- Физиология сердца и сосудов

Содержание
- 2. Основные функции сердца: Автоматия Возбудимость Проводимость Сократимость
- 3. 1. Автоматия сердца
- 4. Автоматия сердца - это способность сердца к ритмическому сокращению без всяких видимых раздражений под влиянием импульсов,
- 5. Сердце человека сокращается: 70 ударов в минуту; 100 тысяч раз в сутки; 40 млн. в год;
- 6. АВТОМАТИЯ СЕРДЕЧНОЙ МЫШЦЫ: МИОГЕННАЯ – импульсы появляются в самих мышечных волокнах. Обеспечивает сокращения сердца на ранних
- 7. Проводящая система сердца синусный узел атриовентрикулярный узел пучок Гиса ножки пучка Гиса волокна Пуркинье
- 8. Частота активности элементов проводящей системы: синусно-предсердный узел (60-80 ударов в минуту). Пейсмекер первого порядка. Определяет активность
- 10. Закон градиента автоматизма сердца– у всех позвоночных степень автоматии тем выше, чем ближе расположен участок проводящей
- 11. 2. Природа автоматизма
- 12. Особенности клеток атипичной мышечной ткани: Только несколько клеток синусно-предсердного узла, называемых истинными пейсмекерами, обладают способностью к
- 13. Природа миогенного автоматизма. За генерацию импульсов отвечает мышечная ткань. У плода сердечная импульсация начинается на 18-20
- 14. Природа нейрогенного автоматизма: Теория медленной спонтанной диастолической деполяризации
- 15. Механизмы фаз потенциала действия атипичных кардиомиоцитов 1 – фаза спонтанной диастолической деполяризации- медленный вход через мембрану
- 16. Мембранный потенциал атипичных кардиомиоцитов нестабилен - в период диастолы происходит постепенное его уменьшение, приводя к периодическому
- 17. 3. Возбудимость сердца
- 18. Возбудимость — свойство способность возбуждаться (генерировать ПД) в ответ на воздействие раздражителя. Под действием электрических, химических,
- 19. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше, чем в клетках
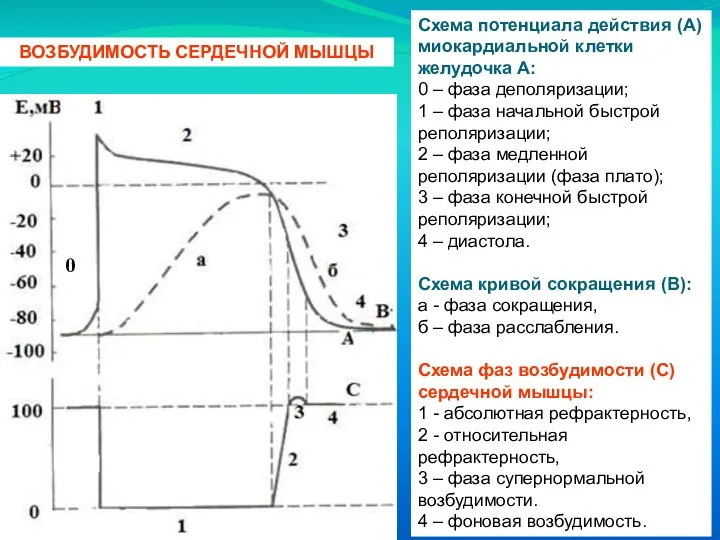
- 20. Механизм формирования фаз ПД типичного кардиомиоцита. 0 - Фаза деполяризации. Под влиянием возбуждения клеток водителей ритма,
- 21. 1 - Фаза быстрой начальной реполяризации. Кратковременна. Связана с выходом из типичного кардиомиоцита ионов К+ и
- 22. 2. Фаза замедленной реполяризации ("плато"). Возникает вследствие наличия в кардиомиоците специфических потенциал-чувствительных Ca-Na –каналов, которые открываются
- 23. 3. Фаза быстрой конечной реполяризации — связана с быстрым выходом из клеток К+. Активно работает электрогенный
- 24. Схема потенциала действия (А) миокардиальной клетки желудочка А: 0 – фаза деполяризации; 1 – фаза начальной
- 25. Длительность потенциала действия кардиомиоцитов составляет 300-400 мс, что соответствует длительности сокращения миокарда, что в 150 раз
- 26. Длительный абсолютный рефрактерный период, т. е. полная невозбудимость сердечной мышцы: предохраняет ее от быстрого повторного возбуждения
- 27. Экстрасистола Желудочковая экстрасистола: Предсердная экстрасистола:
- 28. 4. Сократимость сердца
- 29. Сократимость — способность сердца сокращаться, реализуя тем самым насосную функцию.
- 30. Работа сердечной мышцы подчинена закону «все или ничего» : при достижении раздражающим импульсом пороговой величины возбуждение
- 31. В миокардиальных клетках сокращение всегда закономерно обрывается, и наступает расслабление. Причина: наличие в кардиомиоцитах специальной системы
- 32. 5. Проводимость
- 33. Проводимость – время, в течение которого все сердце как мышечный орган охватывается процессом возбуждения – 15
- 34. Проводимость миокарда и проводящей системы SA-узел - 4,5 – 5 м/с AV-узел - 0,02 – 0,05
- 35. Атриовентрикулярная задержка обеспечивает последовательность сокращений предсердий и желудочков в сердечном цикле. Большая скорость проведения в волокнах
- 36. 6. Гемодинамическая функция сердца
- 37. Последовательность фаз сердечного цикла
- 38. 7. Гемодинамические показатели
- 39. Сердце сокращается в течение жизни человека до 4 млрд. раз, выбрасывая в аорту и способствуя поступлению
- 40. 8. Регуляция сердечной деятельности
- 41. Внутрисердечные механизмы регуляции 1. Внутриклеточная регуляция. 2. Межклеточная регуляция. Внесердечная регуляция Нервная регуляция. Гуморальная регуляция.
- 42. Внутриклеточная регуляция. Гетерометрический механизм Закон Франка-Старлинга: Чем больше растяжение миокарда в диастолу, тем сильнее его сокращение
- 43. Механизмы явления Франка-Старлинга сократительный кардиомиоцит состоит из двух элементов — собственно сократительного и эластического. Сократительный элемент
- 44. Внутриклеточная регуляция. Гомеометрический (метаболический) механизм не связан с изменением длины саркомера. Кардиомиоциты характеризуются цикличностью протекания в
- 45. 2. Межклеточная регуляция Вставочные диски имеют различную структуру и функцию: Одни участки вставочных дисков соединяют миофибриллы,
- 46. Нарушение межклеточных взаимодействий приводит к асинхронному возбуждению клеток миокарда и появлению сердечных аритмий.
- 47. Внесердечная регуляция Нервная регуляция Гуморальная регуляция
- 48. Нервные центры и эффекторы внесердечной нервной регуляции принадлежат автономной нервной системе. За счет нервных влияний осуществляются:
- 49. Парасимпатическая иннервация Ядра блуждающего нерва, иннервирующего сердце, расположены в продолговатом мозге. Блуждающие нервы заканчиваются на интрамуральных
- 50. Отрицательное хронотропное влияние - ацетилхолин взаимодействует с М-холинорецепторами синоатриального узла. В результате открываются калиевые каналы (повышается
- 51. Отрицательное инотропное влияние – ацетилхолин взаимодействует с М-холинорецепторами рабочих кардиомиоцитов. В результате тормозится активность аденилатциклазы и
- 52. Симпатическая иннервация Тела симпатических нейронов, иннервирующих сердце, расположены в боковых рогах 5-ти верхних грудных сегментов спинного
- 53. Положительный хронотропный эффект. Медиатор норадреналин взаимодействует с β- адренорецепторами синоатриального узла. В результате открываются Са2+-каналы -
- 54. Положительный инотропный эффект - норадреналин взаимодействует с β- адренорецепторами рабочих кардиоцитов. Эффекты: а) в клетках активируется
- 55. Блуждающий и симпатический нервы оказывают влияние на работу сердца обычно одновременно (взаимосвязанное влияние) с некоторым преобладанием
- 56. Гипоталамическая регуляция. Гипоталамус имеет прямые связи с ядами блуждающих нервов в продолговатом мозге и боковыми рогами
- 57. Корковая регуляция. Кора головного мозга является органом психической деятельности. Она обеспечивает целостные приспособительные реакции организма. Работа
- 58. 2. Гуморальная регуляция Ацетилхолин Взаимодействуя с М-холинорецепторами наружной мембраны кардиомиоцитов, повышает мембранную проницаемость для ионов калия,
- 59. Адреналин Взаимодействуя с β-адренорецепторами миокардиальных клеток (атипичных и типичных кардиомиоцитов), повышает мембранную проницаемость для ионов кальция,
- 60. Ионы кальция. Увеличение концентрации в плазме крови ионов кальция приводит к повышению возбудимости и сократимости миокарда.
- 61. Ионы калия. Избыток ионов калия во внеклеточной жидкости (около 8 ммоль/л) приводит к слабости сердечной мышцы,
- 62. Типы сосудов
- 64. Артериальное давление
- 65. Капиллярный кровоток
- 66. Венозный возврат
- 67. Присасывающее действие грудной клетки и сердца
- 69. Скачать презентацию

