температуры на каждые 10 оС в
- 44. Температура, при которой активность фермента максимальна, называется температурным оптимумом. Температурный оптимум ферментов термофильных микроорганизмов находится в
- 45. Зависимость скорости ферментативной реакции от рН График зависимости от рН имеет колоколообразную форму Значение рН, при
- 46. Значения рН-оптимума для различных ферментов колеблются в широких пределах.
- 47. Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает влияние на: ионизацию аминокислотных
- 48. Ингибирование ферментов Скорость ферментативной реакции может быть снижена при участиии ингибиторов. Ингибитор – вещество, специфически уменьшающее
- 49. Ингибиторы можно разделить на два основных типа: необратимые и обратимые. Необратимые ингибиторы (I) связываются с ферментом
- 50. Примером необратимого ингибитора является диизопропилфторфосфат (ДФФ). ДФФ ингибирует фермент ацетилхолинэстеразу, играющего важную роль в передаче нервного
- 51. Обратимые ингибиторы при определенных условиях могут быть легко отделены от фермента. Активность последнего при этом восстанавливается:
- 52. Типы ингибирования Конкурентное ингибирование. Конкурентный ингибитор, являясь структурным аналогом субстрата, взаимодействует с активным центром фермента и
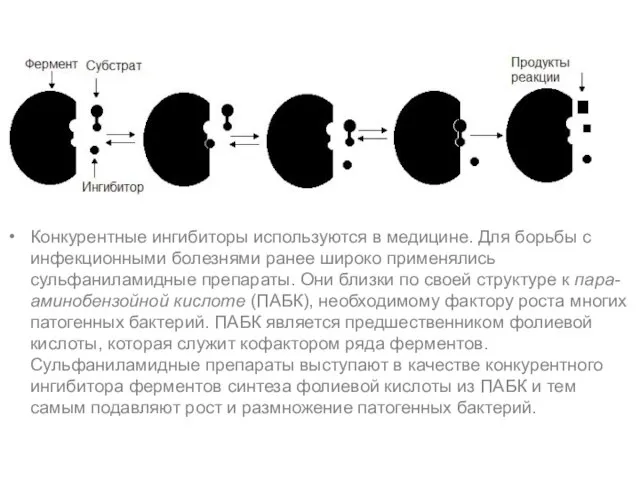
- 53. Конкурентные ингибиторы используются в медицине. Для борьбы с инфекционными болезнями ранее широко применялись сульфаниламидные препараты. Они
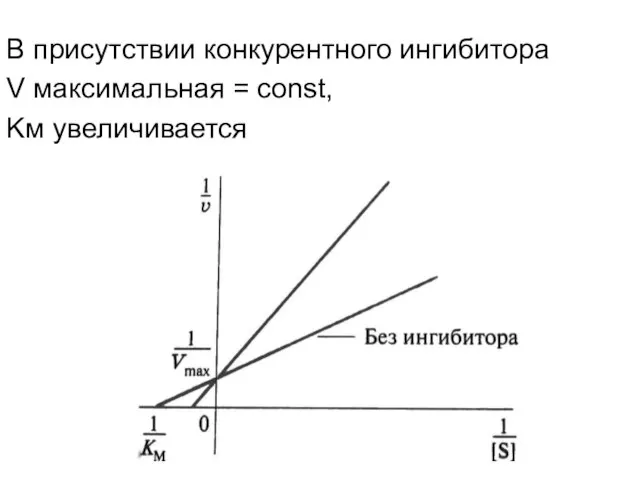
- 54. В присутствии конкурентного ингибитора V максимальная = const, Kм увеличивается
- 55. Неконкурентное ингибирование Неконкурентные ингибиторы по структуре не сходны с субстратом и при образовании EI взаимодействуют не
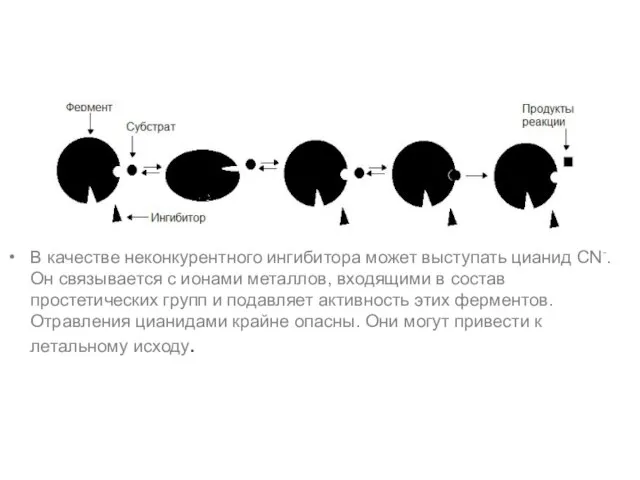
- 56. В качестве неконкурентного ингибитора может выступать цианид CN-. Он связывается с ионами металлов, входящими в состав
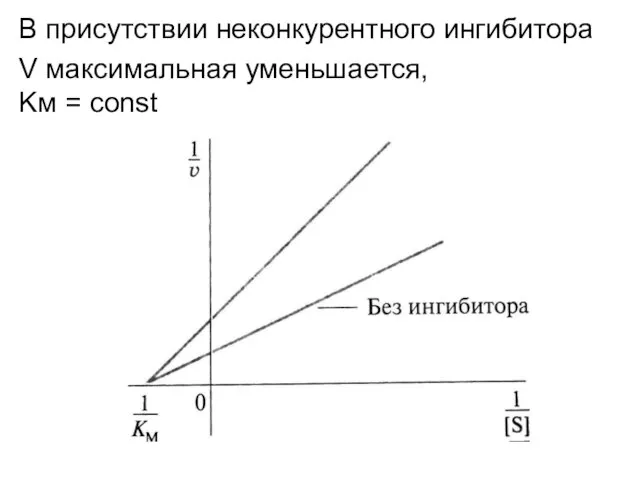
- 57. В присутствии неконкурентного ингибитора V максимальная уменьшается, Kм = const
- 58. Бесконкурентное ингибирование Ингибитор обратимо взаимодействует с ферментом только после образования фермент-субстратного комплекса. Образующийся в этом случае
