- Клетки оргнизма

Содержание
- 4. Прокариоты имеют неоформленное ядро (нуклеоид).На ультратонких срезах зона нуклеоида представлена тонкими рыхлыми фибрилами толщиной 2-7нм Зона
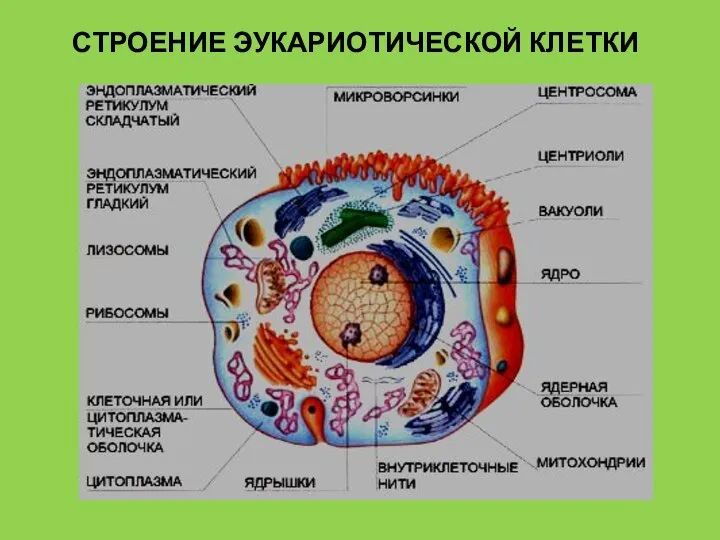
- 5. СТРОЕНИЕ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
- 6. ПОВЕРХНОСТНЫЙ АППАРАТ КЛЕТОК Общая характеристика поверхностного аппарата Любая разновидность прокаиотических и эукариотических клеток состоит из трех
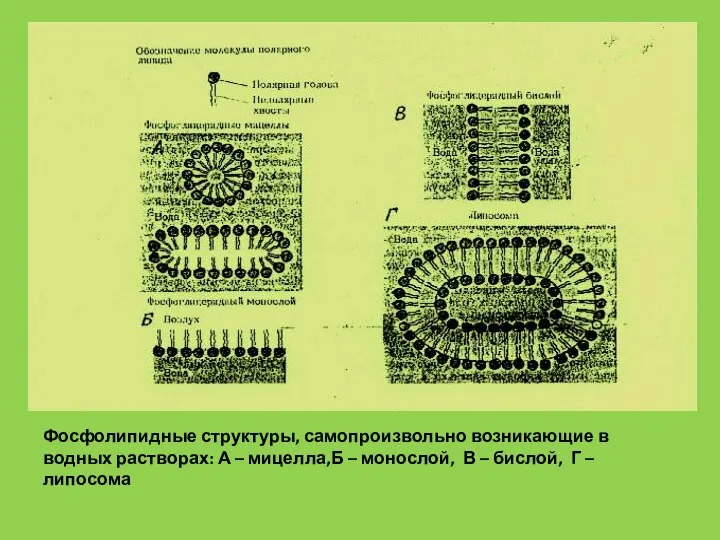
- 7. Фосфолипидные структуры, самопроизвольно возникающие в водных растворах: А – мицелла,Б – монослой, В – бислой, Г
- 9. поверхностное натяжение таких пленок намного выше , чем у биологических мембран. При добавлении к липидным пленкам
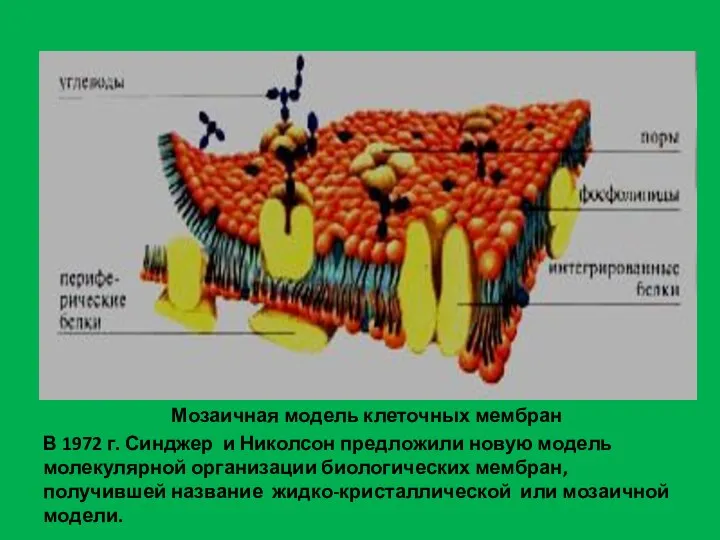
- 10. Мозаичная модель клеточных мембран В 1972 г. Синджер и Николсон предложили новую модель молекулярной организации биологических
- 12. Липиды – органические соединения с различной структурой , но общими свойствами. Они нерастворимы в воде, но
- 13. Липиды мембран - липидный состав клеточных мембран весьма разнообразен. Характерными представителями липидов клеточных мембран являются фосфолипиды
- 14. Жирные кислоты
- 15. * В классификации жирных кислот вслед за числом углеродных атомов через двоеточие указывается количество двойных связей,
- 16. Липидный состав некоторых биологических мембран
- 17. В состав биологических мембран входят липиды трёх основных классов: фосфолипиды, гликолипиды, стероиды Фосфолипиды – наиболее часто
- 18. Гликолипиды мембран представлены цереброзидами, сулфатидами и ганглиозидами. Все они содержат углеводные компоненты. Цереброзиды – углеводные производные
- 19. Молекула холестерина состоит из компактного, жёсткого гидрофобного ядра, а полярной головкой является гидроксильная группа. Молекула холестерина
- 20. Липидные плотики (рафты). Холестерин участвует в формировании рафтов — дискретных мембранных доменов, богатых сфинголипидами и холестерином.
- 21. В липидном бислое выделяют два пула липидов: аннулярные, или пограничные липиды взаимодействуют с мембранными белками и
- 22. Функции липидов мембран состоят в том, что они: • формируют липидный бислой - структурную основу мембран;
- 23. Липидный бислой мембран находится в динамичном состоянии. Известно 4 вида интрамолекулярных движений липидов: латеральная диффузия, скорость
- 24. при постоянной длине ацильной цепи и температуре увеличение количества двойных связей повышают текучесть мембраны; при постоянном
