- Клеточные механизмы запуска мышечного сокращения. Лекция № 11

Содержание
- 2. 1. Назовите три типа нитей в саркомере поперечно-полосатых мышц. 2. Какой белок формирует третий тип нитей?
- 3. Новости науки Раскрыта причина непобедимости рака Ученые Калифорнийского университета в Сан-Диего выяснили, что в опухолевых клетках
- 4. НОВЫЙ АППАРАТ ОТ TOSHIBA ПО КАПЛЕ КРОВИ ВЫЯВЛЯЕТ 13 ВИДОВ РАКА Эксперты из компании Toshiba создали
- 5. Ну теперь к нашим баранам…
- 6. Три основных события, необходимых для реализации сокращения любой мышцы: Деполяризация мышечной мембраны и генерация потенциала действия
- 7. Внутриклеточный метаболизм ионов кальция Внутриклеточный кальций присутствует в мышечных клетках в основном в связанном состоянии, и
- 8. Посчитаем!
- 9. Т.о., мышечные клетки и волокна в состоянии покоя имеют ~ одинаковый уровень ионов кальция в миоплазме
- 10. Механизмы, поддерживающие низкий уровень кальция в покое (кстати, зачем?) кальций-зависимая АТРаза мембраны клетки – кальциевый насос
- 11. Повышение уровня кальция при сокращении В покое концентрация ионов кальция в миоплазме = ~ 5х10 (в
- 12. У скелетных волокон и кардиомиоцитов главным источником прироста кальция, запускающего сокращение, является СР. В СР есть
- 13. Открывание кальциевого канала в составе молекулы рианодинового рецептора происходит при действии на РиР ионов кальция (в
- 14. В кардиомиоцитах акт сокращения также запускается за счет высвобождения кальция из цистерн СР. Но! Важным источником
- 15. В гладких мышечных клетках СР развит слабо. Пути повышения концентрации кальция в миоплазме: Поступление в клетку
- 16. Особенности генерации потенциала действия (ПД) в разных типах мышечных клеток В скелетных мышечных волокнах позвоночных восходящая
- 17. В сердечных миоцитах восходящая фаза ПД обусловлена входящим натриевым током и!!! кальциевым током! Имеется фаза «плато
- 18. У гладкомышечных клеток, как и у кардиомиоцитов, ПД имеет три фазы: деполяризацию, плато и реполяризацию. Длительность
- 19. Особенности ЭМС в разных мышцах (обобщение и дополнение) 1 – потенциалзависимый выброс кальция в скелетных мышцах
- 20. Фармако-механическое сопряжение в гладких мышцах
- 21. Регуляция взаимодействия актина и миозина на уровне сократительного аппарата
- 22. Миозин - наномотор, гидролиз АТФ в моторном домене и развитие усилия
- 23. Кальций-связывающие белки (EF-hand) – обеспечивают инициацию сокращения в поперечно-полосатых и гладких мышцах EF-hand-белки условно подразделяются на
- 24. Самые простые по строению кальций-связывающие белки содержат в своем составе только 2 EF-hand домена: S-100 (из
- 25. К EF-hand-белкам относятся также: альфа-актинин, дистрофин! Группа белков, содержащая 6 и более центров связывания ионов кальция
- 26. Ионы магния имеют меньший радиус, чем ионы кальция. Вследствие этого они менее прочно связываются в катионсвязывающих
- 27. EF-hand-белки участвуют в регуляции сократительной активности поперечно-полосатых и гладких мышц! Эти белки связаны в мышце либо
- 28. 2 Тип миозиновой регуляции – конформационные изменения в области шейки головки формируются путем фосфорилирования регуляторных легких
- 29. Актиновый тип регуляции - основной тип регуляции мышечного сокращения в поперечно-полосатых мышцах животных. В основе этого
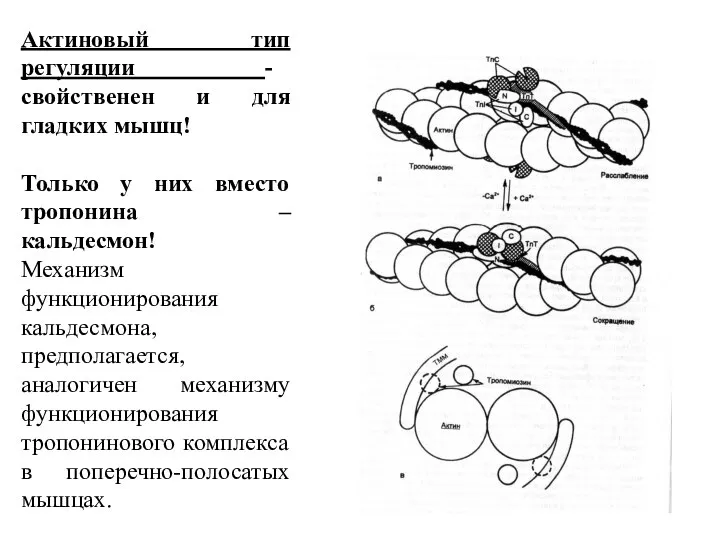
- 30. Актиновый тип регуляции - свойственен и для гладких мышц! Только у них вместо тропонина – кальдесмон!
- 34. Т.о., актиновый и миозиновый типы регуляции свойственны для всех типов мышц!
- 35. Следующая лекция
- 37. Скачать презентацию

