- Ферменты модификации ДНК

Содержание
- 2. ФЕРМЕНТЫ МОДИФИКАЦИИ ДНК Ферменты модификации ДНК в настоящее время научились синтезировать искусственно и применять для воздействия
- 3. ЛИГАЗЫ ДНК-лигазы вирусов, бактерий, млекопитающих соединяют 5'-фосфатную и З'-гидроксильную группы нуклеотидов, находящихся на противоположных концах одноцепочечного
- 4. ФОСФАТАЗЫ Это фермент гидролаза, отщепляющая фосфат от нуклеотидов, белков и алкалоидов. Наиболее часто щелочная фосфатаза применяется
- 5. КИНАЗЫ Полинуклеотидкиназа катализирует передачу гамма-фосфата из АТФ в 5'-ОН-группу одно- и двухцепочечных ДНК и РНК, олигонуклеотидов
- 6. Т4 ДНК-ПОЛИМЕРАЗА ДНК-полимераза T4 - матрично-зависимая ДНК-полимераза, которая катализирует синтез 5'-3 'из праймированной одноцепочечной ДНК. Фермент
- 7. ФРАГМЕНТ КЛЁНОВА Это большой белковый фрагмент, образующийся при ферментативном расщеплении ДНК-полимеразы I из Escherichia coli протеазой
- 9. Скачать презентацию
Слайд 2ФЕРМЕНТЫ МОДИФИКАЦИИ ДНК
Ферменты модификации ДНК в настоящее время научились синтезировать искусственно и
ФЕРМЕНТЫ МОДИФИКАЦИИ ДНК
Ферменты модификации ДНК в настоящее время научились синтезировать искусственно и

Слайд 3ЛИГАЗЫ
ДНК-лигазы вирусов, бактерий, млекопитающих соединяют 5'-фосфатную и З'-гидроксильную группы нуклеотидов, находящихся на
ЛИГАЗЫ
ДНК-лигазы вирусов, бактерий, млекопитающих соединяют 5'-фосфатную и З'-гидроксильную группы нуклеотидов, находящихся на

Слайд 4ФОСФАТАЗЫ
Это фермент гидролаза, отщепляющая фосфат от нуклеотидов, белков и алкалоидов.
Наиболее часто щелочная
ФОСФАТАЗЫ
Это фермент гидролаза, отщепляющая фосфат от нуклеотидов, белков и алкалоидов.
Наиболее часто щелочная

