- Ферменты. Часть 1

Содержание
- 2. ФЕРМЕНТЫ (энзимы) - это высокоспецифичные белки, выполняющие функции биологических катализаторов. Катализатор - это вещество, которое ускоряет
- 3. Ферменты могут быть простыми или сложными белками. Если фермент - сложный белок, то его небелковую часть
- 4. Роль ионов металлов: Zn2+, Mg2+, Fe2+, Cu2+ и т.д. Механизмы участия ионов металлов в ферментативных реакциях
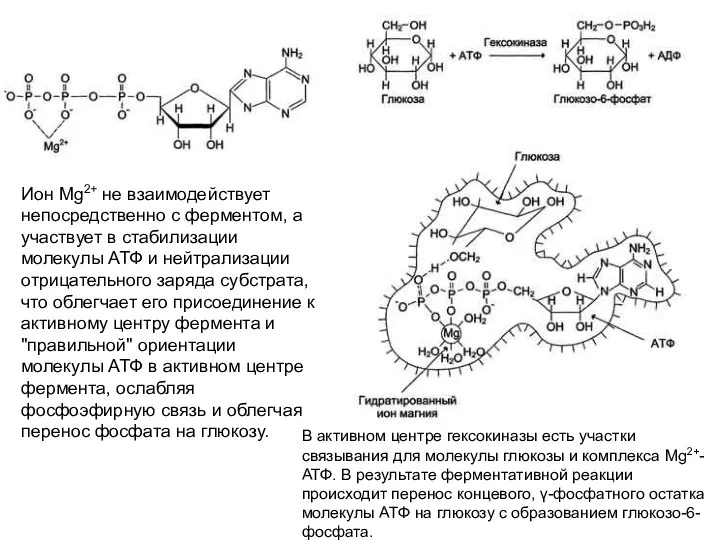
- 5. Ион Mg2+ не взаимодействует непосредственно с ферментом, а участвует в стабилизации молекулы АТФ и нейтрализации отрицательного
- 6. Поскольку ферменты - белковые молекулы, следовательно, они обладают всеми свойствами, характерными для белков. В то же
- 7. Активный центр ферментов - это определенный участок белковой молекулы, способный комплементарно связываться с субстратом и обеспечивающий
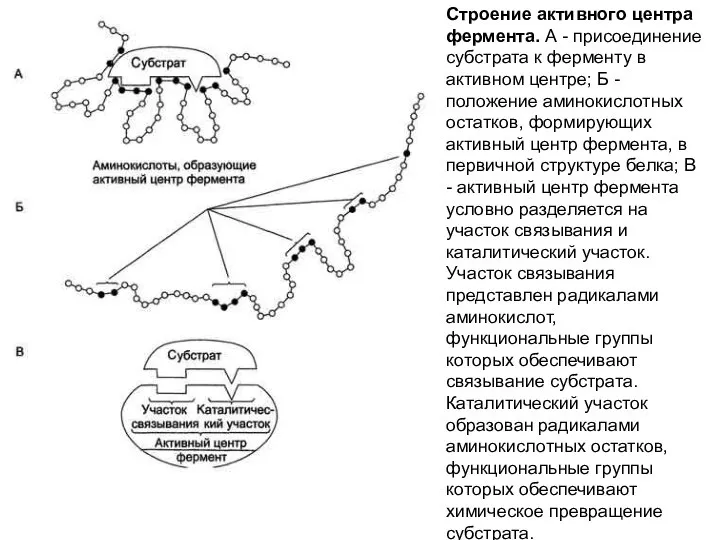
- 8. Строение активного центра фермента. А - присоединение субстрата к ферменту в активном центре; Б - положение
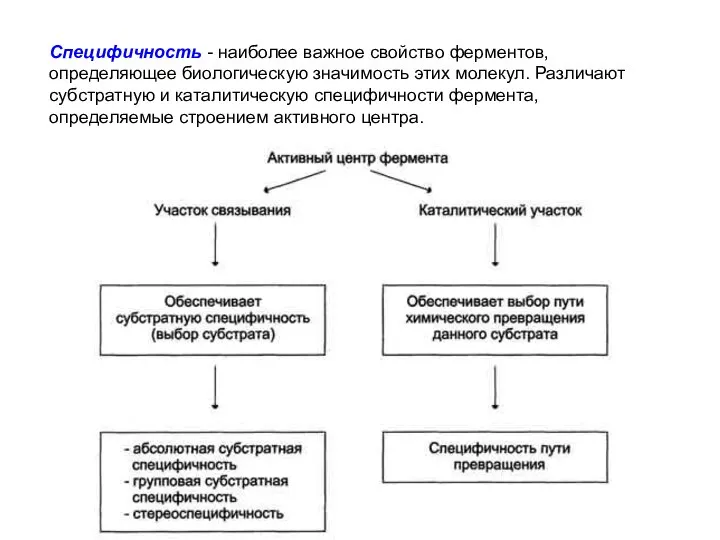
- 9. Специфичность - наиболее важное свойство ферментов, определяющее биологическую значимость этих молекул. Различают субстратную и каталитическую специфичности
- 10. Субстратная специфичность Под субстратной специфичностью понимают способность каждого фермента взаимодействовать лишь с одним или несколькими определёнными
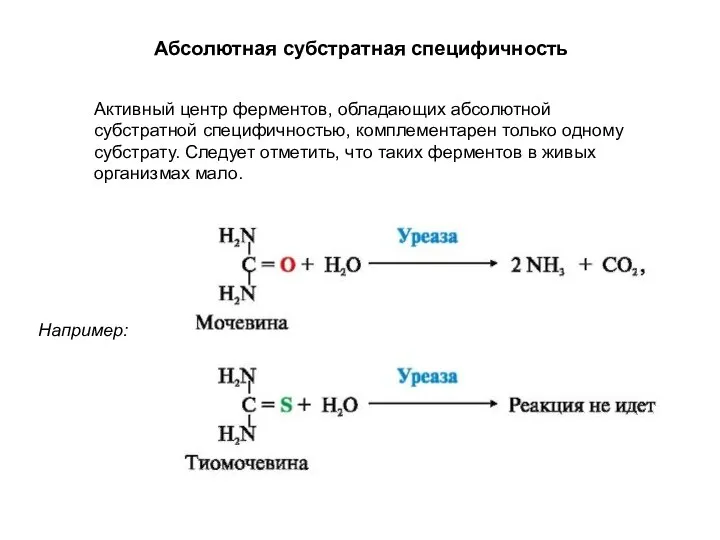
- 11. Активный центр ферментов, обладающих абсолютной субстратной специфичностью, комплементарен только одному субстрату. Следует отметить, что таких ферментов