- 25. а - изменение структуры и свойств мембран при изменении температуры окружающей среды и осмотического давления. А1
- 26. Р ≥ 10 мкм/с Р ≤ 1 мкм/с Р Фазовое состояние липидного бислоя (текучесть зависит от
- 27. Мембранные белки Мембраны содержат от 20 до 82 % белка. Белки ответственны за функциональную активность мембран.
- 28. Периферические белки связаны с поверхностью мембраны за счёт слабых электростатических взаимодействий с полярными головками липидных молекул
- 29. Интегральные, или внутренние, белки погружены в липидный бислой или пронизывают его, фиксируясь в мембране с помощью
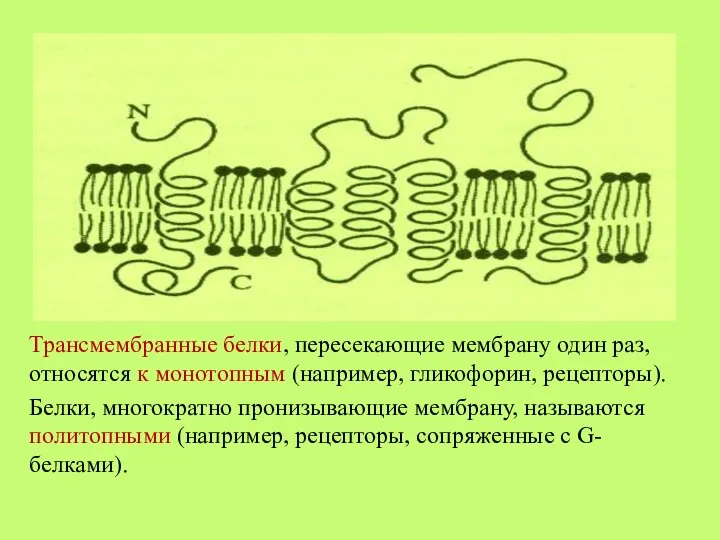
- 30. Трансмембранные белки, пересекающие мембрану один раз, относятся к монотопным (например, гликофорин, рецепторы). Белки, многократно пронизывающие мембрану,
- 31. Ассиметричное расположение молекул белков в мембранах, определякмое расположени полипептидных цепейем С и N – концевых и
- 32. Углеводы мембраны Данная группа веществ составляетпримерно около 10% от общего состава соединений плазмалеммы. В простом виде
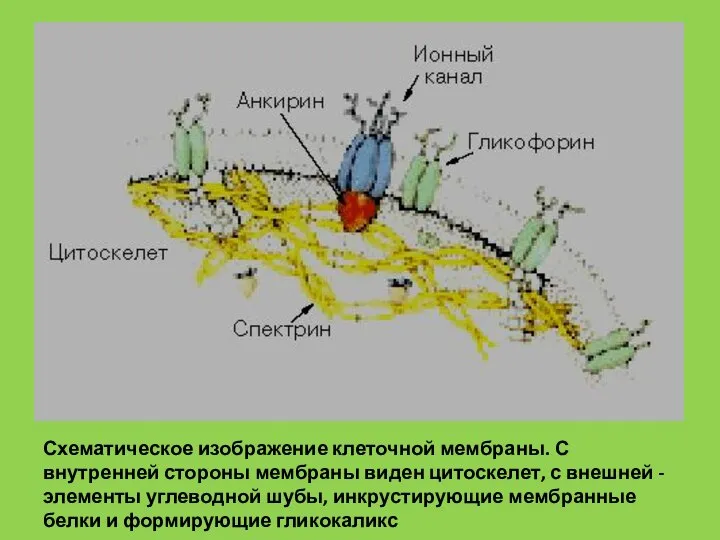
- 33. Схематическое изображение клеточной мембраны. С внутренней стороны мембраны виден цитоскелет, с внешней - элементы углеводной шубы,
- 35. Наружная часть содержит преимущественно фосфатидилхолин(75%) , сфингомиелин (80%) , и фосфотидилэтаноламн(20%) . Внутренняя часть плазмолеммы, обращённая
- 36. 80%сфингомиелина,75%фосфотидилхолина и 20%фосфотидилэтаноламна локализованы на наружной поверхности плазматической мембраны На внутренней – располагается весь фосфатидилсерин и
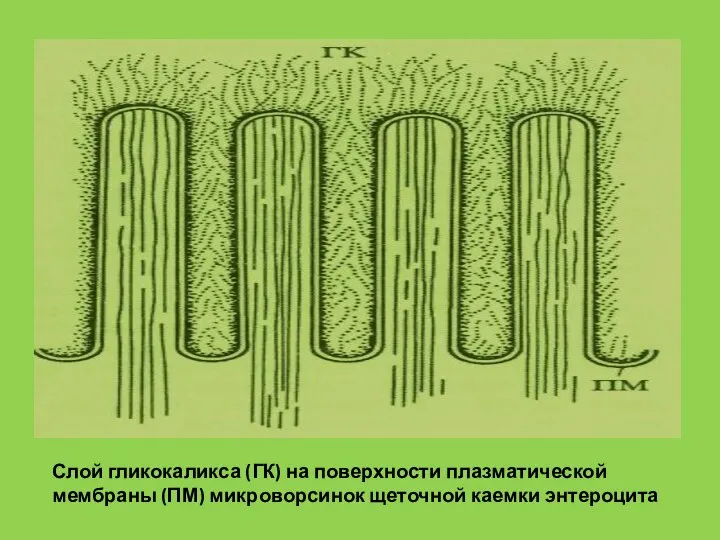
- 37. Слой гликокаликса (ГК) на поверхности плазматической мембраны (ПМ) микроворсинок щеточной каемки энтероцита
- 38. Гликокаликс имеет вид рыхлого волокнистого слоя толщиной 3-4 нм, покрывающего всю поверхность клетки. Гликокаликс обнаружен практически
- 39. Основным компонентом клеточных стенок, значение которого часто недооценивается, является вода. Матрикс клеточной стенки на 75% состоит
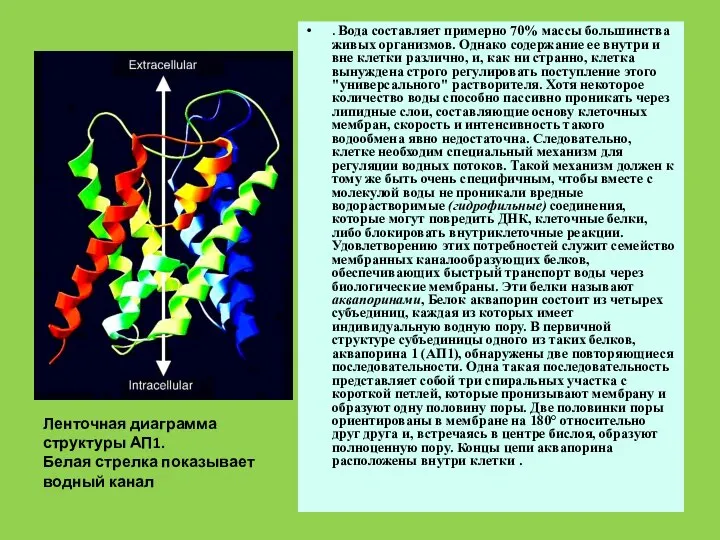
- 40. Вода составляет примерно 70% массы большинства живыхорганизмов. Однако содержание ее внутри и вне клетки различно, и,
- 41. Схема третичной структуры аквапорина:модель «песочные часы» структура тетрамера (Из: Ren et al., 200 (из Kjellbom et.
- 42. Аквапорины в высшей степени селективны для воды, они не пропускают даже ион гидроксония (H3O+). В то
- 43. Молекулы целлюлозы (несколько десятков на поперечном срезе)собраны за счет водородных связей в микрофибриллы. Микрофибриллы – «голые