- 59. Смешанное ингибирование сочетает в себе конкурентное и неконкурентное торможение. Ингибитор, присоединяется в активном центре фермента, изменяет
- 60. Регуляция каталитической активности ферментов Регуляция путём посттрансляционной ковалентной модификации молекулы фермента (ограниченный протеолиз, фосфорилирование, метилирование, гликозилирование
- 61. Регуляция функционирования ферментных систем Ферментные системы обладают способностью поддерживать необходимую скорость суммарного процесса преобразования исходного субстрата
- 62. Регуляция биосинтеза ферментов Индукция фермента (синтез de novo) происходит при повышении концентрации субстрата в клетке. Репрессия
- 63. Аллостерические ферменты Термин «аллостерический» происходит от греческих слов allo – другой, stereo – участок. Аллостерические ферменты
- 64. С аллостерическим центром связываются аллостерические эффекторы - вещества, способные изменять активность ферментов. Эффекторы бывают положительными, активирующими
- 65. Регуляция мультиферментных систем Некоторые ферменты действуют согласованно, объединяясь в мультиферментные системы, в которых каждый фермент катализирует
- 66. В мультиферментной системе есть фермент, который определяет скорость всей последовательности реакций. Этот фермент, как правило, бывает
- 67. Применение ферментов Отрасли промышленности, в которых применяются ферменты: хлебопечение, пивоварение, виноделие, чайное, кожевенное и меховое производства,
- 68. Применение ферментов в медицине Энзимопатология - изучает молекулярные основы развития патологического процесса, исходя из данных о
- 69. Энзимотерапия – использование ферментов, активаторов и ингибиторов в качестве лекарственных средств. Например: протеолитические ферменты: пепсин, трипсин,
- 71. Скачать презентацию
Слайд 2ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ФЕРМЕНТАХ
Ферменты, или энзимы, – это биологические катализаторы, ускоряющие химические
ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ФЕРМЕНТАХ
Ферменты, или энзимы, – это биологические катализаторы, ускоряющие химические