- 12. Групповая субстратная специфичность Большинство ферментов катализирует однотипные реакции с небольшим количеством (группой) структурно похожих субстратов. Например:
- 13. Стереоспецифичность При наличии у субстрата нескольких стереоизомеров фермент проявляет абсолютную специфичность к одному из них. В
- 14. Каталитическая специфичность Фермент катализирует превращение присоединённого субстрата по одному из возможных путей его превращения. Это свойство
- 15. Для ферментов характерны: • специфичность. • каталитическая эффективность. Большинство катализируемых ферментами реакций высокоэффективны, они протекают в
- 16. • Активность ферментов может регулироваться. Действие ферментов в клетке, как правило, строго упорядочено: продукт одной ферментативной
- 17. ОСОБЕННОСТИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
- 18. Условия, необходимы для химического взаимодействия молекул, чтобы произошла химическая реакция: 1) Молекулы должны сблизиться (столкнуться). Но
- 19. При достижении этого энергетического барьера в молекуле происходят изменения, вызывающие перераспределение химических связей и образование новых
- 20. Для превращения субстрата в продукт необходимо, чтобы молекула субстрата преодолела определенный энергетический барьер (энергия активации) Кирюхин
- 21. Чем больше молекул обладает энергией, превышающей уровень Еа, тем выше скорость химической реакции. Повысить скорость химической
- 22. Энергия активации Кирюхин Д.О.
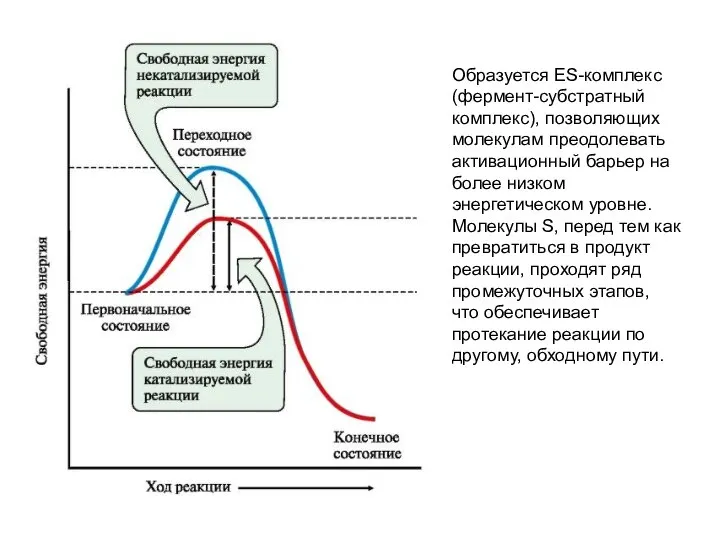
- 23. Образуется ES-комплекс (фермент-субстратный комплекс), позволяющих молекулам преодолевать активационный барьер на более низком энергетическом уровне. Молекулы S,