- 44. Гликозилтрансферазы, участвующие в формировании полисахаридных молекул, являются мембраносвязанными ферментами. По характеру взаимодействия с мембраной гликозилтрансферазы подразделяются
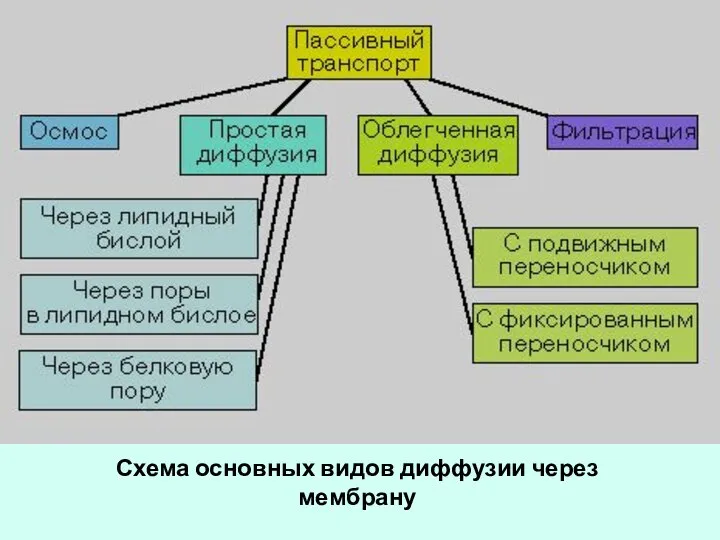
- 45. Схема основных видов диффузии через мембрану
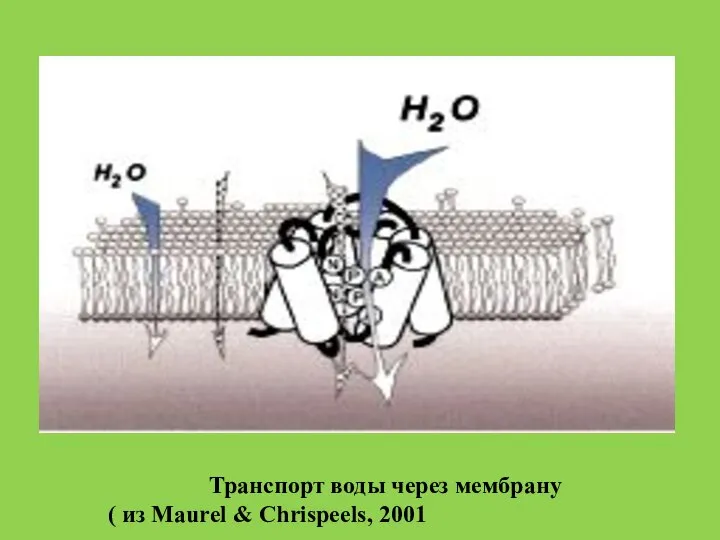
- 46. Транспорт воды через мембрану ( из Maurel & Chrispeels, 2001
- 47. . Вода составляет примерно 70% массы большинства живых организмов. Однако содержание ее внутри и вне клетки
- 48. Схема третичной структуры аквапорина:модель «песочные часы» (из Kjellbom et. al., структура тетрамера (Из: Ren et al.,
- 49. Аквапорины в высшей степени селективны для воды, они не пропускают даже ион гидроксония (H3O+). В то
- 51. Пассивный транспорт характеризуется низкой специфичностью. Молекулы в обоих направлениях перемещаются по градиенту концентрации без затрат энергии.
- 54. Окаймленные пузырьки а–вид со стороны цитозоля Б – трискелионы на поверхности пузырька Последовательные стадии образования и
- 56. с. Окаймленные ямки и вакуоли Последовательные стадии связывания лигандов и образование вакуолей: 1— плазматическая мембрана, 2
- 57. . Образование отщепляющейся вакуоли 1- мембрана; 2- рецепторный белок; 3- белок, связывающий ГТФ; 4- белки, покрывающие
- 58. . Окаймленные пузырьки а - вид со стороны цитозоля; б - трискелеоны на поверхности пузырька
- 59. Поступление в клетку ЛНП(липопротеид низкой плотности) с помощью опосредуемого рецепторами эндоцитоза 1 - плазмалемма; 2 частица
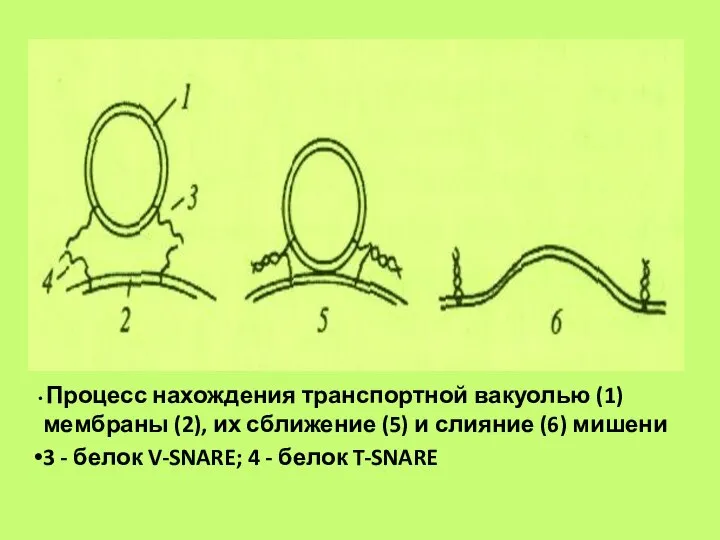
- 60. Процесс нахождения транспортной вакуолью (1) мембраны (2), их сближение (5) и слияние (6) мишени 3 -
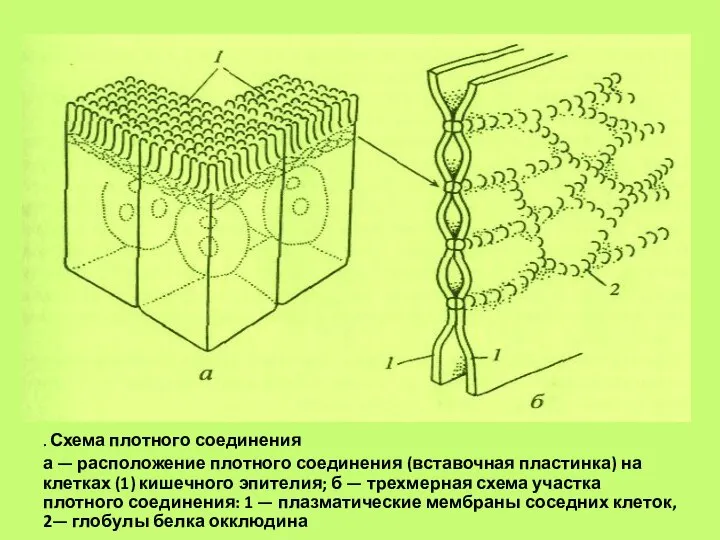
- 61. . Схема плотного соединения а — расположение плотного соединения (вставочная пластинка) на клетках (1) кишечного эпителия;
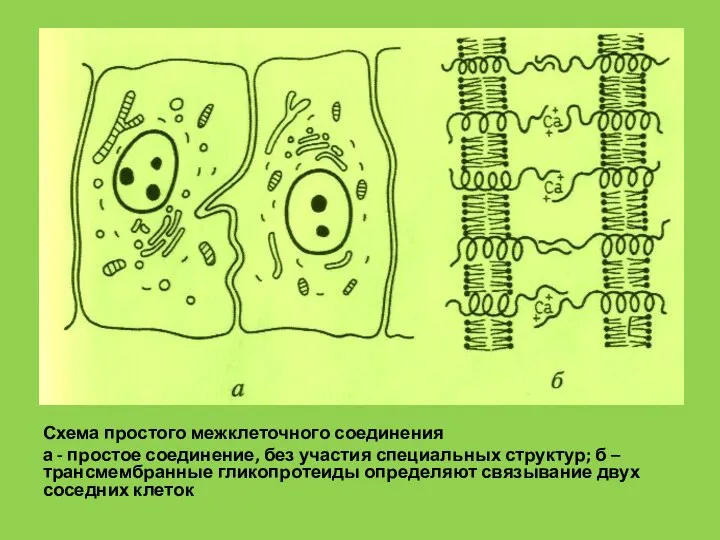
- 62. Схема простого межклеточного соединения а - простое соединение, без участия специальных структур; б – трансмембранные гликопротеиды
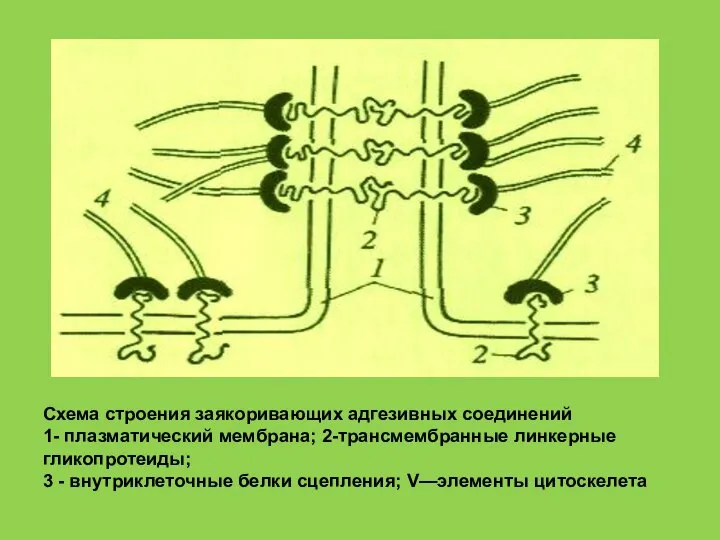
- 63. Схема строения заякоривающих адгезивных соединений 1- плазматический мембрана; 2-трансмембранные линкерные гликопротеиды; 3 - внутриклеточные белки сцепления;
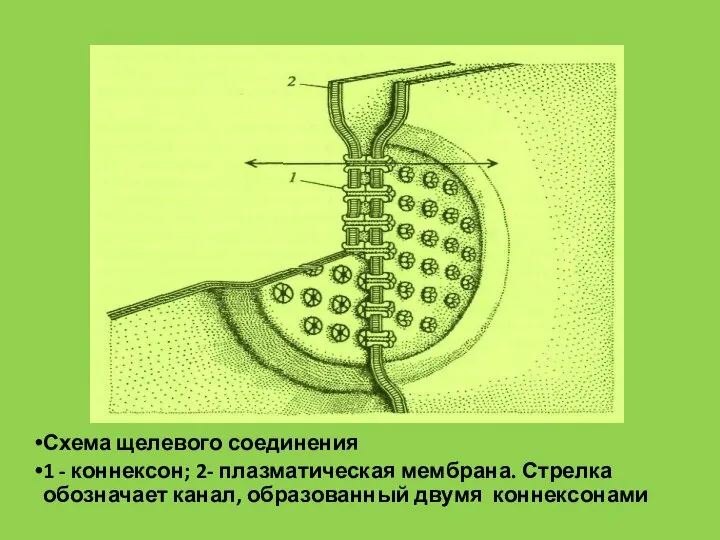
- 64. Схема щелевого соединения 1 - коннексон; 2- плазматическая мембрана. Стрелка обозначает канал, образованный двумя коннексонами
- 66. Скачать презентацию
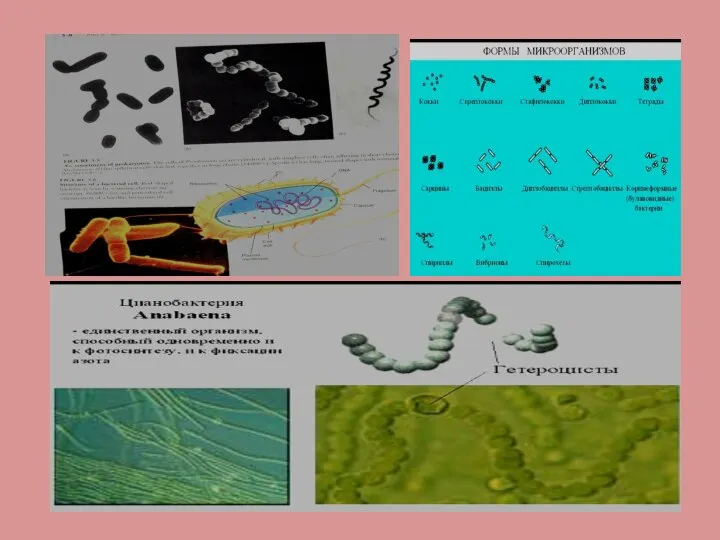