Слайд 31. Автоматия сердца
1. Автоматия сердца

Слайд 4 Автоматия сердца - это способность сердца к ритмическому сокращению без всяких
Автоматия сердца - это способность сердца к ритмическому сокращению без всяких

Слайд 5Сердце человека сокращается:
70 ударов в минуту;
100 тысяч раз в сутки;
40 млн. в год;
2,5 млрд. за всю жизнь.
Сердце человека сокращается:
70 ударов в минуту;
100 тысяч раз в сутки;
40 млн. в год;
2,5 млрд. за всю жизнь.

Слайд 6АВТОМАТИЯ СЕРДЕЧНОЙ МЫШЦЫ:
МИОГЕННАЯ – импульсы появляются в самих мышечных волокнах. Обеспечивает сокращения
АВТОМАТИЯ СЕРДЕЧНОЙ МЫШЦЫ:
МИОГЕННАЯ – импульсы появляются в самих мышечных волокнах. Обеспечивает сокращения

Слайд 7Проводящая система сердца
синусный узел
атриовентрикулярный узел
пучок Гиса
ножки пучка Гиса
волокна Пуркинье
Проводящая система сердца
синусный узел
атриовентрикулярный узел
пучок Гиса
ножки пучка Гиса
волокна Пуркинье
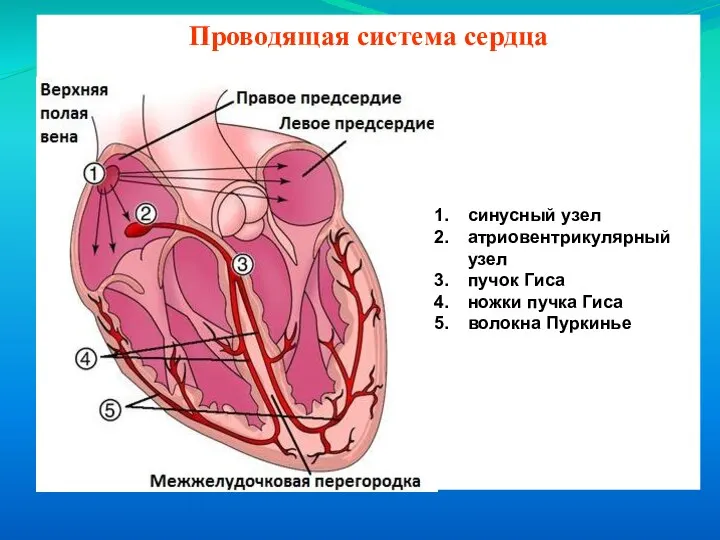
Слайд 8Частота активности элементов проводящей системы:
синусно-предсердный узел (60-80 ударов в минуту). Пейсмекер
Частота активности элементов проводящей системы:
синусно-предсердный узел (60-80 ударов в минуту). Пейсмекер

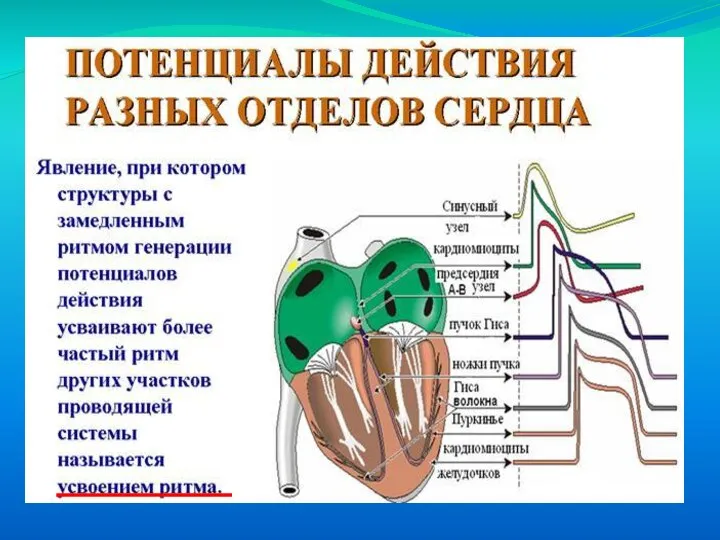
Слайд 10Закон градиента автоматизма сердца– у всех позвоночных степень автоматии тем выше, чем
Закон градиента автоматизма сердца– у всех позвоночных степень автоматии тем выше, чем

Слайд 112. Природа автоматизма
2. Природа автоматизма

Слайд 12Особенности клеток атипичной мышечной ткани:
Только несколько клеток синусно-предсердного узла, называемых истинными пейсмекерами,
Только несколько клеток синусно-предсердного узла, называемых истинными пейсмекерами,