- 24. • На первом этапе фермент взаимодействует с субстратом с образованием нового соединения - ES E +
- 25. Иначе говоря, катализатор открывает новый, дополнительный путь превращения исходных веществ в продукты реакции через такие состояния,
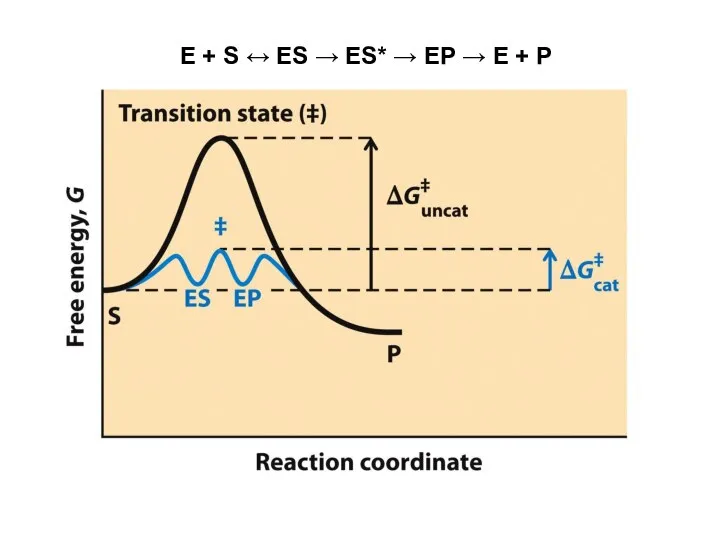
- 26. E + S ↔ ES → ES* → EP → E + P
- 27. 1 – некатализируемая реакция 2 – катализируемая реакция
- 28. ЭНЕРГЕТИЧЕСКИЙ ИТОГ РЕАКЦИИ - разность между энергетическим уровнем исходных веществ (субстратов) и энергетическим уровнем продуктов реакции,
- 29. Теории узнавания ферментом субстрата
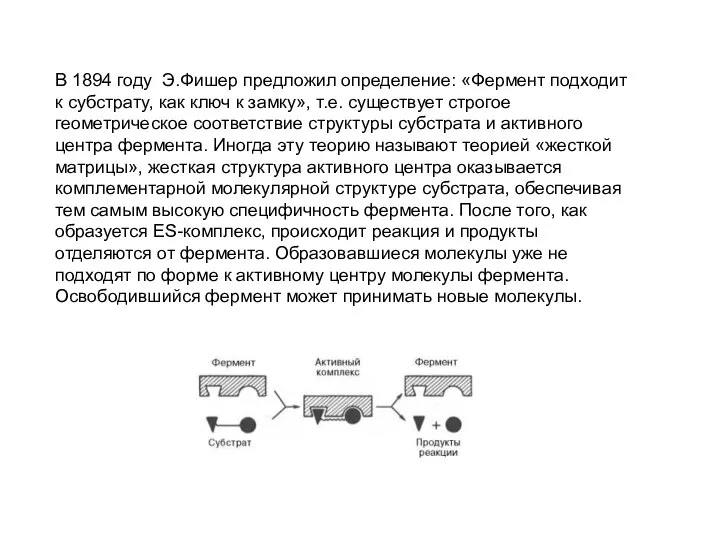
- 30. В 1894 году Э.Фишер предложил определение: «Фермент подходит к субстрату, как ключ к замку», т.е. существует
- 31. В противоположность модели Э. Фишера Д. Кошлендом в 50-е годы 20 века была разработана теория «индуцированного
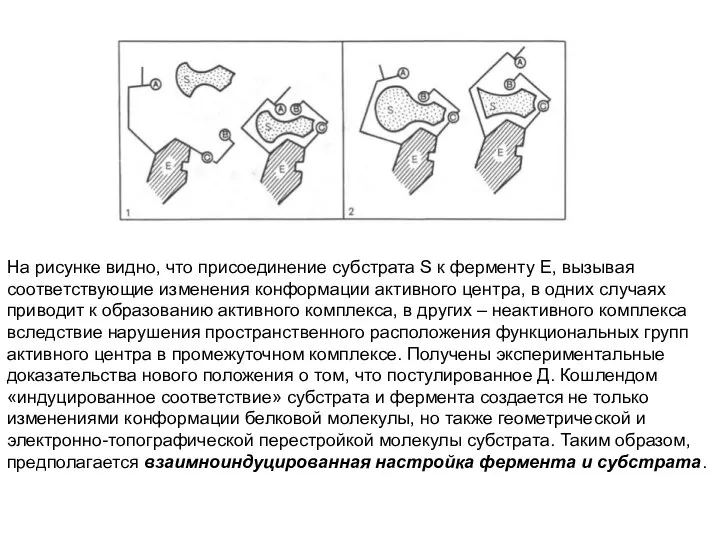
- 32. На рисунке видно, что присоединение субстрата S к ферменту Е, вызывая соответствующие изменения конформации активного центра,
- 33. КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
- 34. Кинетика ферментативных реакций - это раздел энзимологии, изучающий зависимость скорости химических реакций, катализируемых ферментами, от химической
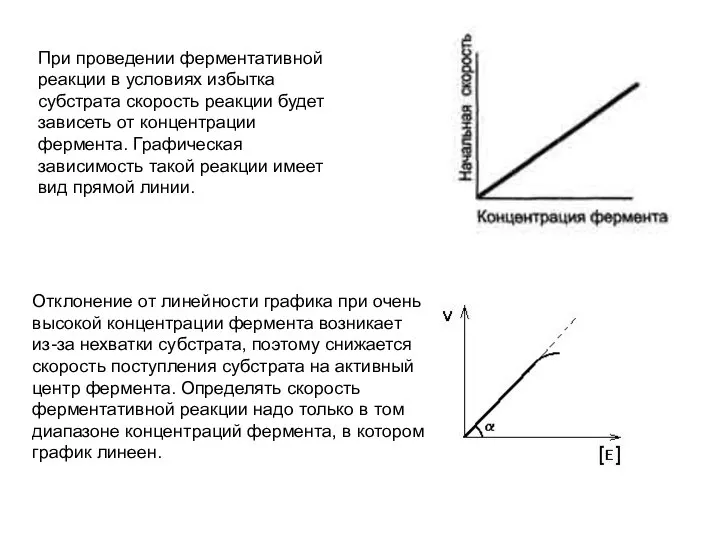
- 35. При проведении ферментативной реакции в условиях избытка субстрата скорость реакции будет зависеть от концентрации фермента. Графическая
- 36. Количество фермента часто невозможно определить в абсолютных величинах, поэтому о количестве ферментов судят по скорости катализируемой