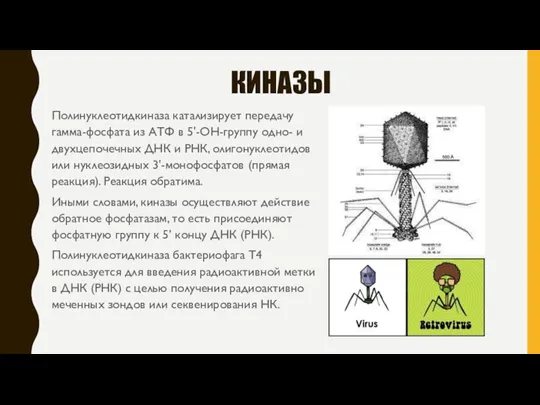
Слайд 5КИНАЗЫ
Полинуклеотидкиназа катализирует передачу гамма-фосфата из АТФ в 5'-ОН-группу одно- и двухцепочечных ДНК
КИНАЗЫ
Полинуклеотидкиназа катализирует передачу гамма-фосфата из АТФ в 5'-ОН-группу одно- и двухцепочечных ДНК
Слайд 6Т4 ДНК-ПОЛИМЕРАЗА
ДНК-полимераза T4 - матрично-зависимая ДНК-полимераза, которая катализирует синтез 5'-3 'из праймированной
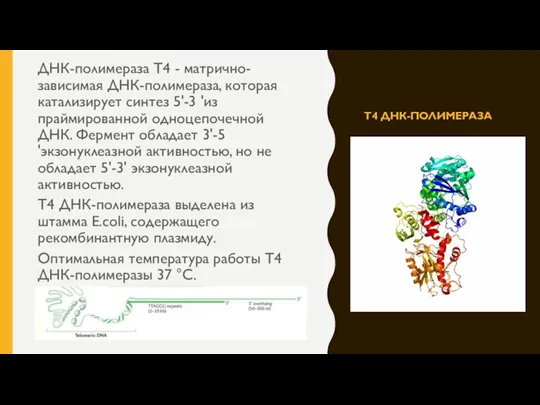
Т4 ДНК-ПОЛИМЕРАЗА
ДНК-полимераза T4 - матрично-зависимая ДНК-полимераза, которая катализирует синтез 5'-3 'из праймированной
Слайд 7ФРАГМЕНТ КЛЁНОВА
Это большой белковый фрагмент, образующийся при ферментативном расщеплении ДНК-полимеразы I из
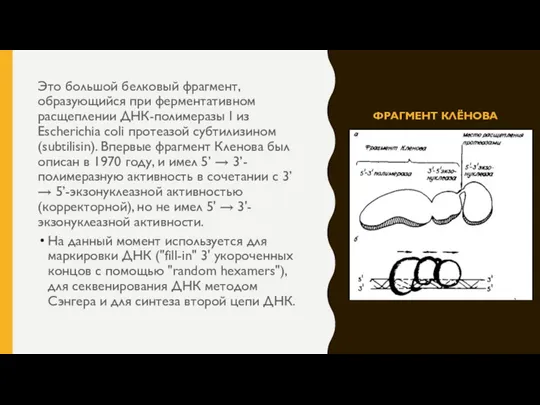
ФРАГМЕНТ КЛЁНОВА
Это большой белковый фрагмент, образующийся при ферментативном расщеплении ДНК-полимеразы I из
 Насекомые вредители на станции юных натуралистов г. Тайшета
Насекомые вредители на станции юных натуралистов г. Тайшета Рыбы и насекомые пресных водоемов
Рыбы и насекомые пресных водоемов Грибы, цветы и ягоды леса
Грибы, цветы и ягоды леса Чей хвостик
Чей хвостик Семейство крестоцветных
Семейство крестоцветных Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба
Строение клетки. Инфузория туфелька. Эвглена зеленая. Амеба Неклеточные формы жизни
Неклеточные формы жизни Раселение живых организмов
Раселение живых организмов Эпителий
Эпителий Презентация на тему Строение и химический состав клетки
Презентация на тему Строение и химический состав клетки  MOLEKUlyarnaya_biologia_LPZ_1
MOLEKUlyarnaya_biologia_LPZ_1 Фотодыхание
Фотодыхание Вирусология - наука о вирусах
Вирусология - наука о вирусах Кровь (физиология системы крови) - Презентация
Кровь (физиология системы крови) - Презентация Чабрец (богородская трава, тимьян ползучий)
Чабрец (богородская трава, тимьян ползучий) Составление генетических паспортов и установление родственных связей
Составление генетических паспортов и установление родственных связей От клюва до хвоста
От клюва до хвоста Модификационная изменчивость
Модификационная изменчивость Строение и многообразие грибов
Строение и многообразие грибов Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности
Проверка домашнего задания 1. рассказать о способах питания грибов и бактерий 2. рассказать о круговороте веществ и непрерывности  Что было на Земле, когда она появилась?
Что было на Земле, когда она появилась? Биохимия. Разделы биохимии. Становление биохимии как науки
Биохимия. Разделы биохимии. Становление биохимии как науки История собаководства
История собаководства Морфология бактерий
Морфология бактерий Жизненная Ёмкость Лёгких
Жизненная Ёмкость Лёгких Ткани животных
Ткани животных 2. Урок - .ДНК
2. Урок - .ДНК Лишайники
Лишайники