Слайд 4Прокариоты имеют неоформленное ядро (нуклеоид).На ультратонких срезах зона нуклеоида представлена тонкими рыхлыми
Прокариоты имеют неоформленное ядро (нуклеоид).На ультратонких срезах зона нуклеоида представлена тонкими рыхлыми

Слайд 5СТРОЕНИЕ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
СТРОЕНИЕ ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ
Слайд 6 ПОВЕРХНОСТНЫЙ АППАРАТ КЛЕТОК
Общая характеристика поверхностного аппарата
Любая разновидность прокаиотических и
ПОВЕРХНОСТНЫЙ АППАРАТ КЛЕТОК
Общая характеристика поверхностного аппарата
Любая разновидность прокаиотических и

Слайд 7Фосфолипидные структуры, самопроизвольно возникающие в водных растворах: А – мицелла,Б – монослой,
Фосфолипидные структуры, самопроизвольно возникающие в водных растворах: А – мицелла,Б – монослой,

Слайд 9поверхностное натяжение таких пленок намного выше , чем у биологических мембран. При
поверхностное натяжение таких пленок намного выше , чем у биологических мембран. При

Слайд 10Мозаичная модель клеточных мембран
В 1972 г. Синджер и Николсон предложили новую модель
Мозаичная модель клеточных мембран
В 1972 г. Синджер и Николсон предложили новую модель
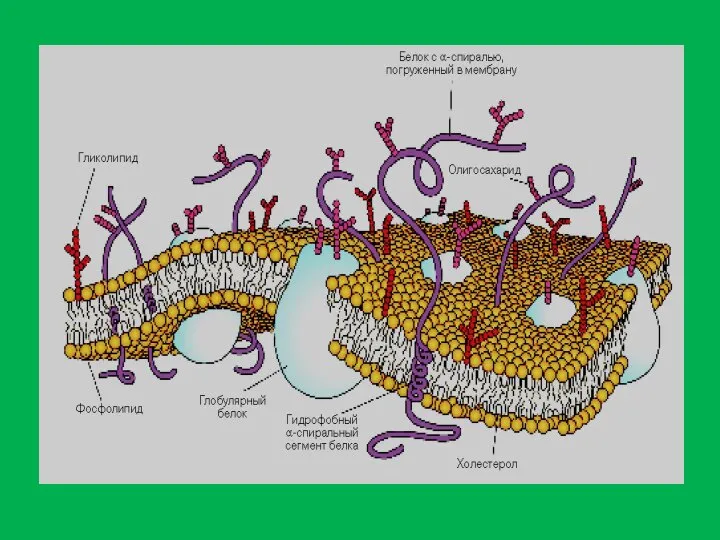
Слайд 12Липиды – органические соединения с различной структурой , но общими свойствами. Они
Липиды – органические соединения с различной структурой , но общими свойствами. Они

Слайд 13Липиды мембран - липидный состав клеточных мембран весьма разнообразен. Характерными представителями липидов
Липиды мембран - липидный состав клеточных мембран весьма разнообразен. Характерными представителями липидов

Слайд 14Жирные кислоты
Жирные кислоты
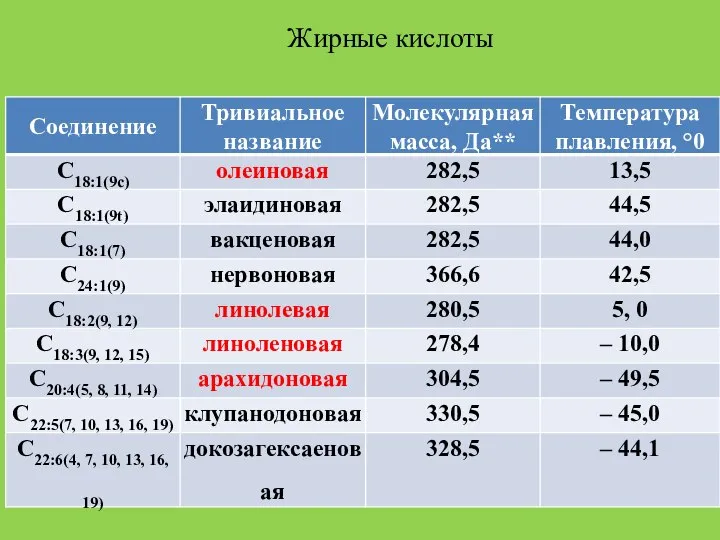
Слайд 15* В классификации жирных кислот вслед за числом углеродных атомов через двоеточие
* В классификации жирных кислот вслед за числом углеродных атомов через двоеточие
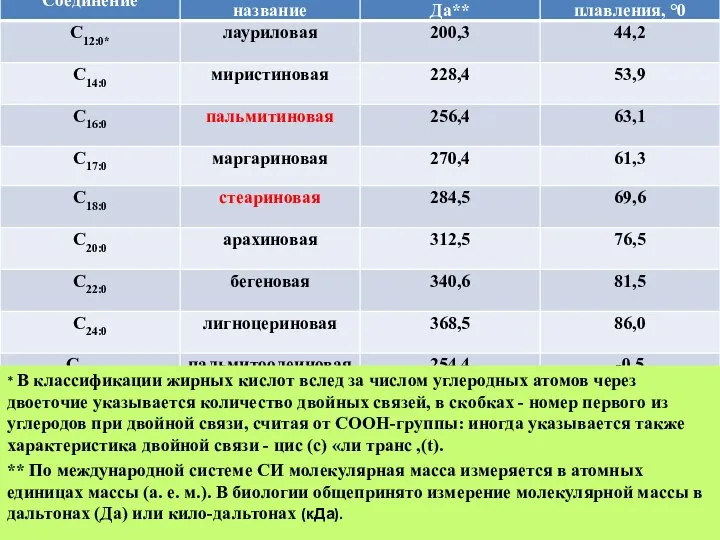
Слайд 16Липидный состав некоторых биологических мембран
Липидный состав некоторых биологических мембран