- 37. Для выражения концентрации фермента и количественной оценки его активности в условных единицах Комиссией по ферментам Международного
- 38. В медицине активность ферментов выражают чаще всего в единицах активности на 1 л биологической жидкости либо
- 39. В простейшем случае ферментативную реакцию можно представить как двухстадийный процесс. Зависимость скорости реакции v от концентрации
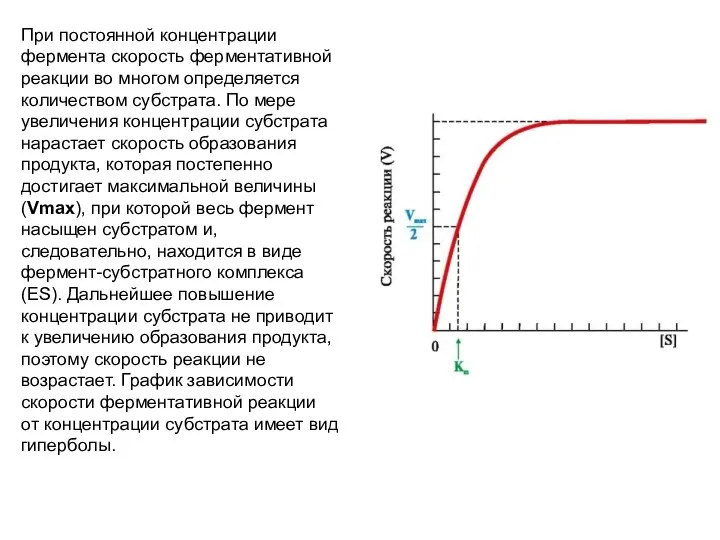
- 40. При постоянной концентрации фермента скорость ферментативной реакции во многом определяется количеством субстрата. По мере увеличения концентрации
- 41. Когда половина молекул фермента находится в комплексе с молекулами субстрата, скорость реакции равна 1/2 Vmax. Концентрация
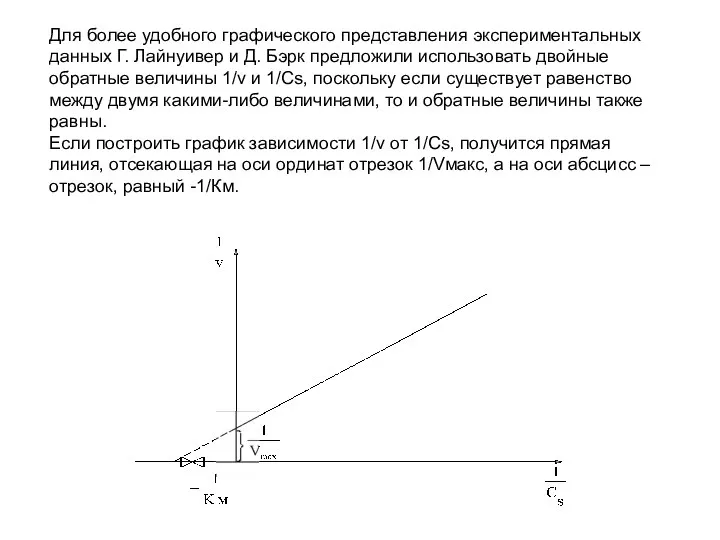
- 42. Для более удобного графического представления экспериментальных данных Г. Лайнуивер и Д. Бэрк предложили использовать двойные обратные
- 43. Влияние температуры на скорость ферментативной реакции Скорость ферментативных реакций, как и любых химических реакций, зависит от
- 44. Влияние pH на активность ферментов Зависимость скорости ферментативной реакции от рН имеет колоколообразный вид. Значения рН,
- 46. Отклонение рН среды от оптимального вызывает изменение: - ионизации функциональных групп фермента, а иногда и субстрата;
- 48. Скачать презентацию
Слайд 2 ФЕРМЕНТЫ (энзимы) - это высокоспецифичные белки, выполняющие функции биологических катализаторов.
Катализатор -
ФЕРМЕНТЫ (энзимы) - это высокоспецифичные белки, выполняющие функции биологических катализаторов.
Катализатор -