Слайд 3Ферменты ускоряют химические реакции в 108 – 1020 раз.
Слово фермент произошло
Ферменты ускоряют химические реакции в 108 – 1020 раз.
Слово фермент произошло

Слайд 4
Большинство ферментов по своей природе являются белками.
У некоторых РНК обнаружена способность
Большинство ферментов по своей природе являются белками.
У некоторых РНК обнаружена способность

Слайд 5Ферменты имеют ряд общих свойств с химическими небелковыми катализаторами:
не расходуются в
Ферменты имеют ряд общих свойств с химическими небелковыми катализаторами:
не расходуются в

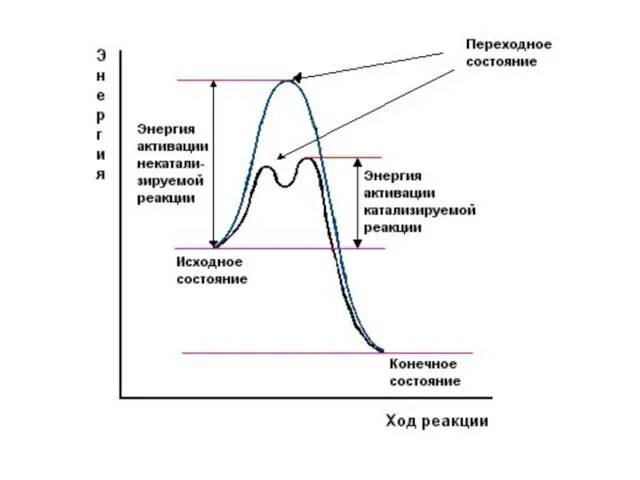
Слайд 7Ферменты от химических катализаторов отличаются по ряду параметров:
ферменты обладают более высокой
ферменты обладают более высокой

Слайд 8ферменты эффективно действуют в мягких условиях: при температуре 0 – 40 оС,

Слайд 9Структура ферментов
Относительная молекулярная масса ферментов может колебаться от 104 до 106 и
Структура ферментов
Относительная молекулярная масса ферментов может колебаться от 104 до 106 и

Слайд 10В качестве кофакторов могут выступать ионы металлов Fe2+, Mn2+, Mg2+, Zn2+
или
В качестве кофакторов могут выступать ионы металлов Fe2+, Mn2+, Mg2+, Zn2+
или
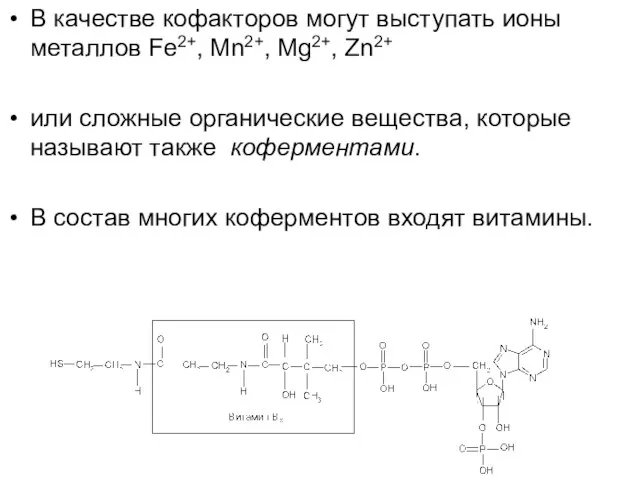
Слайд 11Если кофермент прочно связанный с ферментом называется простетической группой сложного белка.
Кофактор
Если кофермент прочно связанный с ферментом называется простетической группой сложного белка.
Кофактор

Слайд 12Каталитически активный комплекс фермент – кофактор называют холоферментом.
Отделение кофактора от холофермента
Отделение кофактора от холофермента

Слайд 13В молекуле фермента присутствует активный центр.
Активный центр – это область молекулы
В молекуле фермента присутствует активный центр.
Активный центр – это область молекулы

Слайд 14Фермент может иметь аллостерический центр, служащий для контакта с регуляторной молекулой.
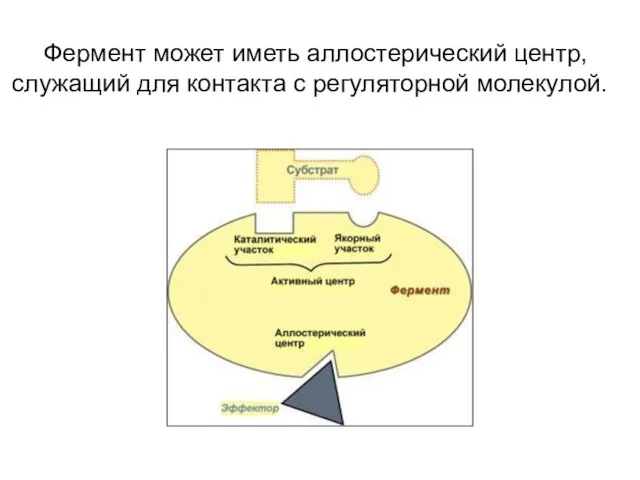
Слайд 15Активный центр образуют аминокислотные остатки полипептидной цепи.
В состав активного центра может
В состав активного центра может