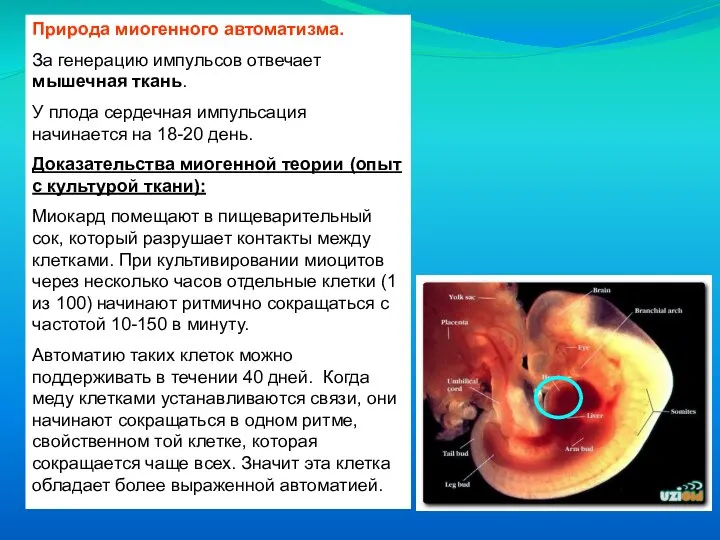
Слайд 13Природа миогенного автоматизма.
За генерацию импульсов отвечает мышечная ткань.
У плода сердечная
Природа миогенного автоматизма.
За генерацию импульсов отвечает мышечная ткань.
У плода сердечная
Слайд 14Природа нейрогенного автоматизма: Теория медленной спонтанной диастолической деполяризации
Природа нейрогенного автоматизма: Теория медленной спонтанной диастолической деполяризации
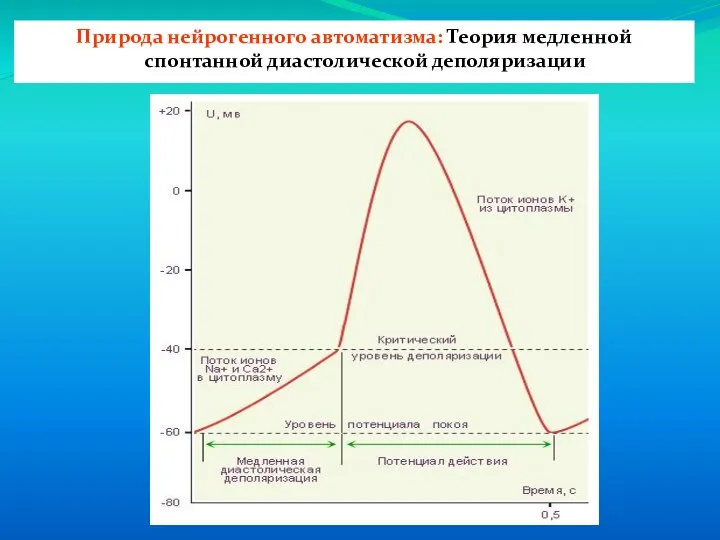
Слайд 15Механизмы фаз потенциала действия атипичных кардиомиоцитов
1 – фаза спонтанной диастолической деполяризации- медленный
1 – фаза спонтанной диастолической деполяризации- медленный
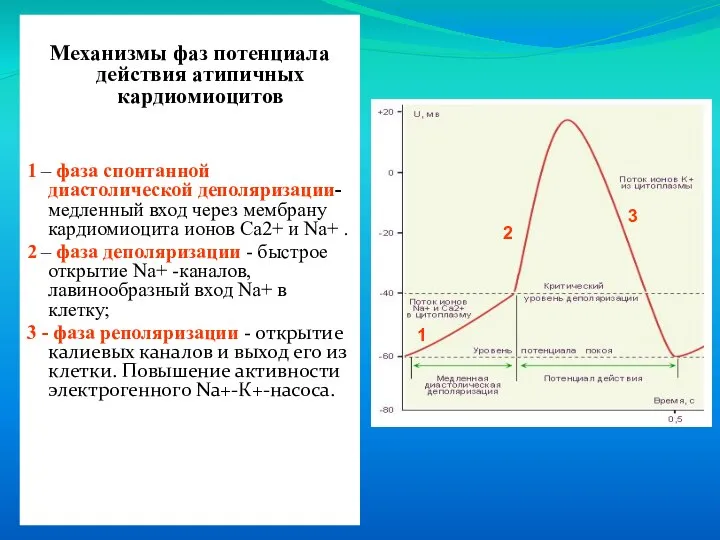
Слайд 16Мембранный потенциал атипичных кардиомиоцитов нестабилен - в период диастолы происходит постепенное его

Слайд 173. Возбудимость сердца
3. Возбудимость сердца

Слайд 18Возбудимость — свойство способность возбуждаться (генерировать ПД) в ответ на воздействие раздражителя.
Под
Возбудимость — свойство способность возбуждаться (генерировать ПД) в ответ на воздействие раздражителя.
Под

Слайд 19В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина
В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина

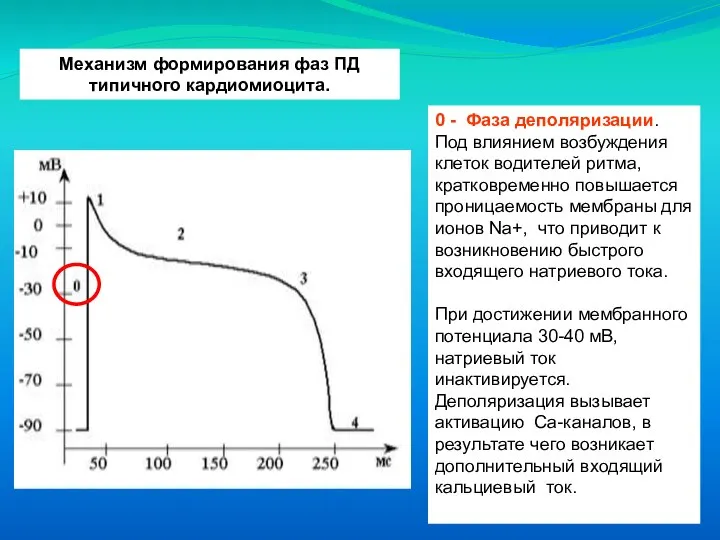
Слайд 20Механизм формирования фаз ПД типичного кардиомиоцита.
0 - Фаза деполяризации. Под влиянием возбуждения
Механизм формирования фаз ПД типичного кардиомиоцита.
0 - Фаза деполяризации. Под влиянием возбуждения
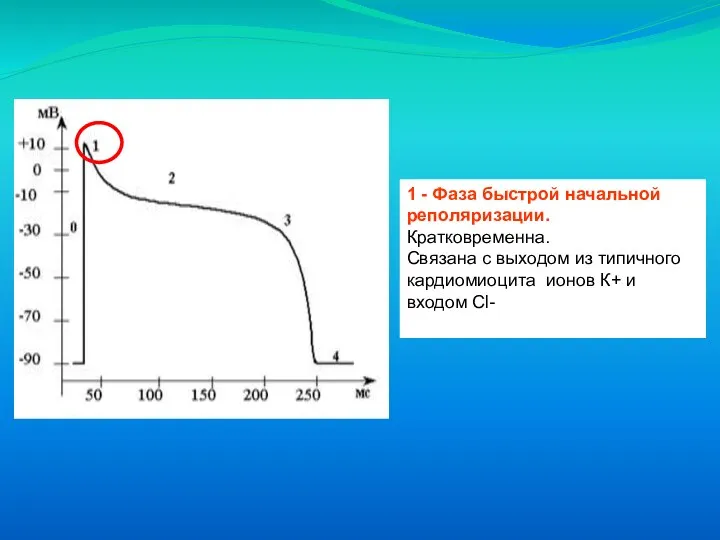
Слайд 211 - Фаза быстрой начальной реполяризации. Кратковременна.
Связана с выходом из типичного
1 - Фаза быстрой начальной реполяризации. Кратковременна.
Связана с выходом из типичного
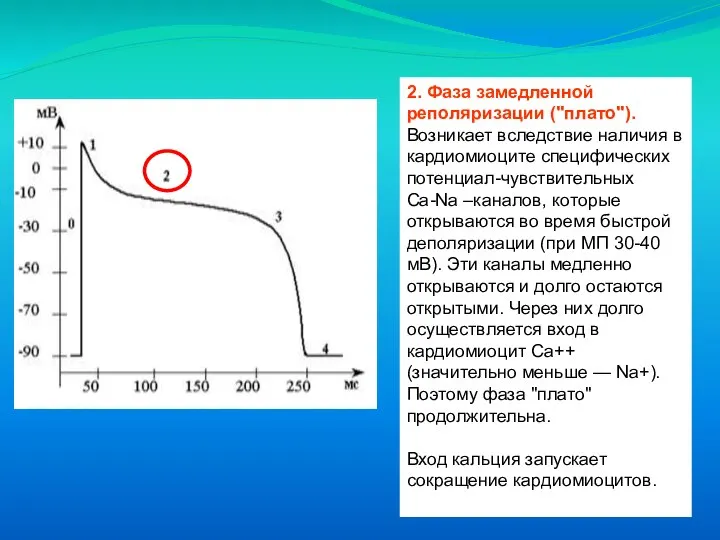
Слайд 222. Фаза замедленной реполяризации ("плато"). Возникает вследствие наличия в кардиомиоците специфических потенциал-чувствительных
2. Фаза замедленной реполяризации ("плато"). Возникает вследствие наличия в кардиомиоците специфических потенциал-чувствительных
Слайд 233. Фаза быстрой конечной реполяризации — связана с быстрым выходом из клеток
3. Фаза быстрой конечной реполяризации — связана с быстрым выходом из клеток
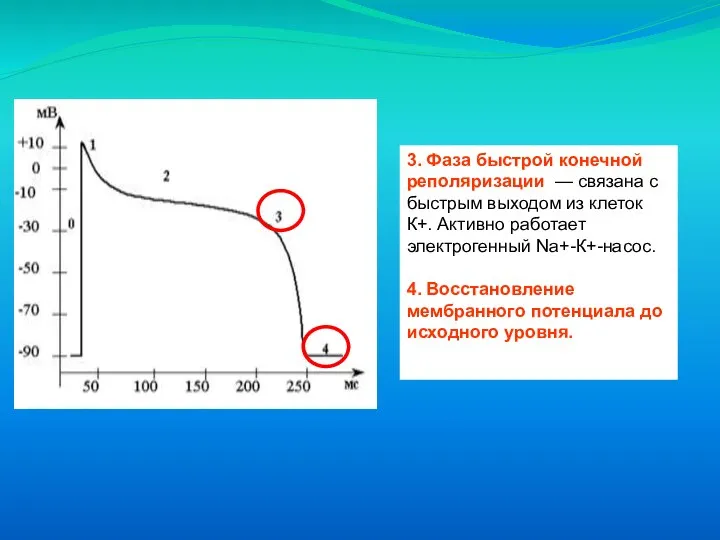
Слайд 24Схема потенциала действия (А) миокардиальной клетки желудочка А:
0 – фаза деполяризации;
Схема потенциала действия (А) миокардиальной клетки желудочка А:
0 – фаза деполяризации;
Слайд 25Длительность потенциала действия кардиомиоцитов составляет 300-400 мс, что соответствует длительности сокращения миокарда, что
Длительность потенциала действия кардиомиоцитов составляет 300-400 мс, что соответствует длительности сокращения миокарда, что