Слайд 3Ферменты могут быть простыми или сложными белками. Если фермент - сложный белок,
Ферменты могут быть простыми или сложными белками. Если фермент - сложный белок,

Слайд 4Роль ионов металлов: Zn2+, Mg2+, Fe2+, Cu2+ и т.д.
Механизмы участия ионов
Роль ионов металлов: Zn2+, Mg2+, Fe2+, Cu2+ и т.д.
Механизмы участия ионов

Слайд 5Ион Mg2+ не взаимодействует непосредственно с ферментом, а участвует в стабилизации молекулы
Ион Mg2+ не взаимодействует непосредственно с ферментом, а участвует в стабилизации молекулы
Слайд 6Поскольку ферменты - белковые молекулы, следовательно, они обладают всеми свойствами, характерными для
Поскольку ферменты - белковые молекулы, следовательно, они обладают всеми свойствами, характерными для

Слайд 7Активный центр ферментов - это определенный участок белковой молекулы, способный комплементарно связываться
Активный центр ферментов - это определенный участок белковой молекулы, способный комплементарно связываться

Слайд 8Строение активного центра фермента. А - присоединение субстрата к ферменту в активном
Строение активного центра фермента. А - присоединение субстрата к ферменту в активном
Слайд 9Специфичность - наиболее важное свойство ферментов, определяющее биологическую значимость этих молекул. Различают
Специфичность - наиболее важное свойство ферментов, определяющее биологическую значимость этих молекул. Различают
Слайд 10Субстратная специфичность
Под субстратной специфичностью понимают способность каждого фермента взаимодействовать лишь с
Субстратная специфичность
Под субстратной специфичностью понимают способность каждого фермента взаимодействовать лишь с

Слайд 11Активный центр ферментов, обладающих абсолютной субстратной специфичностью, комплементарен только одному субстрату. Следует
Активный центр ферментов, обладающих абсолютной субстратной специфичностью, комплементарен только одному субстрату. Следует
Слайд 12Групповая субстратная специфичность
Большинство ферментов катализирует однотипные реакции с небольшим количеством (группой)
Групповая субстратная специфичность
Большинство ферментов катализирует однотипные реакции с небольшим количеством (группой)

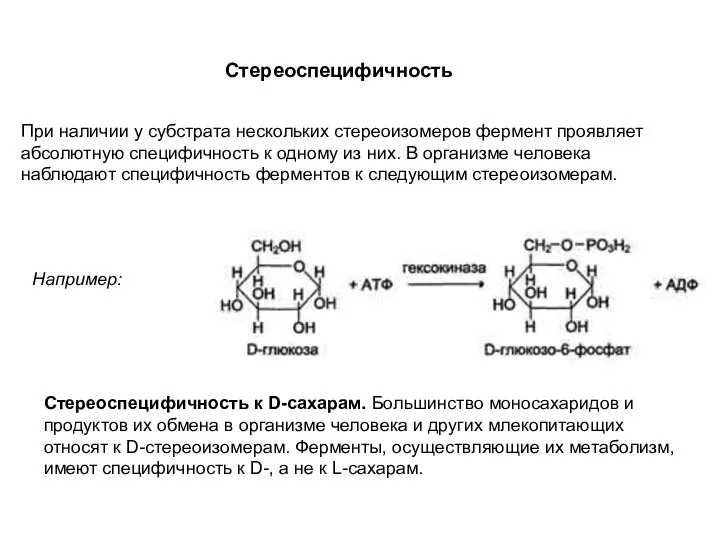
Слайд 13Стереоспецифичность
При наличии у субстрата нескольких стереоизомеров фермент проявляет абсолютную специфичность к
Стереоспецифичность
При наличии у субстрата нескольких стереоизомеров фермент проявляет абсолютную специфичность к
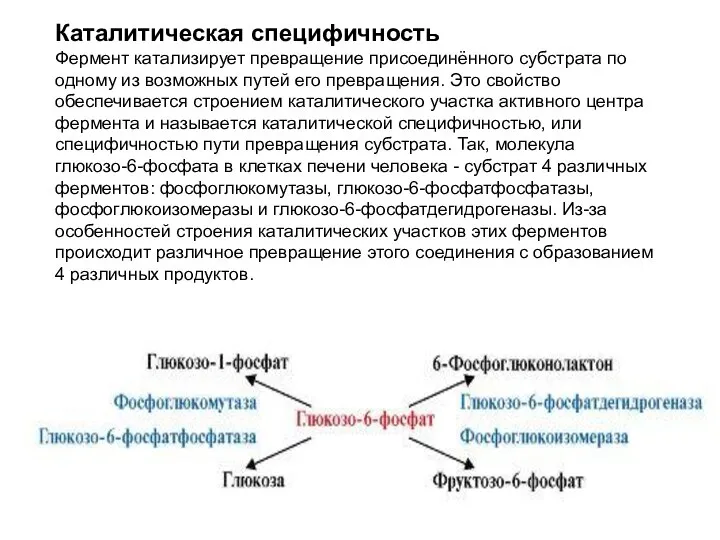
Слайд 14Каталитическая специфичность
Фермент катализирует превращение присоединённого субстрата по одному из возможных путей
Каталитическая специфичность
Фермент катализирует превращение присоединённого субстрата по одному из возможных путей
Слайд 15Для ферментов характерны:
• специфичность.
• каталитическая эффективность. Большинство катализируемых ферментами реакций высокоэффективны, они
Для ферментов характерны:
• специфичность.
• каталитическая эффективность. Большинство катализируемых ферментами реакций высокоэффективны, они