Слайд 16Аминокислотные остатки, образующие активный центр, в полипептидной цепи находятся на значительном расстоянии
Аминокислотные остатки, образующие активный центр, в полипептидной цепи находятся на значительном расстоянии
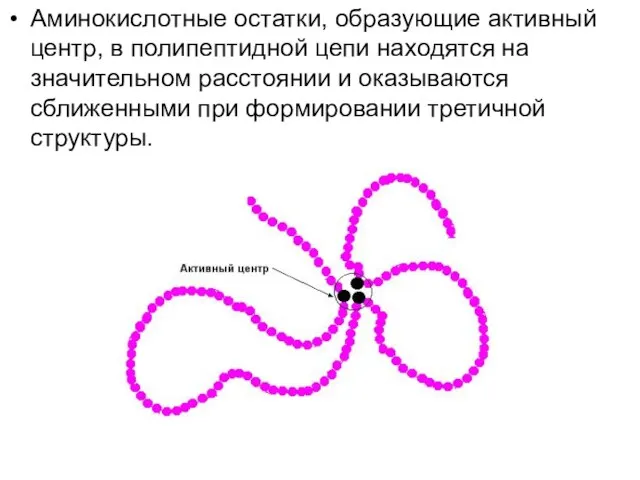
Слайд 17Некоторые ферменты синтезируются в неактивной форме в виде проферментов, которые затем под

Слайд 18Изоферменты – это группа ферментов, выполняющих идентичную каталитическую функцию у представителей одного
Изоферменты – это группа ферментов, выполняющих идентичную каталитическую функцию у представителей одного

Слайд 19Номенклатура и классификация ферментов
Тривиальная номенклатура.
Например,
пепсин (от греч. “пепсис” – “пищеварение”),
Номенклатура и классификация ферментов
Тривиальная номенклатура.
Например,
пепсин (от греч. “пепсис” – “пищеварение”),

Слайд 20Рациональная номенклатура.
Название фермента составляется из названия субстрата и характерного окончания “-аза”.
Рациональная номенклатура.
Название фермента составляется из названия субстрата и характерного окончания “-аза”.

Слайд 21Существуют и систематические названия ферментов, включающие названия субстратов и отражающие характер катализируемой

Слайд 22В соответствии с катализируемой реакцией все ферменты делятся на 6 классов.
Оксидоредуктазы.
Катализируют
В соответствии с катализируемой реакцией все ферменты делятся на 6 классов.
Оксидоредуктазы.
Катализируют

Слайд 23Лиазы.
Катализируют реакции присоединения групп по двойным связям и обратные реакции.
Изомеразы.
Катализируют
Лиазы.
Катализируют реакции присоединения групп по двойным связям и обратные реакции.
Изомеразы.
Катализируют

Слайд 24В свою очередь каждый класс подразделяют
на подклассы,
подклассы – на подподклассы.
В свою очередь каждый класс подразделяют
на подклассы,
подклассы – на подподклассы.

Слайд 25МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ
Последовательность стадий катализа:
Взаимодействие субстрата с ферментом в активном центре.
Химическое превращение
МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ
Последовательность стадий катализа:
Взаимодействие субстрата с ферментом в активном центре.
Химическое превращение

Слайд 26Взаимодействие фермента Е с субстратом S приводит к образованию промежуточного фермент-субстратного комплексаES.
Взаимодействие фермента Е с субстратом S приводит к образованию промежуточного фермент-субстратного комплексаES.