Слайд 17В состав биологических мембран входят липиды трёх основных классов: фосфолипиды, гликолипиды, стероиды
В состав биологических мембран входят липиды трёх основных классов: фосфолипиды, гликолипиды, стероиды

Слайд 18Гликолипиды мембран представлены цереброзидами, сулфатидами и ганглиозидами. Все они содержат углеводные компоненты.
Цереброзиды
Цереброзиды

Слайд 19Молекула холестерина состоит из компактного, жёсткого гидрофобного ядра, а полярной головкой является
Молекула холестерина состоит из компактного, жёсткого гидрофобного ядра, а полярной головкой является

Слайд 20Липидные плотики (рафты). Холестерин участвует в формировании рафтов — дискретных мембранных доменов, богатых
Липидные плотики (рафты). Холестерин участвует в формировании рафтов — дискретных мембранных доменов, богатых

Слайд 21В липидном бислое выделяют два пула липидов:
аннулярные, или пограничные липиды взаимодействуют
В липидном бислое выделяют два пула липидов:
аннулярные, или пограничные липиды взаимодействуют

Слайд 22Функции липидов мембран состоят в том, что они:
• формируют липидный бислой -
Функции липидов мембран состоят в том, что они:
• формируют липидный бислой -

Слайд 23Липидный бислой мембран находится в динамичном состоянии. Известно 4 вида интрамолекулярных движений
Липидный бислой мембран находится в динамичном состоянии. Известно 4 вида интрамолекулярных движений

Слайд 24при постоянной длине ацильной цепи и температуре увеличение количества двойных связей повышают
при постоянной длине ацильной цепи и температуре увеличение количества двойных связей повышают

Слайд 25а - изменение структуры и свойств мембран при изменении температуры окружающей среды
а - изменение структуры и свойств мембран при изменении температуры окружающей среды

Слайд 26 Р ≥ 10 мкм/с Р ≤ 1 мкм/с Р
Фазовое состояние липидного
Р ≥ 10 мкм/с Р ≤ 1 мкм/с Р
Фазовое состояние липидного

Слайд 27Мембранные белки
Мембраны содержат от 20 до 82 % белка. Белки ответственны за функциональную
Мембранные белки
Мембраны содержат от 20 до 82 % белка. Белки ответственны за функциональную

Слайд 28Периферические белки связаны с поверхностью мембраны за счёт слабых электростатических взаимодействий с
Периферические белки связаны с поверхностью мембраны за счёт слабых электростатических взаимодействий с
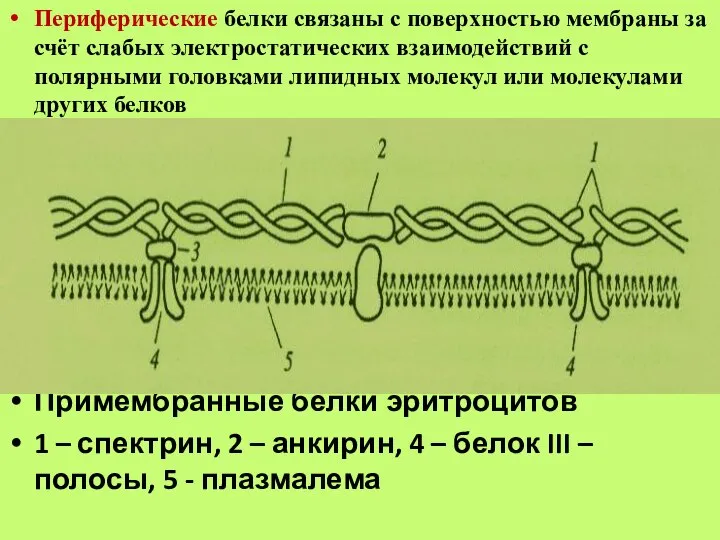
Слайд 29Интегральные, или внутренние, белки погружены в липидный бислой или пронизывают его, фиксируясь
Интегральные, или внутренние, белки погружены в липидный бислой или пронизывают его, фиксируясь

Слайд 30Трансмембранные белки, пересекающие мембрану один раз, относятся к монотопным (например, гликофорин, рецепторы).
Белки,
Трансмембранные белки, пересекающие мембрану один раз, относятся к монотопным (например, гликофорин, рецепторы).
Белки,
Слайд 31Ассиметричное расположение молекул белков в мембранах, определякмое расположени полипептидных цепейем С и
Ассиметричное расположение молекул белков в мембранах, определякмое расположени полипептидных цепейем С и

Слайд 32 Углеводы мембраны
Данная группа веществ составляетпримерно около 10% от общего состава
Углеводы мембраны
Данная группа веществ составляетпримерно около 10% от общего состава

Слайд 33Схематическое изображение клеточной мембраны. С внутренней стороны мембраны виден цитоскелет, с внешней
Схематическое изображение клеточной мембраны. С внутренней стороны мембраны виден цитоскелет, с внешней

Слайд 35Наружная часть содержит преимущественно фосфатидилхолин(75%) , сфингомиелин (80%) , и фосфотидилэтаноламн(20%) .
Наружная часть содержит преимущественно фосфатидилхолин(75%) , сфингомиелин (80%) , и фосфотидилэтаноламн(20%) .
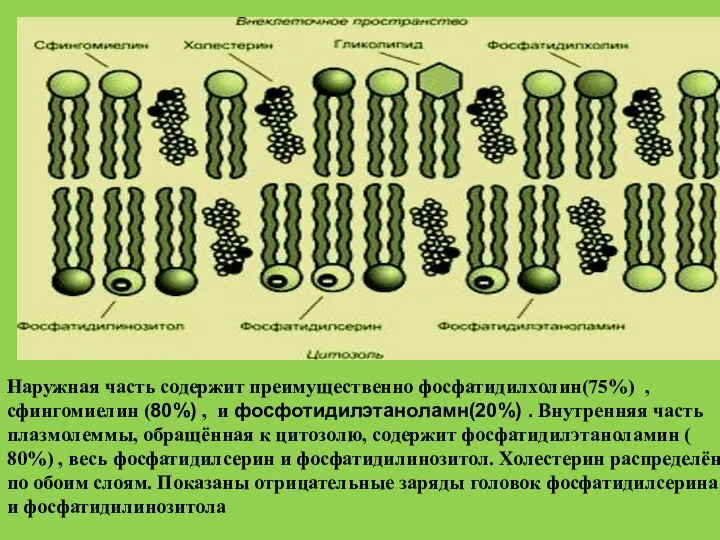
Слайд 3680%сфингомиелина,75%фосфотидилхолина и 20%фосфотидилэтаноламна локализованы на наружной поверхности плазматической мембраны
На внутренней –
80%сфингомиелина,75%фосфотидилхолина и 20%фосфотидилэтаноламна локализованы на наружной поверхности плазматической мембраны
На внутренней –

Слайд 37Слой гликокаликса (ГК) на поверхности плазматической мембраны (ПМ) микроворсинок щеточной каемки энтероцита
Слой гликокаликса (ГК) на поверхности плазматической мембраны (ПМ) микроворсинок щеточной каемки энтероцита
Слайд 38Гликокаликс имеет вид рыхлого волокнистого слоя толщиной 3-4 нм, покрывающего всю поверхность
Гликокаликс имеет вид рыхлого волокнистого слоя толщиной 3-4 нм, покрывающего всю поверхность