Слайд 16• Активность ферментов может регулироваться. Действие ферментов в клетке, как правило, строго
• Активность ферментов может регулироваться. Действие ферментов в клетке, как правило, строго

Слайд 17ОСОБЕННОСТИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
ОСОБЕННОСТИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА

Слайд 18Условия, необходимы для химического взаимодействия молекул, чтобы произошла химическая реакция:
1) Молекулы
Условия, необходимы для химического взаимодействия молекул, чтобы произошла химическая реакция:
1) Молекулы

Слайд 19При достижении этого энергетического барьера в молекуле происходят изменения, вызывающие перераспределение химических
При достижении этого энергетического барьера в молекуле происходят изменения, вызывающие перераспределение химических

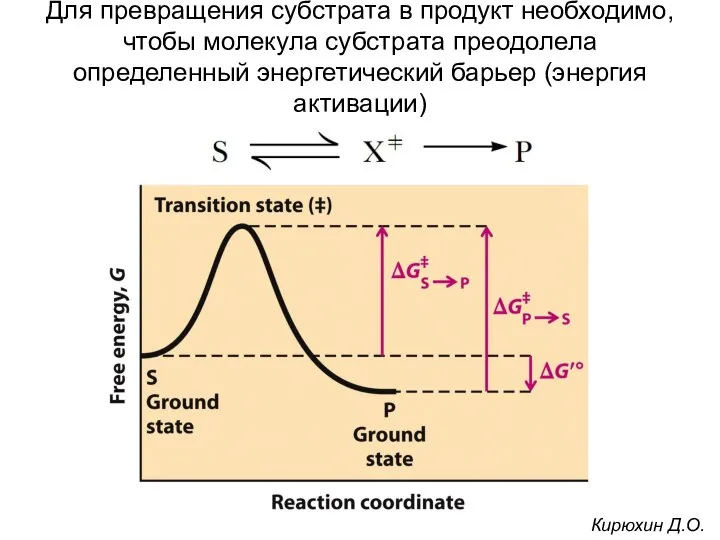
Слайд 20Для превращения субстрата в продукт необходимо, чтобы молекула субстрата преодолела определенный энергетический
Для превращения субстрата в продукт необходимо, чтобы молекула субстрата преодолела определенный энергетический
Слайд 21Чем больше молекул обладает энергией, превышающей уровень Еа, тем выше скорость химической
Чем больше молекул обладает энергией, превышающей уровень Еа, тем выше скорость химической

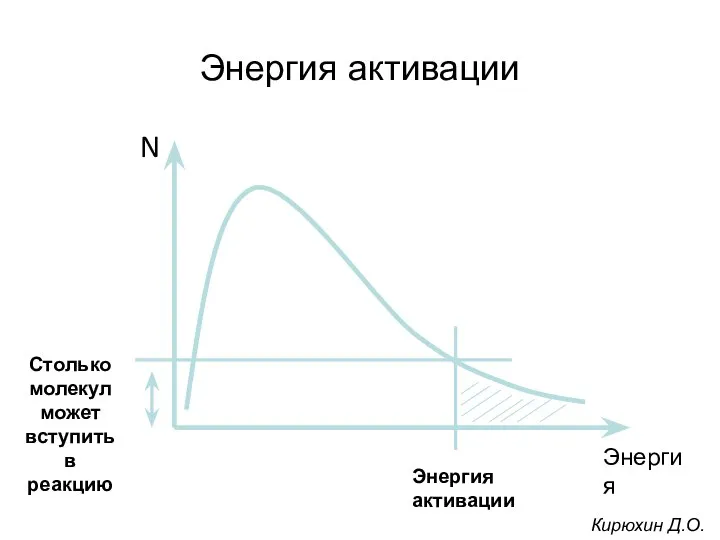
Слайд 22Энергия активации
Кирюхин Д.О.
Энергия активации
Кирюхин Д.О.
Слайд 23Образуется ES-комплекс (фермент-субстратный комплекс), позволяющих молекулам преодолевать активационный барьер на более низком
Образуется ES-комплекс (фермент-субстратный комплекс), позволяющих молекулам преодолевать активационный барьер на более низком
Слайд 24• На первом этапе фермент взаимодействует с субстратом с образованием нового соединения
• На первом этапе фермент взаимодействует с субстратом с образованием нового соединения

Слайд 25Иначе говоря, катализатор открывает новый, дополнительный путь превращения исходных веществ в продукты
Иначе говоря, катализатор открывает новый, дополнительный путь превращения исходных веществ в продукты