Слайд 27Модель жесткой матрицы Э. Фишера (1894) (модель «ключ-замок») основана на теории существования
Модель жесткой матрицы Э. Фишера (1894) (модель «ключ-замок») основана на теории существования
Слайд 28Модель и теория индуцированного конформационного соответствия
между Е и S Д. Кошланда
Модель и теория индуцированного конформационного соответствия между Е и S Д. Кошланда

Слайд 30В катализе принимают участие:
функциональные группы, которые могут быть донорами или акцепторами Н+
функциональные группы, которые могут быть донорами или акцепторами Н+

Слайд 31КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их
Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их

Слайд 32Способы выражения ферментативной активности:
Единица активности фермента – количество фермента, которое в стандартных
Способы выражения ферментативной активности:
Единица активности фермента – количество фермента, которое в стандартных

Слайд 33Удельная активность – число единиц ферментативной активности, приходящееся на 1 мг белка
Удельная активность – число единиц ферментативной активности, приходящееся на 1 мг белка

Слайд 34В процессе ферментативной реакции фермент (Е) взаимодействует с субстратом (S) с образованием

Слайд 35Влияние концентрации субстрата на скорость ферментативной реакции
(Уравнение Михаэлиса-Ментен)
При низких концентрациях субстрата
Влияние концентрации субстрата на скорость ферментативной реакции
(Уравнение Михаэлиса-Ментен)
При низких концентрациях субстрата
Слайд 36
При высоких концентрациях субстрата все молекулы фермента находятся в составе фермент-субстратного комплекса,
При высоких концентрациях субстрата все молекулы фермента находятся в составе фермент-субстратного комплекса,

Слайд 37График зависимости активности фермента от концентрации субстрата описывается уравнением Михаэлиса – Ментен,
График зависимости активности фермента от концентрации субстрата описывается уравнением Михаэлиса – Ментен,

Слайд 38
Физический смысл константы Михаэлиса:
При условии, что v = ½ Vmax, получаем
Физический смысл константы Михаэлиса:
При условии, что v = ½ Vmax, получаем

Слайд 39Зависимость скорости реакции
от концентрации субстрата
в двойных обратных
координатах (уравнение
Лайнуивера-Бэрка)
Определение
Зависимость скорости реакции
от концентрации субстрата
в двойных обратных
координатах (уравнение
Лайнуивера-Бэрка)
Определение
Слайд 40Зависимость скорости реакции от концентрации фермента
Зависимость скорости реакции от концентрации фермента является
Зависимость скорости реакции от концентрации фермента
Зависимость скорости реакции от концентрации фермента является
Слайд 41Не прямолинейная зависимость
- при высоких концентрациях фермента наблюдается вследствие нехватки субстрата
- при высоких концентрациях фермента наблюдается вследствие нехватки субстрата

Слайд 42Зависимость скорости ферментативной реакции от температуры
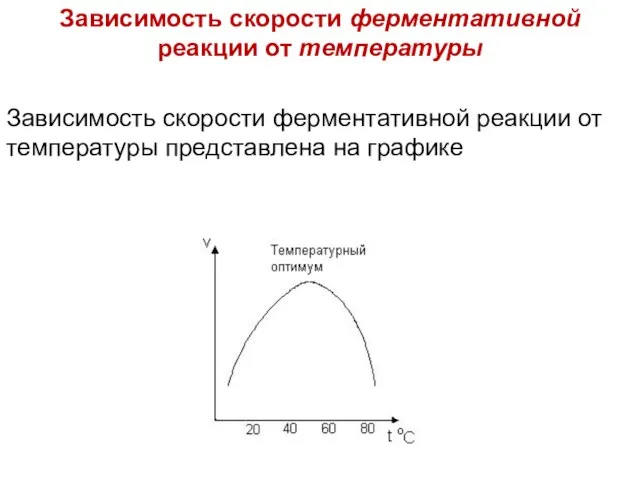
Зависимость скорости ферментативной реакции от температуры представлена
Зависимость скорости ферментативной реакции от температуры
Зависимость скорости ферментативной реакции от температуры представлена
Слайд 43При низких температурах (приблизительно до 40 – 50 оС) повышение температуры на
При низких температурах (приблизительно до 40 – 50 оС) повышение температуры на

Слайд 44Температура, при которой активность фермента максимальна, называется температурным оптимумом.
Температурный оптимум ферментов
Температурный оптимум ферментов