Слайд 26Длительный абсолютный рефрактерный период, т. е. полная невозбудимость сердечной мышцы:
предохраняет ее от
Длительный абсолютный рефрактерный период, т. е. полная невозбудимость сердечной мышцы:
предохраняет ее от

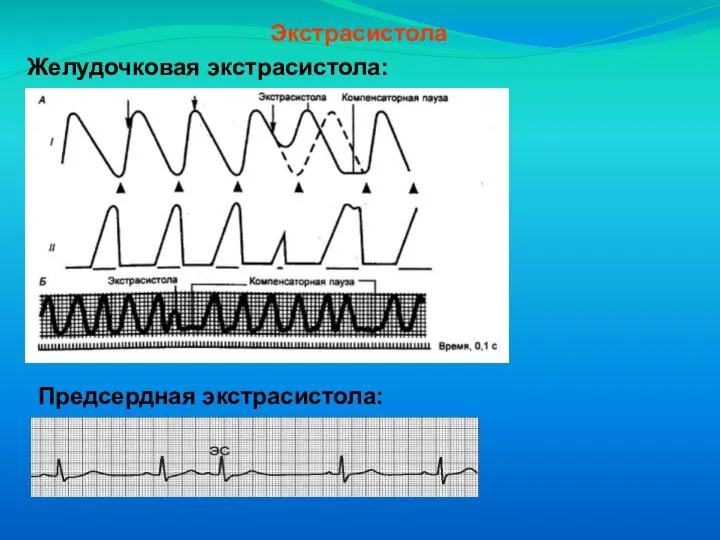
Слайд 27Экстрасистола
Желудочковая экстрасистола:
Предсердная экстрасистола:
Экстрасистола
Желудочковая экстрасистола:
Предсердная экстрасистола:
Слайд 284. Сократимость сердца
4. Сократимость сердца

Слайд 29Сократимость — способность сердца сокращаться, реализуя тем самым насосную функцию.
Сократимость — способность сердца сокращаться, реализуя тем самым насосную функцию.

Слайд 30Работа сердечной мышцы подчинена закону «все или ничего» : при достижении раздражающим
Работа сердечной мышцы подчинена закону «все или ничего» : при достижении раздражающим

Слайд 31В миокардиальных клетках сокращение всегда закономерно обрывается, и наступает расслабление.
Причина: наличие
В миокардиальных клетках сокращение всегда закономерно обрывается, и наступает расслабление.
Причина: наличие

Слайд 325. Проводимость
5. Проводимость

Слайд 33Проводимость – время, в течение которого все сердце как мышечный орган охватывается
Проводимость – время, в течение которого все сердце как мышечный орган охватывается

Слайд 34Проводимость миокарда и проводящей системы
SA-узел - 4,5 – 5 м/с
AV-узел -
Проводимость миокарда и проводящей системы
SA-узел - 4,5 – 5 м/с
AV-узел -

Слайд 35Атриовентрикулярная задержка обеспечивает последовательность сокращений предсердий и желудочков в сердечном цикле.
Большая скорость
Атриовентрикулярная задержка обеспечивает последовательность сокращений предсердий и желудочков в сердечном цикле.
Большая скорость

Слайд 366. Гемодинамическая функция сердца
6. Гемодинамическая функция сердца

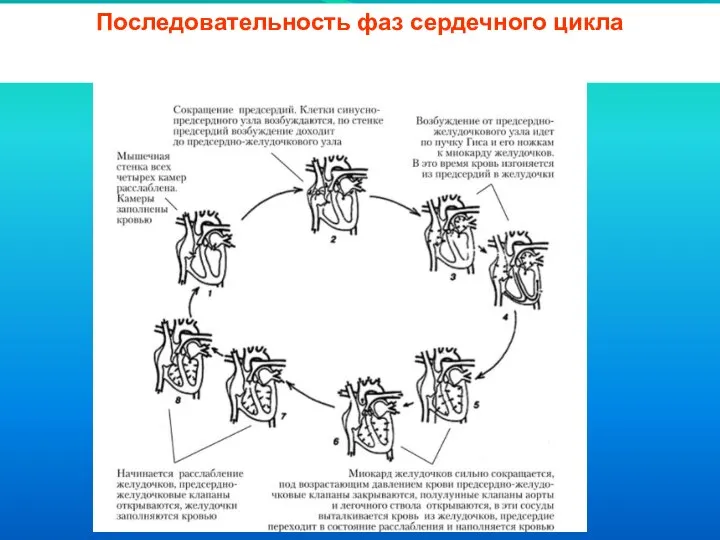
Слайд 37Последовательность фаз сердечного цикла
Последовательность фаз сердечного цикла
Слайд 387. Гемодинамические показатели
7. Гемодинамические показатели