Слайд 39Основным компонентом клеточных стенок, значение которого часто недооценивается, является вода. Матрикс клеточной
Основным компонентом клеточных стенок, значение которого часто недооценивается, является вода. Матрикс клеточной

Слайд 40Вода составляет примерно 70% массы большинства живыхорганизмов. Однако содержание ее внутри и
Вода составляет примерно 70% массы большинства живыхорганизмов. Однако содержание ее внутри и
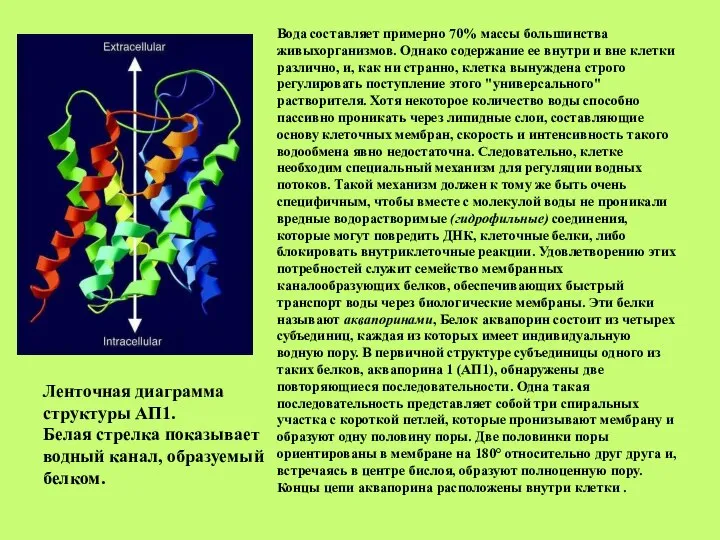
Слайд 41Схема третичной структуры аквапорина:модель «песочные часы»
структура тетрамера
(Из: Ren et al., 200
(из Kjellbom
Схема третичной структуры аквапорина:модель «песочные часы»
структура тетрамера
(Из: Ren et al., 200
(из Kjellbom
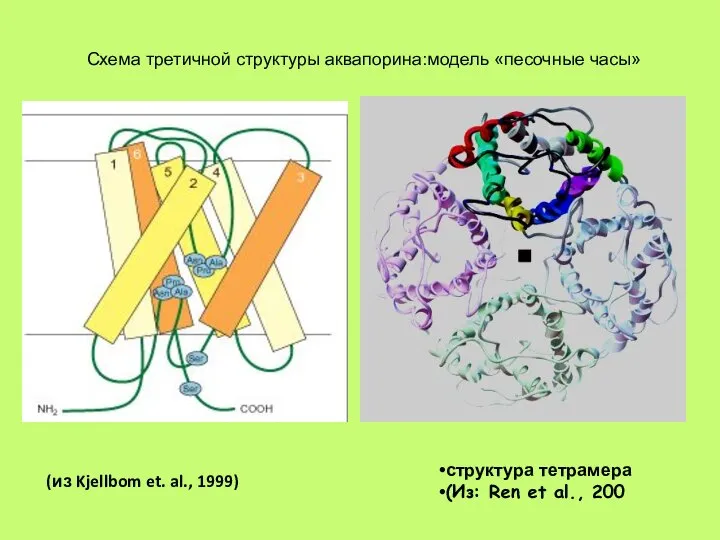
Слайд 42Аквапорины в высшей степени селективны для воды, они не пропускают даже ион
Аквапорины в высшей степени селективны для воды, они не пропускают даже ион

Слайд 43Молекулы целлюлозы (несколько десятков на поперечном срезе)собраны за счет водородных связей в
Молекулы целлюлозы (несколько десятков на поперечном срезе)собраны за счет водородных связей в
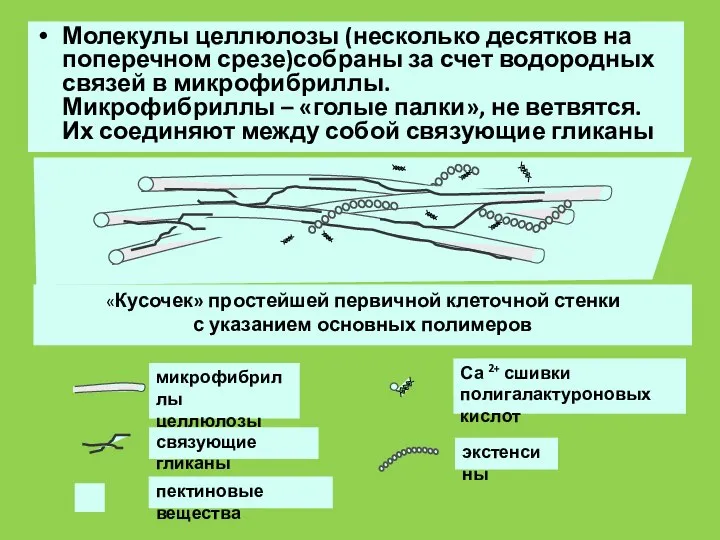
Слайд 44Гликозилтрансферазы, участвующие в формировании полисахаридных молекул, являются мембраносвязанными ферментами.
По характеру взаимодействия
Гликозилтрансферазы, участвующие в формировании полисахаридных молекул, являются мембраносвязанными ферментами.
По характеру взаимодействия
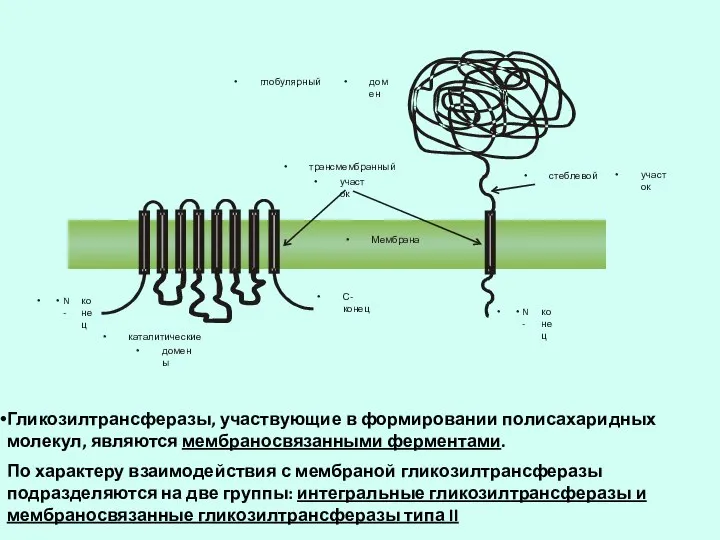
Слайд 45Схема основных видов диффузии через мембрану
Схема основных видов диффузии через мембрану
Слайд 46Транспорт воды через мембрану
( из Maurel & Chrispeels, 2001
Транспорт воды через мембрану
( из Maurel & Chrispeels, 2001
Слайд 47. Вода составляет примерно 70% массы большинства живых организмов. Однако содержание ее
. Вода составляет примерно 70% массы большинства живых организмов. Однако содержание ее
Слайд 48Схема третичной структуры аквапорина:модель «песочные часы»
(из Kjellbom et. al.,
структура тетрамера
(Из: Ren
Схема третичной структуры аквапорина:модель «песочные часы»
(из Kjellbom et. al.,
структура тетрамера
(Из: Ren
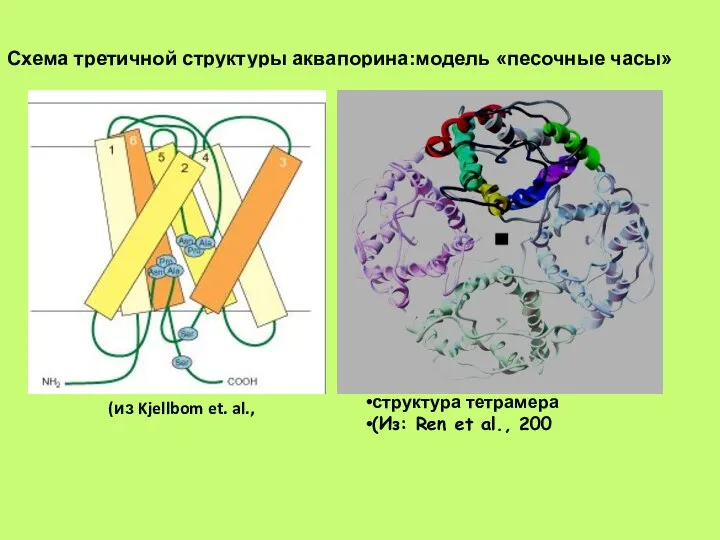
Слайд 49Аквапорины в высшей степени селективны для воды, они не пропускают даже ион
Аквапорины в высшей степени селективны для воды, они не пропускают даже ион