Слайд 45Зависимость скорости ферментативной реакции от рН
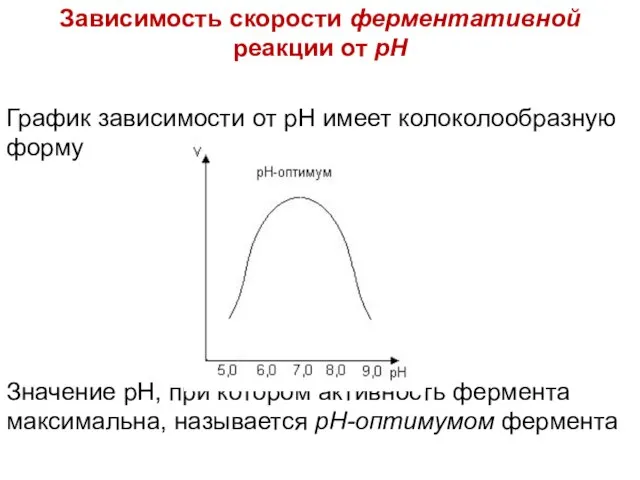
График зависимости от рН имеет колоколообразную форму
Значение
Зависимость скорости ферментативной реакции от рН
График зависимости от рН имеет колоколообразную форму
Значение
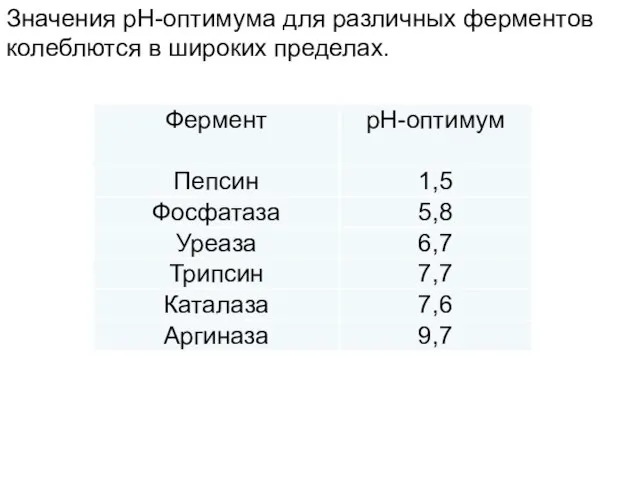
Слайд 46Значения рН-оптимума для различных ферментов колеблются в широких пределах.
Значения рН-оптимума для различных ферментов колеблются в широких пределах.
Слайд 47Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает
Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает

Слайд 48Ингибирование ферментов
Скорость ферментативной реакции может быть снижена при участиии ингибиторов.
Ингибитор –
Ингибирование ферментов
Скорость ферментативной реакции может быть снижена при участиии ингибиторов.
Ингибитор –

Слайд 49Ингибиторы можно разделить на два основных типа: необратимые и обратимые.
Необратимые ингибиторы
Необратимые ингибиторы

Слайд 50
Примером необратимого ингибитора является диизопропилфторфосфат (ДФФ). ДФФ ингибирует фермент ацетилхолинэстеразу, играющего важную
Примером необратимого ингибитора является диизопропилфторфосфат (ДФФ). ДФФ ингибирует фермент ацетилхолинэстеразу, играющего важную

Слайд 51Обратимые ингибиторы при определенных условиях могут быть легко отделены от фермента.
Активность
Активность

Слайд 52Типы ингибирования
Конкурентное ингибирование.
Конкурентный ингибитор, являясь структурным аналогом субстрата, взаимодействует с активным центром
Типы ингибирования
Конкурентное ингибирование.
Конкурентный ингибитор, являясь структурным аналогом субстрата, взаимодействует с активным центром

Слайд 53
Конкурентные ингибиторы используются в медицине. Для борьбы с инфекционными болезнями ранее широко
Конкурентные ингибиторы используются в медицине. Для борьбы с инфекционными болезнями ранее широко
Слайд 54В присутствии конкурентного ингибитора
V максимальная = const,
Kм увеличивается
В присутствии конкурентного ингибитора
V максимальная = const,
Kм увеличивается
Слайд 55Неконкурентное ингибирование
Неконкурентные ингибиторы по структуре не сходны с субстратом и при образовании
Неконкурентное ингибирование
Неконкурентные ингибиторы по структуре не сходны с субстратом и при образовании

Слайд 56
В качестве неконкурентного ингибитора может выступать цианид CN-. Он связывается с ионами
В качестве неконкурентного ингибитора может выступать цианид CN-. Он связывается с ионами
Слайд 57В присутствии неконкурентного ингибитора
V максимальная уменьшается,
Kм = const
В присутствии неконкурентного ингибитора
V максимальная уменьшается,
Kм = const
Слайд 58Бесконкурентное ингибирование
Ингибитор обратимо взаимодействует с ферментом только после образования фермент-субстратного комплекса.
Образующийся в
Ингибитор обратимо взаимодействует с ферментом только после образования фермент-субстратного комплекса.
Образующийся в