Слайд 39Сердце сокращается в течение жизни человека до 4 млрд. раз, выбрасывая в
Сердце сокращается в течение жизни человека до 4 млрд. раз, выбрасывая в

Слайд 408. Регуляция сердечной деятельности
8. Регуляция сердечной деятельности

Слайд 41Внутрисердечные механизмы регуляции
1. Внутриклеточная регуляция.
2. Межклеточная регуляция.
Внесердечная регуляция
Нервная регуляция.
Гуморальная
Внутрисердечные механизмы регуляции
1. Внутриклеточная регуляция.
2. Межклеточная регуляция.
Внесердечная регуляция
Нервная регуляция.
Гуморальная

Слайд 42Внутриклеточная регуляция.
Гетерометрический механизм
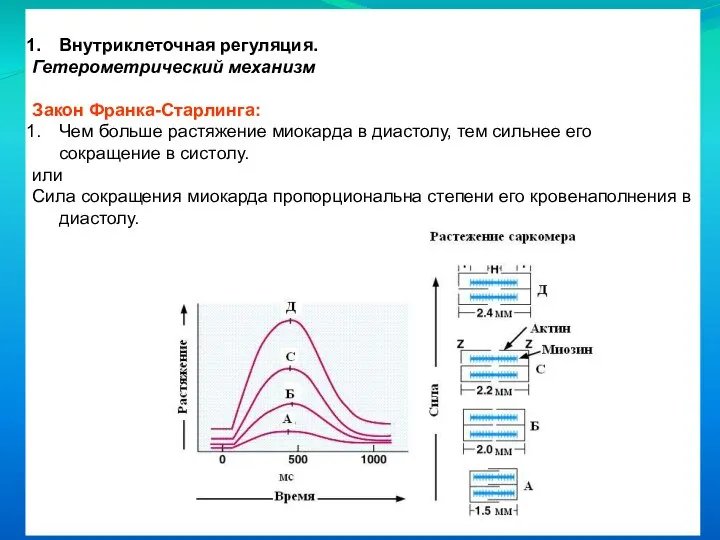
Закон Франка-Старлинга:
Чем больше растяжение миокарда в диастолу, тем
Гетерометрический механизм
Закон Франка-Старлинга:
Чем больше растяжение миокарда в диастолу, тем
Слайд 43Механизмы явления Франка-Старлинга
сократительный кардиомиоцит состоит из двух элементов — собственно сократительного
сократительный кардиомиоцит состоит из двух элементов — собственно сократительного

Слайд 44Внутриклеточная регуляция.
Гомеометрический (метаболический) механизм не связан с изменением длины саркомера.
Кардиомиоциты
Гомеометрический (метаболический) механизм не связан с изменением длины саркомера.
Кардиомиоциты

Слайд 452. Межклеточная регуляция
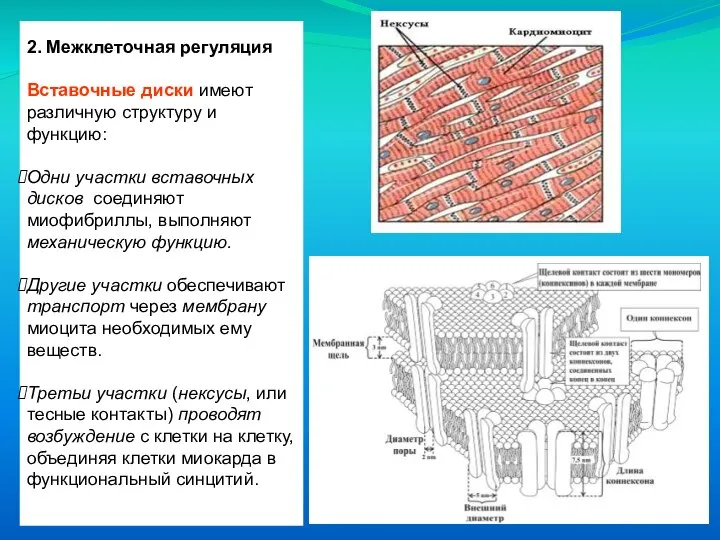
Вставочные диски имеют различную структуру и функцию:
Одни участки вставочных
2. Межклеточная регуляция
Вставочные диски имеют различную структуру и функцию:
Одни участки вставочных
Слайд 46Нарушение межклеточных взаимодействий приводит к асинхронному возбуждению клеток миокарда и появлению сердечных
Нарушение межклеточных взаимодействий приводит к асинхронному возбуждению клеток миокарда и появлению сердечных

Слайд 47Внесердечная регуляция
Нервная регуляция
Гуморальная регуляция
Внесердечная регуляция
Нервная регуляция
Гуморальная регуляция

Слайд 48Нервные центры и эффекторы внесердечной нервной регуляции принадлежат автономной нервной системе.
За
Нервные центры и эффекторы внесердечной нервной регуляции принадлежат автономной нервной системе.
За

Слайд 49Парасимпатическая иннервация
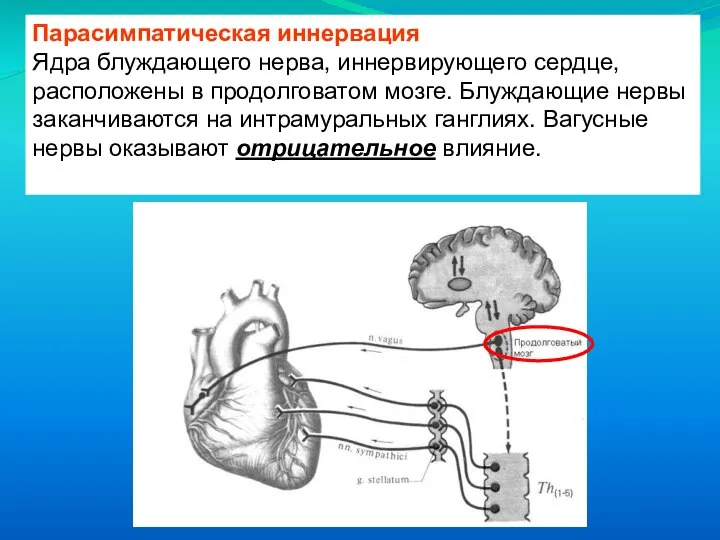
Ядра блуждающего нерва, иннервирующего сердце, расположены в продолговатом мозге. Блуждающие
Парасимпатическая иннервация
Ядра блуждающего нерва, иннервирующего сердце, расположены в продолговатом мозге. Блуждающие
Слайд 50Отрицательное хронотропное влияние - ацетилхолин взаимодействует с М-холинорецепторами синоатриального узла. В результате
Отрицательное хронотропное влияние - ацетилхолин взаимодействует с М-холинорецепторами синоатриального узла. В результате

Слайд 51Отрицательное инотропное влияние –
ацетилхолин взаимодействует с М-холинорецепторами рабочих кардиомиоцитов. В результате
Отрицательное инотропное влияние –
ацетилхолин взаимодействует с М-холинорецепторами рабочих кардиомиоцитов. В результате