Слайд 3Новости науки
Раскрыта причина непобедимости рака
Ученые Калифорнийского университета в Сан-Диего выяснили, что
Новости науки
Раскрыта причина непобедимости рака
Ученые Калифорнийского университета в Сан-Диего выяснили, что
Слайд 4НОВЫЙ АППАРАТ ОТ TOSHIBA ПО КАПЛЕ КРОВИ ВЫЯВЛЯЕТ 13 ВИДОВ РАКА
Эксперты из
НОВЫЙ АППАРАТ ОТ TOSHIBA ПО КАПЛЕ КРОВИ ВЫЯВЛЯЕТ 13 ВИДОВ РАКА
Эксперты из

Слайд 5Ну теперь к нашим баранам…
Ну теперь к нашим баранам…
Слайд 6Три основных события, необходимых для реализации сокращения любой мышцы:
Деполяризация мышечной мембраны
Три основных события, необходимых для реализации сокращения любой мышцы:
Деполяризация мышечной мембраны

Слайд 7Внутриклеточный метаболизм ионов кальция
Внутриклеточный кальций присутствует в мышечных клетках в основном в
Внутриклеточный метаболизм ионов кальция
Внутриклеточный кальций присутствует в мышечных клетках в основном в

Слайд 8Посчитаем!
Посчитаем!

Слайд 9Т.о., мышечные клетки и волокна в состоянии покоя имеют ~ одинаковый уровень
Т.о., мышечные клетки и волокна в состоянии покоя имеют ~ одинаковый уровень

Слайд 10Механизмы, поддерживающие низкий уровень кальция в покое (кстати, зачем?)
кальций-зависимая АТРаза мембраны клетки
Механизмы, поддерживающие низкий уровень кальция в покое (кстати, зачем?)
кальций-зависимая АТРаза мембраны клетки

Слайд 11Повышение уровня кальция при сокращении
В покое концентрация ионов кальция в миоплазме =
Повышение уровня кальция при сокращении
В покое концентрация ионов кальция в миоплазме =

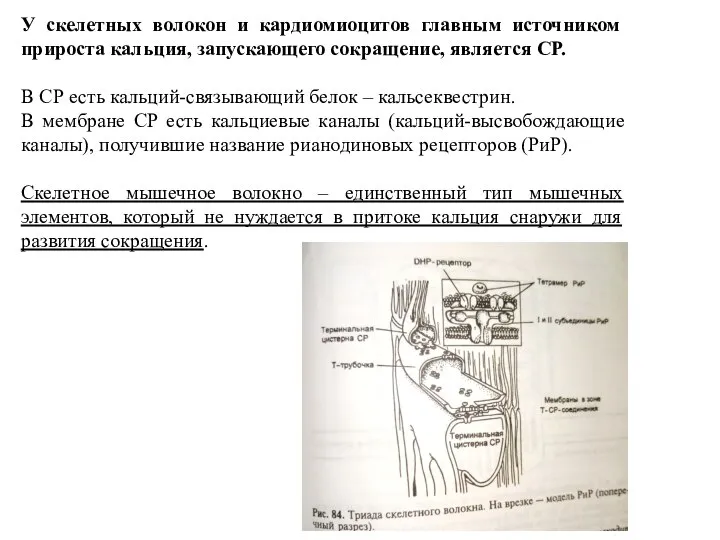
Слайд 12У скелетных волокон и кардиомиоцитов главным источником прироста кальция, запускающего сокращение, является
У скелетных волокон и кардиомиоцитов главным источником прироста кальция, запускающего сокращение, является
Слайд 13Открывание кальциевого канала в составе молекулы рианодинового рецептора происходит при действии на
Открывание кальциевого канала в составе молекулы рианодинового рецептора происходит при действии на