Слайд 59Смешанное ингибирование
сочетает в себе конкурентное и неконкурентное торможение.
Ингибитор, присоединяется в
Смешанное ингибирование
сочетает в себе конкурентное и неконкурентное торможение.
Ингибитор, присоединяется в

Слайд 60Регуляция каталитической активности ферментов
Регуляция путём посттрансляционной ковалентной модификации молекулы фермента (ограниченный протеолиз,
Регуляция каталитической активности ферментов
Регуляция путём посттрансляционной ковалентной модификации молекулы фермента (ограниченный протеолиз,

Слайд 61Регуляция функционирования ферментных систем
Ферментные системы обладают способностью поддерживать необходимую скорость суммарного процесса
Регуляция функционирования ферментных систем
Ферментные системы обладают способностью поддерживать необходимую скорость суммарного процесса

Слайд 62Регуляция биосинтеза ферментов
Индукция фермента (синтез de novo) происходит при повышении концентрации субстрата
Регуляция биосинтеза ферментов
Индукция фермента (синтез de novo) происходит при повышении концентрации субстрата

Слайд 63Аллостерические ферменты
Термин «аллостерический» происходит от греческих слов allo – другой, stereo –
Аллостерические ферменты
Термин «аллостерический» происходит от греческих слов allo – другой, stereo –
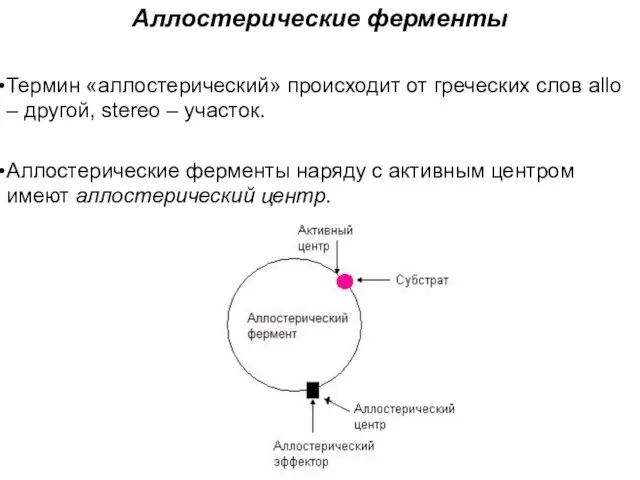
Слайд 64С аллостерическим центром связываются аллостерические эффекторы - вещества, способные изменять активность ферментов.
С аллостерическим центром связываются аллостерические эффекторы - вещества, способные изменять активность ферментов.

Слайд 65Регуляция мультиферментных систем
Некоторые ферменты действуют согласованно, объединяясь в мультиферментные системы, в которых
Регуляция мультиферментных систем
Некоторые ферменты действуют согласованно, объединяясь в мультиферментные системы, в которых

Слайд 66В мультиферментной системе есть фермент, который определяет скорость всей последовательности реакций.
Этот
В мультиферментной системе есть фермент, который определяет скорость всей последовательности реакций.
Этот

Слайд 67Применение ферментов
Отрасли промышленности, в которых применяются ферменты:
хлебопечение, пивоварение, виноделие, чайное, кожевенное и
Применение ферментов
Отрасли промышленности, в которых применяются ферменты:
хлебопечение, пивоварение, виноделие, чайное, кожевенное и

Слайд 68Применение ферментов в медицине
Энзимопатология - изучает молекулярные основы развития патологического процесса, исходя
Применение ферментов в медицине
Энзимопатология - изучает молекулярные основы развития патологического процесса, исходя

Слайд 69Энзимотерапия – использование ферментов, активаторов и ингибиторов в качестве лекарственных средств.
Например: протеолитические
Энзимотерапия – использование ферментов, активаторов и ингибиторов в качестве лекарственных средств.
Например: протеолитические