Слайд 52Симпатическая иннервация
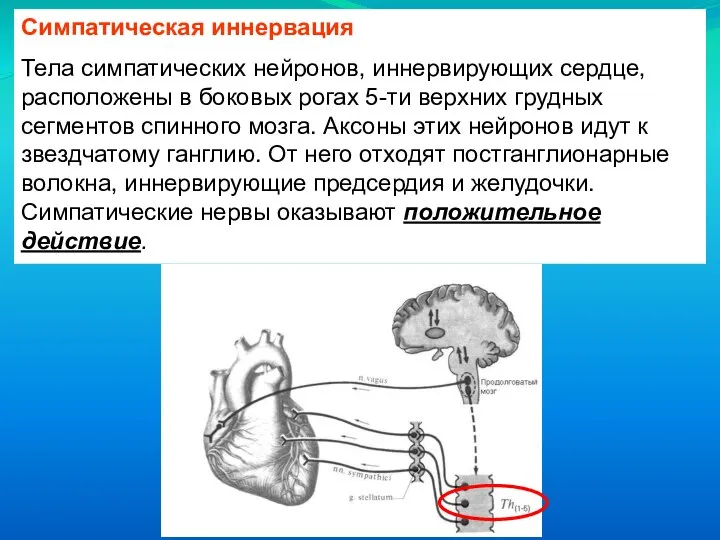
Тела симпатических нейронов, иннервирующих сердце, расположены в боковых рогах 5-ти
Симпатическая иннервация
Тела симпатических нейронов, иннервирующих сердце, расположены в боковых рогах 5-ти
Слайд 53Положительный хронотропный эффект. Медиатор норадреналин взаимодействует с β- адренорецепторами синоатриального узла. В
Положительный хронотропный эффект. Медиатор норадреналин взаимодействует с β- адренорецепторами синоатриального узла. В

Слайд 54Положительный инотропный эффект - норадреналин взаимодействует с β- адренорецепторами рабочих кардиоцитов.
Эффекты:
а)
Положительный инотропный эффект - норадреналин взаимодействует с β- адренорецепторами рабочих кардиоцитов.
Эффекты:
а)

Слайд 55Блуждающий и симпатический нервы оказывают влияние на работу сердца обычно одновременно (взаимосвязанное
Блуждающий и симпатический нервы оказывают влияние на работу сердца обычно одновременно (взаимосвязанное