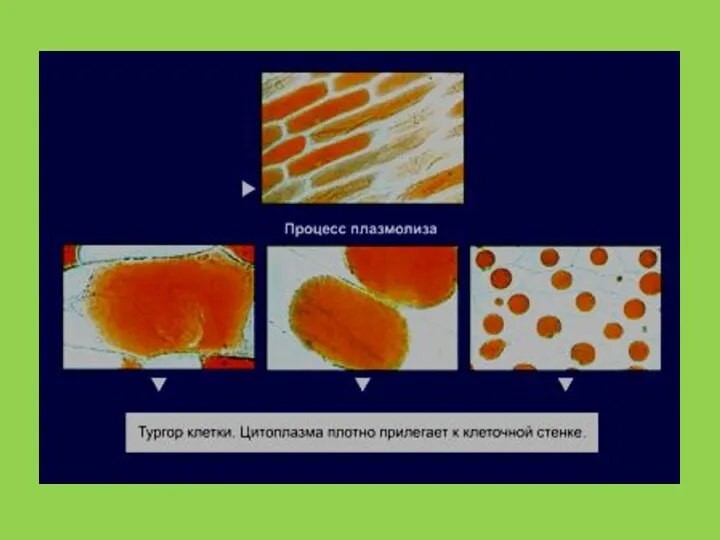
Слайд 51 Пассивный транспорт характеризуется низкой специфичностью. Молекулы в обоих направлениях перемещаются по
Пассивный транспорт характеризуется низкой специфичностью. Молекулы в обоих направлениях перемещаются по