 Презентация на тему Биологические ритмы организма человека
Презентация на тему Биологические ритмы организма человека  Презентация на тему Глаз как оптическая система (8 класс)
Презентация на тему Глаз как оптическая система (8 класс)  Спинной и спинномозговые нервы
Спинной и спинномозговые нервы Побег. Внешнее строение побега
Побег. Внешнее строение побега Жизненный цикл клетки
Жизненный цикл клетки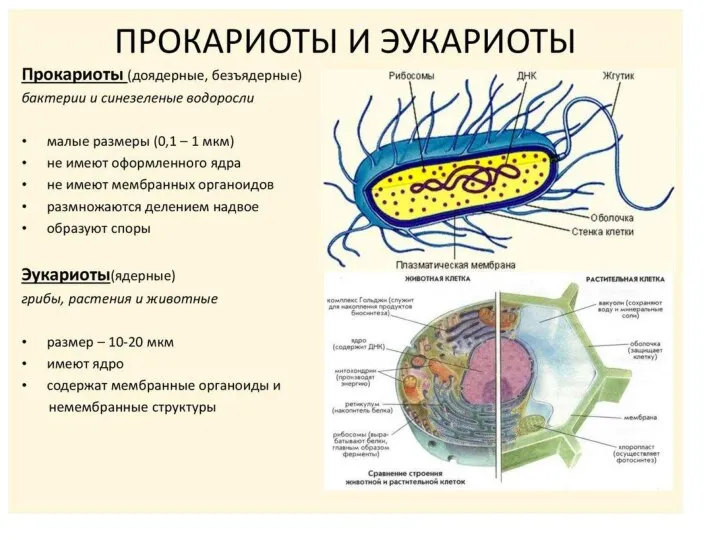 Прокариоты и эукариоты
Прокариоты и эукариоты What are they thinking or saying Warm up
What are they thinking or saying Warm up Генетика. Законы Менделя. Закон Моргана. Аллели. Группы крови
Генетика. Законы Менделя. Закон Моргана. Аллели. Группы крови Презентация на тему Развитие жизни в кайнозойскую эру
Презентация на тему Развитие жизни в кайнозойскую эру  Презентация на тему Почвенная среда обитания
Презентация на тему Почвенная среда обитания 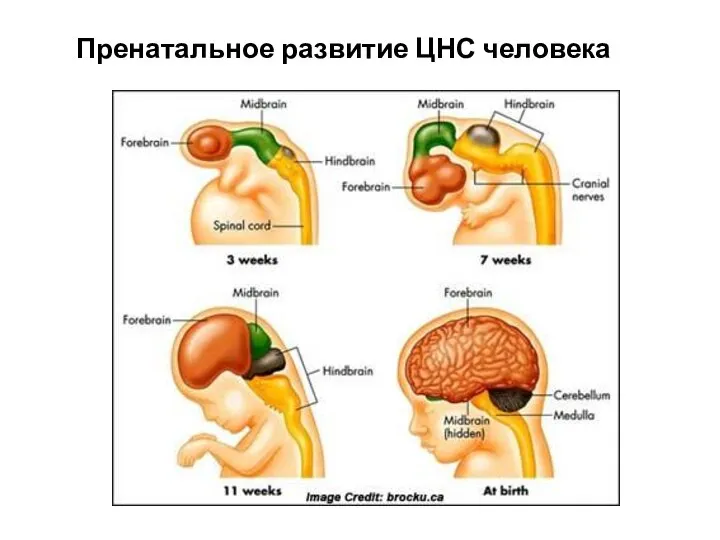 Пренатальное развитие центральной нервной системы человека (ЦНС)
Пренатальное развитие центральной нервной системы человека (ЦНС) Витамины. Классификация
Витамины. Классификация Презентация на тему КЛАССИФИКАЦИЯ МОЛЛЮСКОВ
Презентация на тему КЛАССИФИКАЦИЯ МОЛЛЮСКОВ  Молекулярная биология. Лекция 3
Молекулярная биология. Лекция 3 Обитатели Тайги
Обитатели Тайги Обитатели тайги
Обитатели тайги Аэробный этап энергетического обмена
Аэробный этап энергетического обмена Амеба обыкновенная
Амеба обыкновенная Проект: Зимующие птицы нашего города
Проект: Зимующие птицы нашего города Молекулярные основы наследственности
Молекулярные основы наследственности Корень, его строение и значение
Корень, его строение и значение Презентация на тему Роль гормонов в обмене веществ, росте и развитии организма
Презентация на тему Роль гормонов в обмене веществ, росте и развитии организма  Гемоглобин
Гемоглобин Одомашнивание оленей
Одомашнивание оленей Значение леса. Охрана леса
Значение леса. Охрана леса Головной мозг
Головной мозг Морковь
Морковь Комнатные птицы
Комнатные птицы