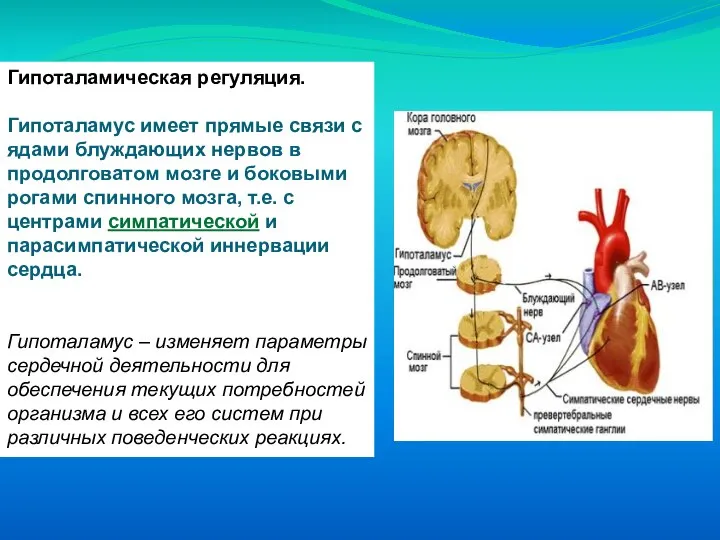
Слайд 56Гипоталамическая регуляция.
Гипоталамус имеет прямые связи с ядами блуждающих нервов в продолговатом мозге
Гипоталамическая регуляция.
Гипоталамус имеет прямые связи с ядами блуждающих нервов в продолговатом мозге
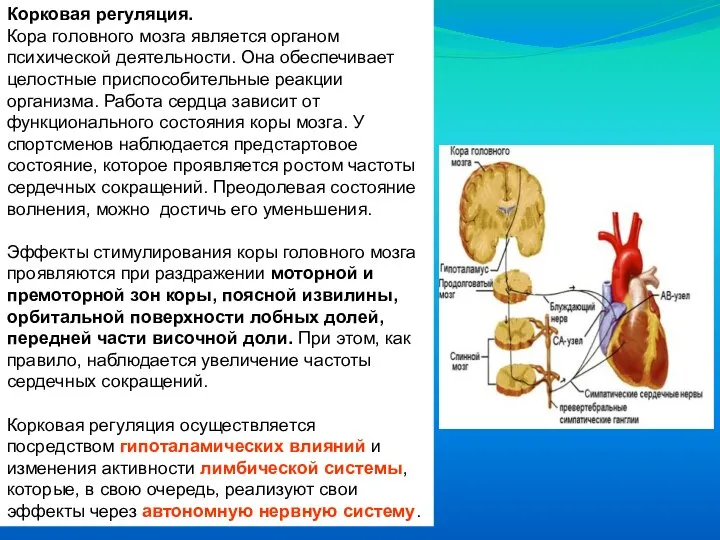
Слайд 57Корковая регуляция.
Кора головного мозга является органом психической деятельности. Она обеспечивает целостные приспособительные
Корковая регуляция.
Кора головного мозга является органом психической деятельности. Она обеспечивает целостные приспособительные
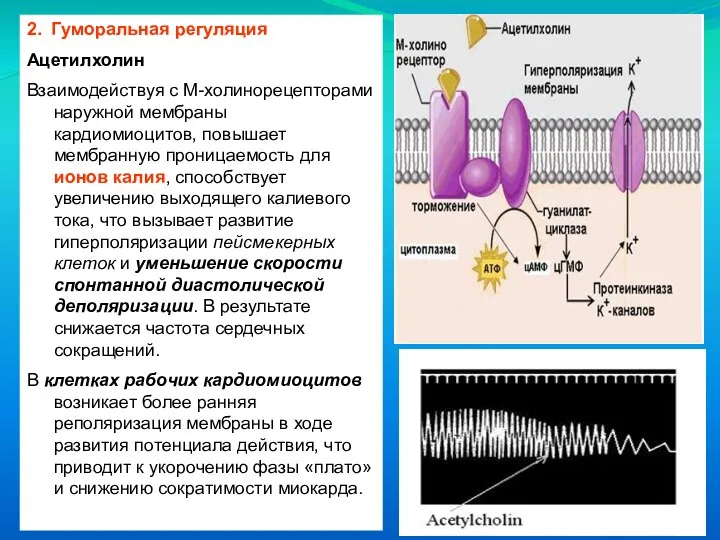
Слайд 582. Гуморальная регуляция
Ацетилхолин
Взаимодействуя с М-холинорецепторами наружной мембраны кардиомиоцитов, повышает мембранную проницаемость для
2. Гуморальная регуляция
Ацетилхолин
Взаимодействуя с М-холинорецепторами наружной мембраны кардиомиоцитов, повышает мембранную проницаемость для
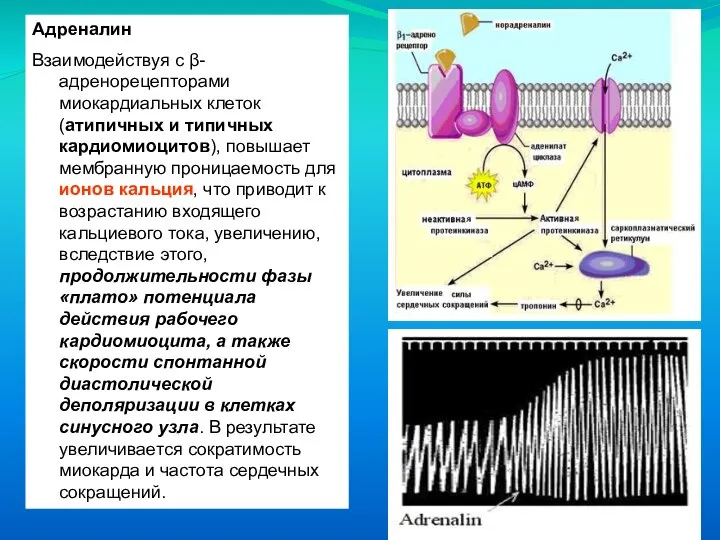
Слайд 59Адреналин
Взаимодействуя с β-адренорецепторами миокардиальных клеток (атипичных и типичных кардиомиоцитов), повышает мембранную проницаемость
Адреналин
Взаимодействуя с β-адренорецепторами миокардиальных клеток (атипичных и типичных кардиомиоцитов), повышает мембранную проницаемость
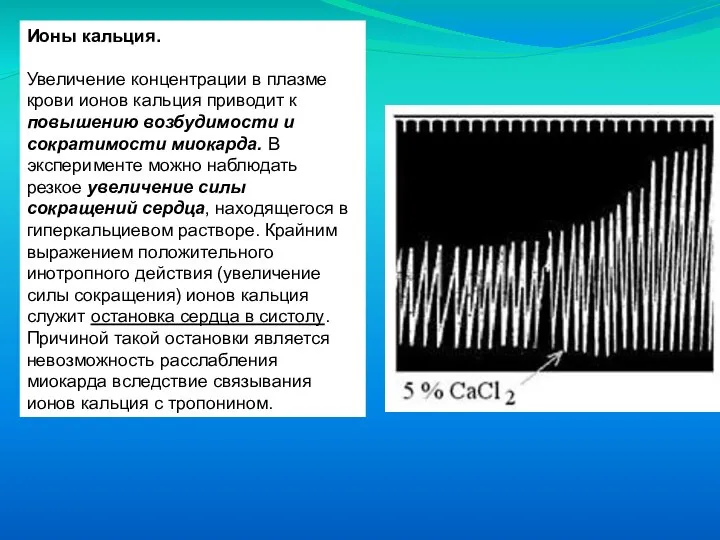
Слайд 60Ионы кальция.
Увеличение концентрации в плазме крови ионов кальция приводит к повышению
Ионы кальция.
Увеличение концентрации в плазме крови ионов кальция приводит к повышению
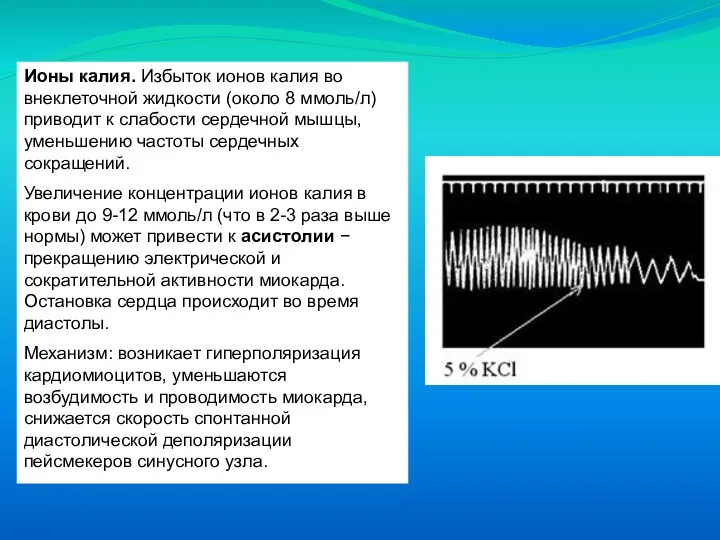
Слайд 61Ионы калия. Избыток ионов калия во внеклеточной жидкости (около 8 ммоль/л) приводит
Ионы калия. Избыток ионов калия во внеклеточной жидкости (около 8 ммоль/л) приводит
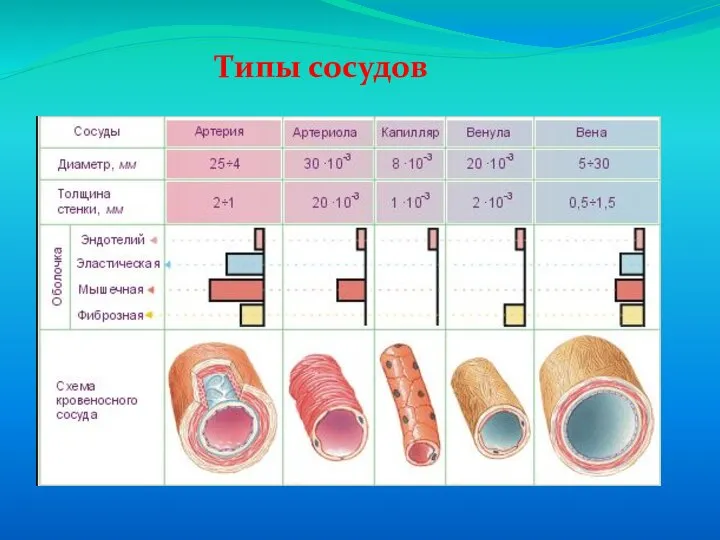
Слайд 62Типы сосудов
Типы сосудов
Слайд 64Артериальное давление
Артериальное давление
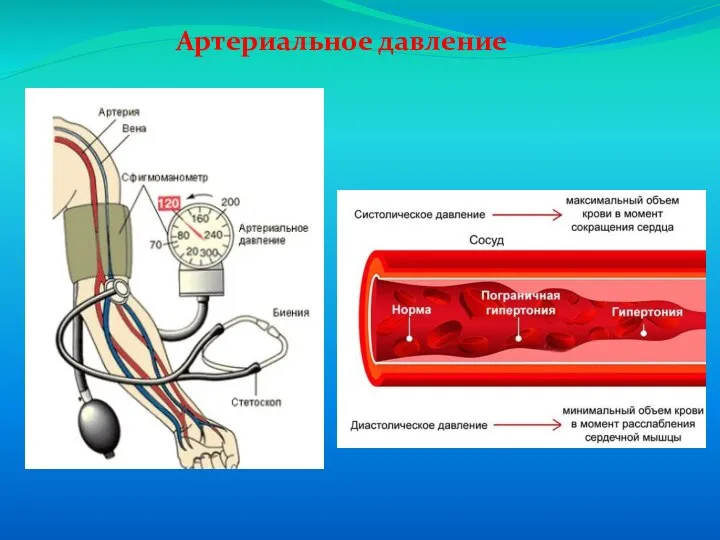
Слайд 65Капиллярный кровоток
Капиллярный кровоток
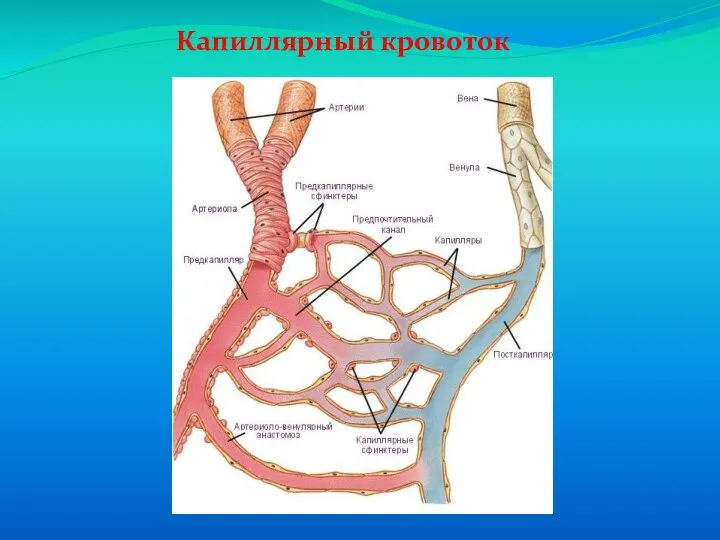
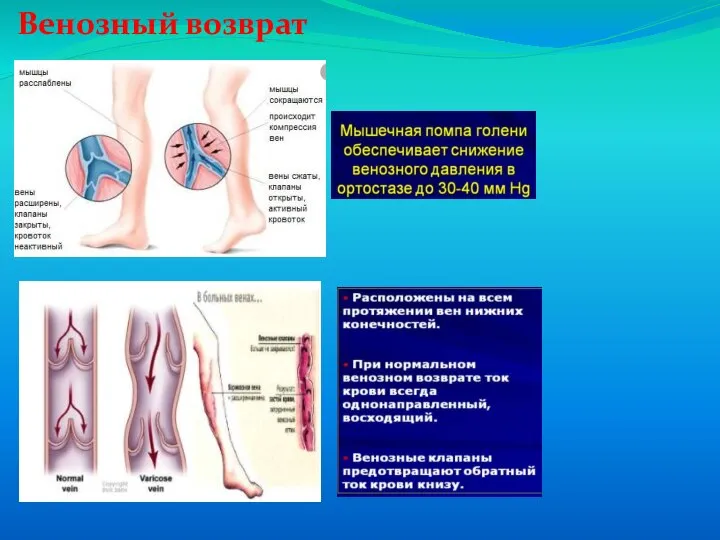
Слайд 66Венозный возврат
Венозный возврат
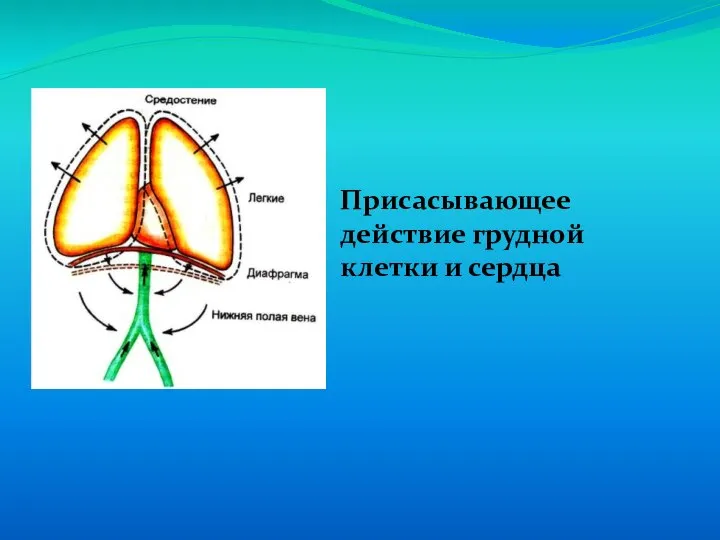
Слайд 67Присасывающее действие грудной клетки и сердца