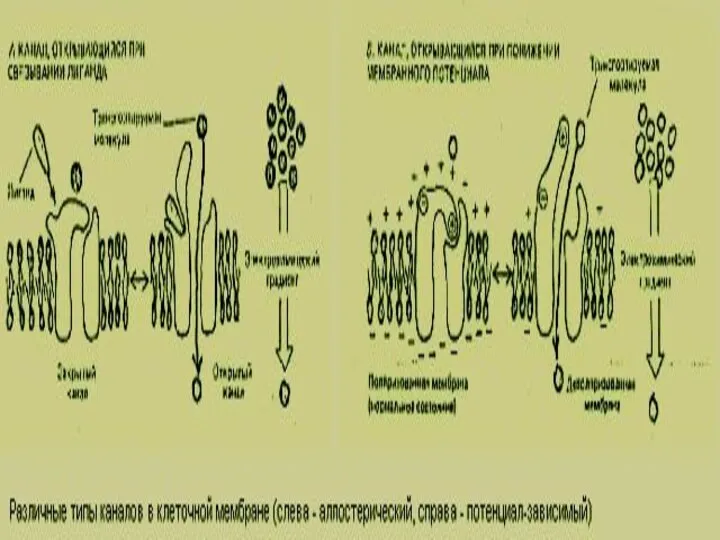
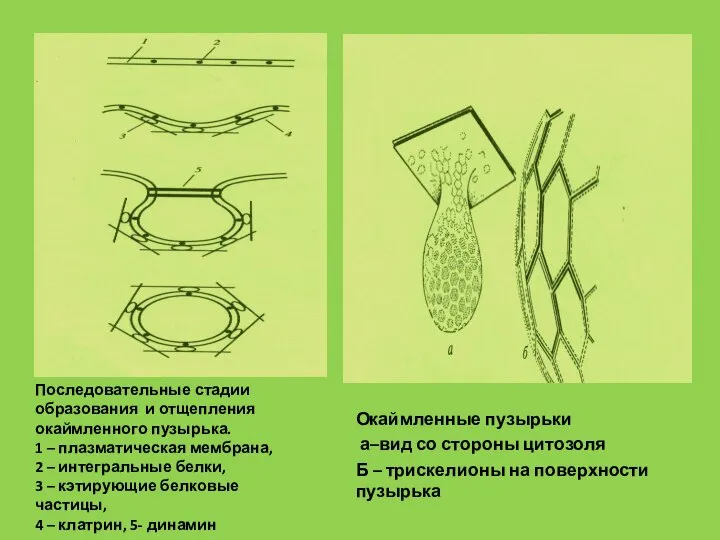
Слайд 54Окаймленные пузырьки
а–вид со стороны цитозоля
Б – трискелионы на поверхности пузырька
Последовательные стадии
Окаймленные пузырьки
а–вид со стороны цитозоля
Б – трискелионы на поверхности пузырька
Последовательные стадии
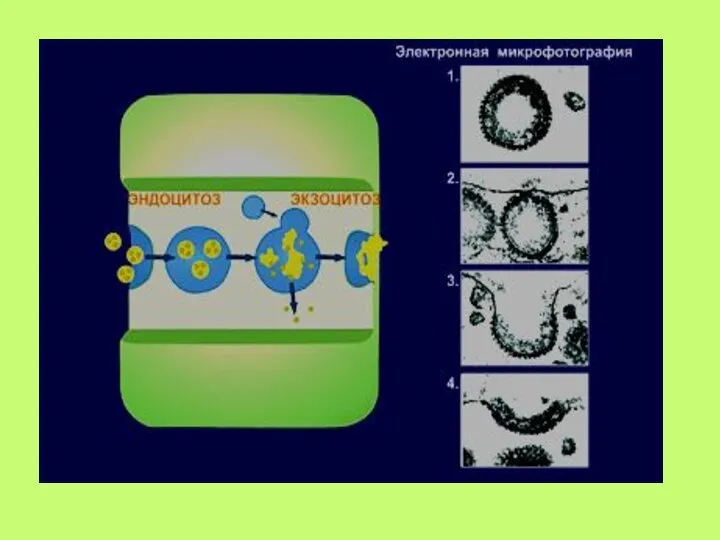
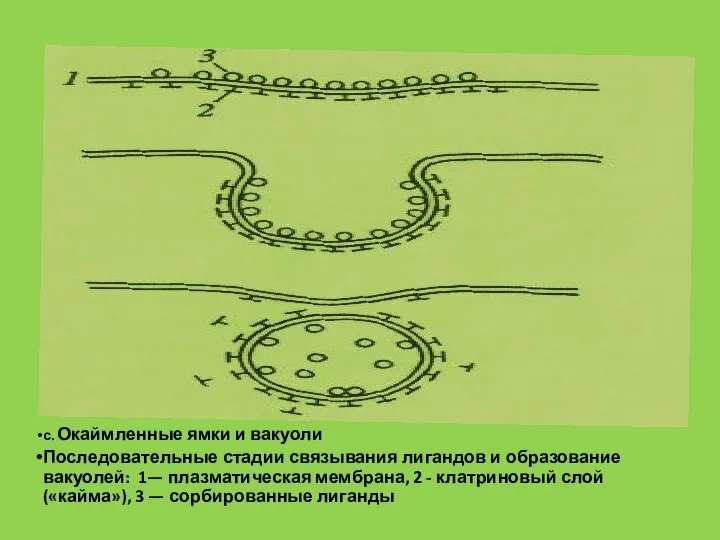
Слайд 56с. Окаймленные ямки и вакуоли
Последовательные стадии связывания лигандов и образование вакуолей: 1—
с. Окаймленные ямки и вакуоли
Последовательные стадии связывания лигандов и образование вакуолей: 1—
Слайд 57. Образование отщепляющейся вакуоли
1- мембрана; 2- рецепторный белок; 3- белок, связывающий ГТФ;
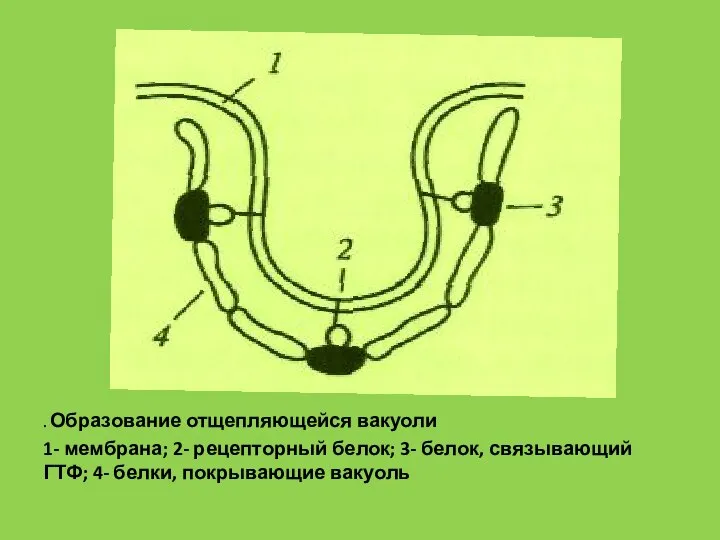
. Образование отщепляющейся вакуоли
1- мембрана; 2- рецепторный белок; 3- белок, связывающий ГТФ;
Слайд 58. Окаймленные пузырьки
а - вид со стороны цитозоля; б - трискелеоны на
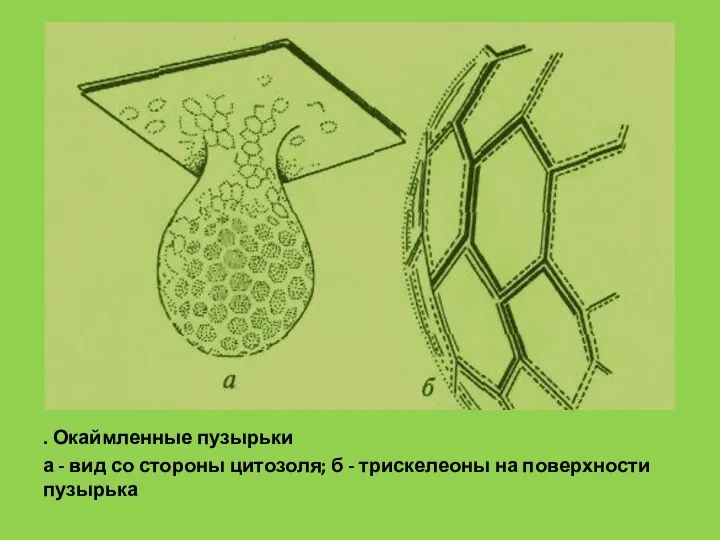
. Окаймленные пузырьки
а - вид со стороны цитозоля; б - трискелеоны на
Слайд 59Поступление в клетку ЛНП(липопротеид низкой плотности) с помощью опосредуемого рецепторами эндоцитоза
1 -
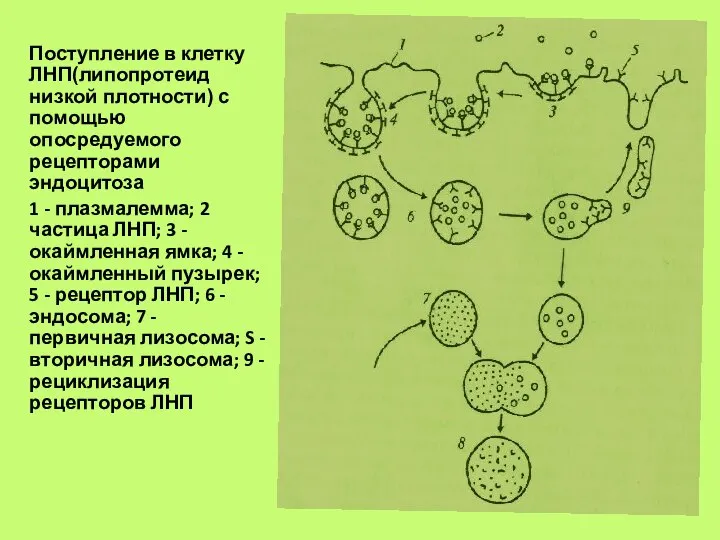
Поступление в клетку ЛНП(липопротеид низкой плотности) с помощью опосредуемого рецепторами эндоцитоза
1 -
Слайд 60 Процесс нахождения транспортной вакуолью (1) мембраны (2), их сближение (5) и
Процесс нахождения транспортной вакуолью (1) мембраны (2), их сближение (5) и
Слайд 61. Схема плотного соединения
а — расположение плотного соединения (вставочная пластинка) на клетках
. Схема плотного соединения
а — расположение плотного соединения (вставочная пластинка) на клетках
Слайд 62Схема простого межклеточного соединения
а - простое соединение, без участия специальных структур; б
Схема простого межклеточного соединения
а - простое соединение, без участия специальных структур; б
Слайд 63Схема строения заякоривающих адгезивных соединений
1- плазматический мембрана; 2-трансмембранные линкерные гликопротеиды;
3 -
Схема строения заякоривающих адгезивных соединений
1- плазматический мембрана; 2-трансмембранные линкерные гликопротеиды;
3 -
Слайд 64Схема щелевого соединения
1 - коннексон; 2- плазматическая мембрана. Стрелка обозначает канал, образованный
Схема щелевого соединения
1 - коннексон; 2- плазматическая мембрана. Стрелка обозначает канал, образованный
 Мадагаскарский дневной геккон, или зелёная фельзума
Мадагаскарский дневной геккон, или зелёная фельзума Пищеварительная система. Ситуационные задачи
Пищеварительная система. Ситуационные задачи Ekzamenatsionnye_preparaty
Ekzamenatsionnye_preparaty Определение сорной и зерновой примесей в кукурузе
Определение сорной и зерновой примесей в кукурузе Внутренняя среда организма. Система кровообращения
Внутренняя среда организма. Система кровообращения Органы чувств и анализаторы
Органы чувств и анализаторы Предупреждение сердечно-сосудистых заболеваний. Первая помощь при кровотечениях
Предупреждение сердечно-сосудистых заболеваний. Первая помощь при кровотечениях Проблемы происхождения человека и первобытная эпоха
Проблемы происхождения человека и первобытная эпоха 1072257
1072257 Сбор углекислого газа из воздуха
Сбор углекислого газа из воздуха Полезные и вредные насекомые
Полезные и вредные насекомые Верблюды. 8 класс
Верблюды. 8 класс Презентация на тему Органические вещества живых организмов
Презентация на тему Органические вещества живых организмов  Животные динозавры
Животные динозавры Прокариотическая клетка
Прокариотическая клетка Гуморальный и клеточный иммунитет. 8 класс
Гуморальный и клеточный иммунитет. 8 класс Экологические группы растений по отношению к влажности
Экологические группы растений по отношению к влажности 8 февраля – День российской науки
8 февраля – День российской науки Рептилии. Внешнее строение
Рептилии. Внешнее строение Гребенчатый орел
Гребенчатый орел Теория рефлекторная деятельность внешней временной связи
Теория рефлекторная деятельность внешней временной связи Органы пищеварения. Пищеварение
Органы пищеварения. Пищеварение Презентация на тему Зоология наука о животных
Презентация на тему Зоология наука о животных  Презентация на тему Кроссворд Происхождение человека
Презентация на тему Кроссворд Происхождение человека  Размножение живых организмов
Размножение живых организмов Тип Моллюски
Тип Моллюски Лесные гости
Лесные гости Биоэлемент фосфор для животных
Биоэлемент фосфор для животных