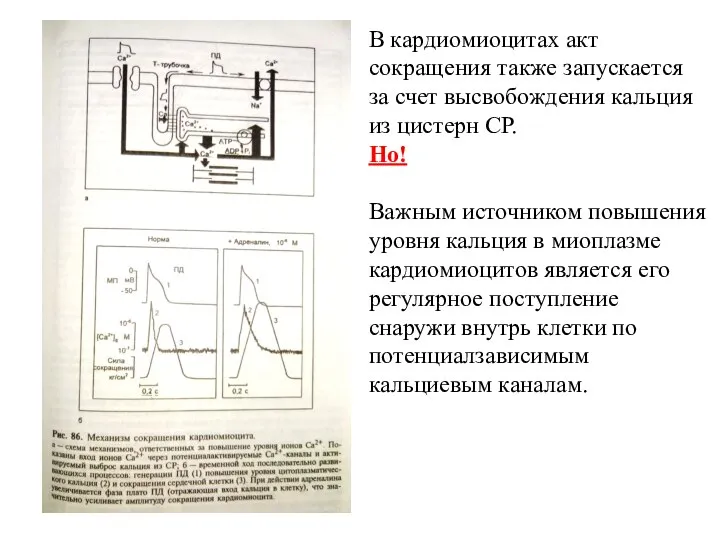
Слайд 14В кардиомиоцитах акт сокращения также запускается за счет высвобождения кальция из цистерн
В кардиомиоцитах акт сокращения также запускается за счет высвобождения кальция из цистерн
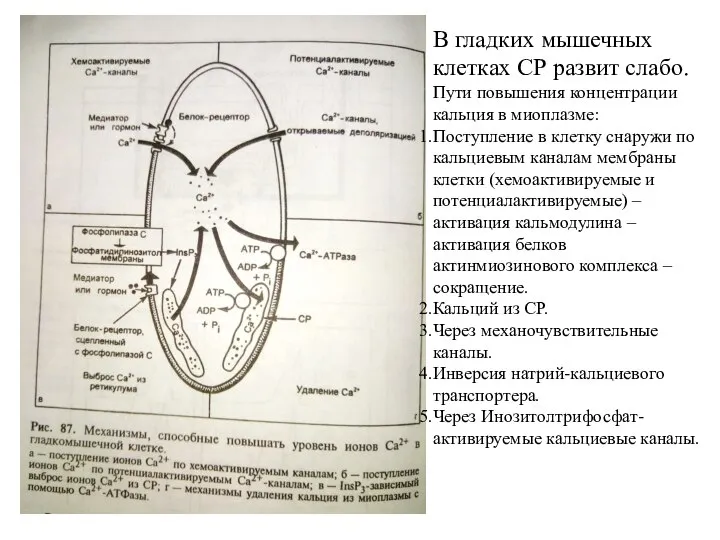
Слайд 15В гладких мышечных клетках СР развит слабо.
Пути повышения концентрации кальция в миоплазме:
В гладких мышечных клетках СР развит слабо.
Пути повышения концентрации кальция в миоплазме:
Слайд 16Особенности генерации потенциала действия (ПД) в разных типах мышечных клеток
В скелетных
Особенности генерации потенциала действия (ПД) в разных типах мышечных клеток
В скелетных

Слайд 17В сердечных миоцитах восходящая фаза ПД обусловлена входящим натриевым током и!!!
кальциевым током!
В сердечных миоцитах восходящая фаза ПД обусловлена входящим натриевым током и!!!
кальциевым током!

Слайд 18У гладкомышечных клеток, как и у кардиомиоцитов, ПД имеет три фазы: деполяризацию,
У гладкомышечных клеток, как и у кардиомиоцитов, ПД имеет три фазы: деполяризацию,