Присасывающее действие грудной клетки и сердца
 Викторина «МЛЕКОПИТАЮЩИЕ»
Викторина «МЛЕКОПИТАЮЩИЕ» Выращивание фасоли. Растение на окошке
Выращивание фасоли. Растение на окошке Родина культурных растений
Родина культурных растений Лабораторная работа № 10. Сравнительный анализ строения скелетов черепахи, ящерицы и змеи (связанные с ее образом жизни)
Лабораторная работа № 10. Сравнительный анализ строения скелетов черепахи, ящерицы и змеи (связанные с ее образом жизни) Презентация на тему Сердце и кровообращение
Презентация на тему Сердце и кровообращение  Лексическая тема Насекомые
Лексическая тема Насекомые Аэробное и анаэробное дыхание микроорганизмов
Аэробное и анаэробное дыхание микроорганизмов Зрение. Открытый классный час
Зрение. Открытый классный час Основы психогенетики. Тема 1
Основы психогенетики. Тема 1 Презентация на тему Понятие о систематике растений
Презентация на тему Понятие о систематике растений  Корень. Строение и значение
Корень. Строение и значение Цианокобаламин (витамин в12)
Цианокобаламин (витамин в12) Строение и значение кожи
Строение и значение кожи Дизонтогенез
Дизонтогенез Скелет поясов и свободных конечностей
Скелет поясов и свободных конечностей Лес. Виды леса
Лес. Виды леса Виды птиц
Виды птиц Презентация на тему Условия жизни и многообразие диких животных
Презентация на тему Условия жизни и многообразие диких животных  Они рядом с нами – редкие и исчезающие животные
Они рядом с нами – редкие и исчезающие животные My little experiment
My little experiment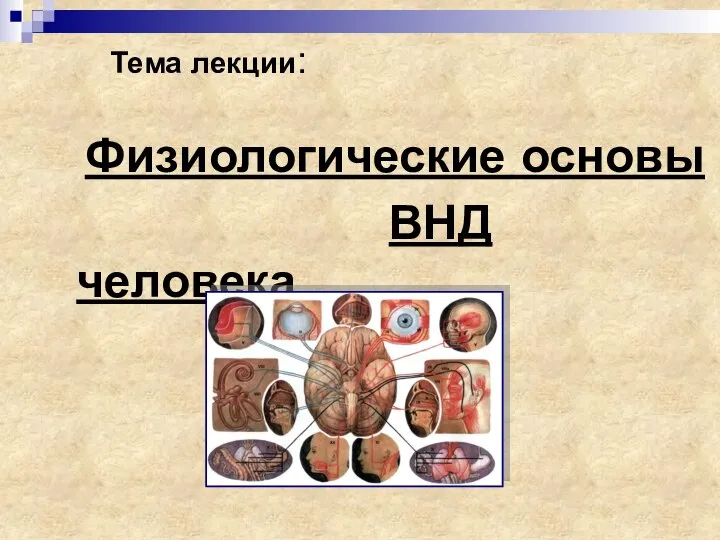 Физиологические основы ВНД человека
Физиологические основы ВНД человека Ферменты
Ферменты Беспозвоночные водоема
Беспозвоночные водоема Презентация на тему Этапы эволюции человека
Презентация на тему Этапы эволюции человека  Презентация на тему Животные луга
Презентация на тему Животные луга  Презентация на тему Основные свойства живых организмов (6 класс)
Презентация на тему Основные свойства живых организмов (6 класс)  Родительский лекторий. Качество питания и здоровье
Родительский лекторий. Качество питания и здоровье Оптическая система глаза
Оптическая система глаза