Слайд 26E + S ↔ ES → ES* → EP → E +
E + S ↔ ES → ES* → EP → E +
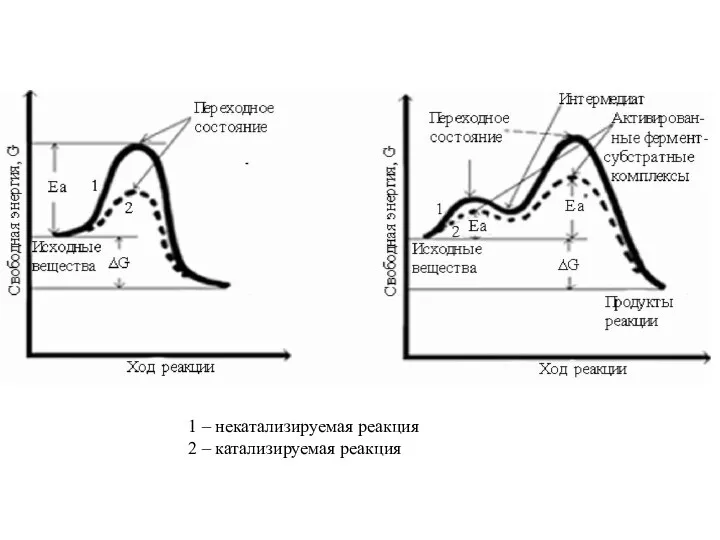
Слайд 271 – некатализируемая реакция
2 – катализируемая реакция
1 – некатализируемая реакция
2 – катализируемая реакция
Слайд 28ЭНЕРГЕТИЧЕСКИЙ ИТОГ РЕАКЦИИ - разность между энергетическим уровнем исходных веществ (субстратов) и
ЭНЕРГЕТИЧЕСКИЙ ИТОГ РЕАКЦИИ - разность между энергетическим уровнем исходных веществ (субстратов) и

Слайд 29Теории узнавания ферментом субстрата
Теории узнавания ферментом субстрата

Слайд 30В 1894 году Э.Фишер предложил определение: «Фермент подходит к субстрату, как ключ
В 1894 году Э.Фишер предложил определение: «Фермент подходит к субстрату, как ключ
Слайд 31В противоположность модели Э. Фишера Д. Кошлендом в 50-е годы 20 века
В противоположность модели Э. Фишера Д. Кошлендом в 50-е годы 20 века

Слайд 32На рисунке видно, что присоединение субстрата S к ферменту Е, вызывая соответствующие
На рисунке видно, что присоединение субстрата S к ферменту Е, вызывая соответствующие
Слайд 33КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ

Слайд 34Кинетика ферментативных реакций - это раздел энзимологии, изучающий зависимость скорости химических реакций,
Кинетика ферментативных реакций - это раздел энзимологии, изучающий зависимость скорости химических реакций,

Слайд 35При проведении ферментативной реакции в условиях избытка субстрата скорость реакции будет зависеть
При проведении ферментативной реакции в условиях избытка субстрата скорость реакции будет зависеть
Слайд 36Количество фермента часто невозможно определить в абсолютных величинах, поэтому о количестве ферментов
Количество фермента часто невозможно определить в абсолютных величинах, поэтому о количестве ферментов

Слайд 37Для выражения концентрации фермента и количественной оценки его активности в условных единицах
Для выражения концентрации фермента и количественной оценки его активности в условных единицах

Слайд 38В медицине активность ферментов выражают чаще всего в единицах активности на 1 л
В медицине активность ферментов выражают чаще всего в единицах активности на 1 л

Слайд 39В простейшем случае ферментативную реакцию можно представить как двухстадийный процесс.
Зависимость скорости
В простейшем случае ферментативную реакцию можно представить как двухстадийный процесс.
Зависимость скорости

Слайд 40При постоянной концентрации фермента скорость ферментативной реакции во многом определяется количеством субстрата.
При постоянной концентрации фермента скорость ферментативной реакции во многом определяется количеством субстрата.
Слайд 41Когда половина молекул фермента находится в комплексе с молекулами субстрата, скорость реакции
Когда половина молекул фермента находится в комплексе с молекулами субстрата, скорость реакции

Слайд 42Для более удобного графического представления экспериментальных данных Г. Лайнуивер и Д. Бэрк
Для более удобного графического представления экспериментальных данных Г. Лайнуивер и Д. Бэрк
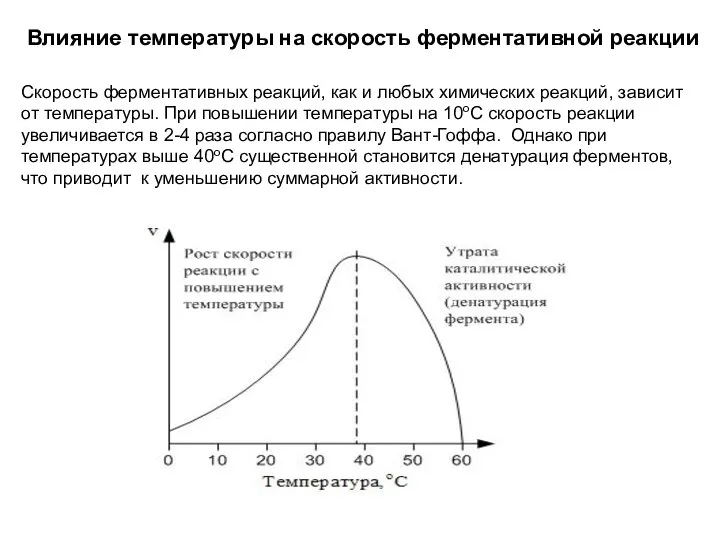
Слайд 43Влияние температуры на скорость ферментативной реакции
Скорость ферментативных реакций, как и любых химических
Влияние температуры на скорость ферментативной реакции
Скорость ферментативных реакций, как и любых химических
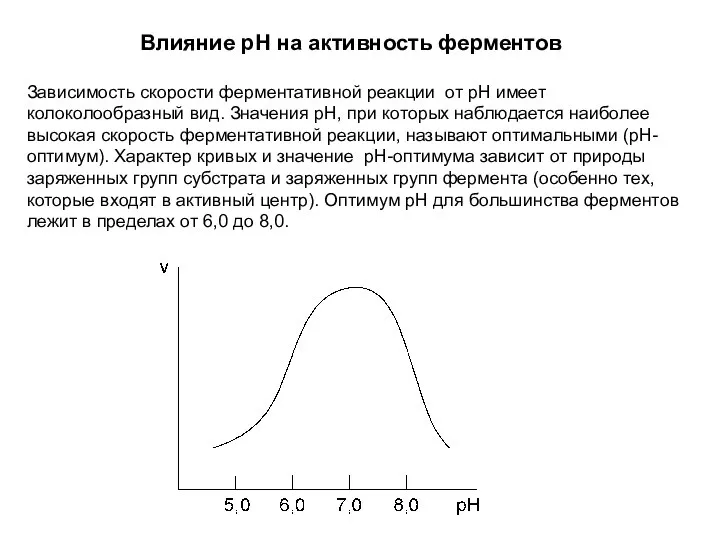
Слайд 44Влияние pH на активность ферментов
Зависимость скорости ферментативной реакции от рН имеет колоколообразный
Влияние pH на активность ферментов
Зависимость скорости ферментативной реакции от рН имеет колоколообразный

Слайд 46Отклонение рН среды от оптимального вызывает изменение:
- ионизации функциональных групп фермента,
Отклонение рН среды от оптимального вызывает изменение:
- ионизации функциональных групп фермента,

 Генетическая структура бактерий
Генетическая структура бактерий Перед.тех
Перед.тех Органы растений, участвующие в транспорте веществ Корни и стебель. Ксилема и флоэма
Органы растений, участвующие в транспорте веществ Корни и стебель. Ксилема и флоэма Физическое развитие собак
Физическое развитие собак Микологические исследования съедобных грибов на территории Сыктывдинского лесничества
Микологические исследования съедобных грибов на территории Сыктывдинского лесничества Презентация на тему "Деление цветковых растений на однодольные и двудольные. Характерные признаки" - презентации по Биологии
Презентация на тему "Деление цветковых растений на однодольные и двудольные. Характерные признаки" - презентации по Биологии Зрительный анализатор
Зрительный анализатор Критерии живых систем
Критерии живых систем Морфология насекомых. Голова и ее придатки
Морфология насекомых. Голова и ее придатки Насекомоядные животные
Насекомоядные животные Og|hidrāti un to nozīme. Monosaharīdi
Og|hidrāti un to nozīme. Monosaharīdi Движущий и направляющий фактор эволюции
Движущий и направляющий фактор эволюции Для чего клетка принимает кислород?
Для чего клетка принимает кислород? Ткани человека. 4
Ткани человека. 4 Типы вегетативного размножения комнатных растений
Типы вегетативного размножения комнатных растений Презентация на тему Голосеменные
Презентация на тему Голосеменные  Класс Птицы. Общая характеристика птиц. Внешнее строение птиц
Класс Птицы. Общая характеристика птиц. Внешнее строение птиц Глюкоза. Строение глюкозы
Глюкоза. Строение глюкозы Парамфистомоз. Дикроцелиоз. Эуритрематоз
Парамфистомоз. Дикроцелиоз. Эуритрематоз Растительный мир. Игра
Растительный мир. Игра Классификация витаминов. Витаминоподобные соединения
Классификация витаминов. Витаминоподобные соединения Живой мир
Живой мир Парк Горни
Парк Горни Лесные ресурсы
Лесные ресурсы Удельная продукция. Р/В коэффициент. Ассимиляционное число
Удельная продукция. Р/В коэффициент. Ассимиляционное число Деревья и плоды
Деревья и плоды Свёрточные нейронные сети. Часть 1: Основные понятия, архитектура
Свёрточные нейронные сети. Часть 1: Основные понятия, архитектура Разнообразие животных
Разнообразие животных