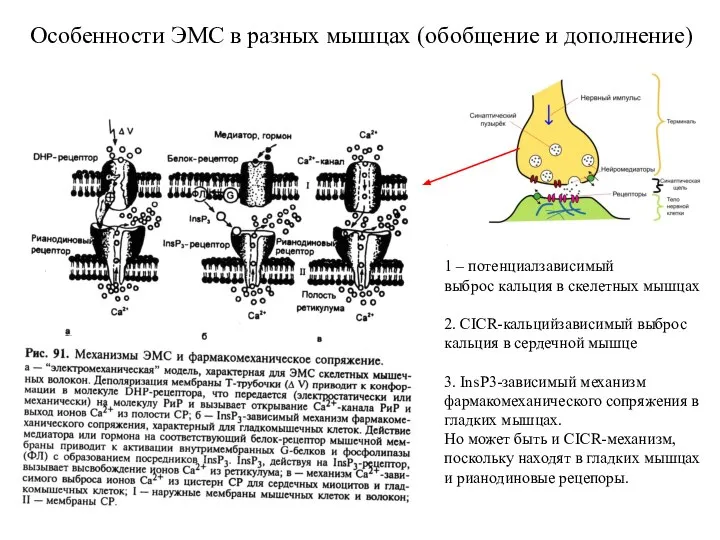
Слайд 19Особенности ЭМС в разных мышцах (обобщение и дополнение)
1 – потенциалзависимый
выброс
Особенности ЭМС в разных мышцах (обобщение и дополнение)
1 – потенциалзависимый
выброс
Слайд 20Фармако-механическое сопряжение в гладких мышцах
Фармако-механическое сопряжение в гладких мышцах

Слайд 21Регуляция взаимодействия актина и миозина на уровне сократительного аппарата
Регуляция взаимодействия актина и миозина на уровне сократительного аппарата

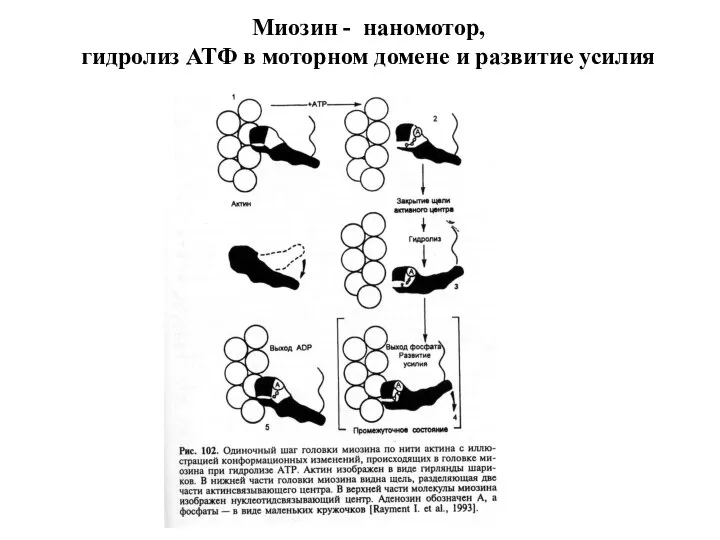
Слайд 22Миозин - наномотор,
гидролиз АТФ в моторном домене и развитие усилия
Миозин - наномотор,
гидролиз АТФ в моторном домене и развитие усилия
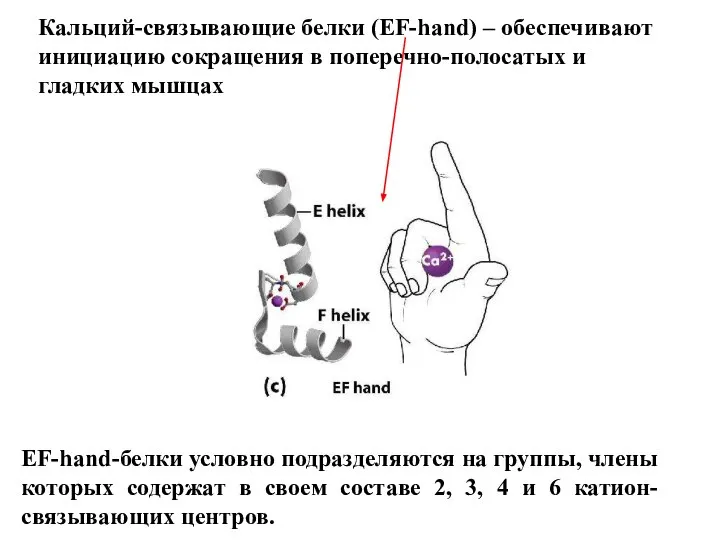
Слайд 23Кальций-связывающие белки (EF-hand) – обеспечивают
инициацию сокращения в поперечно-полосатых и
гладких мышцах
EF-hand-белки
Кальций-связывающие белки (EF-hand) – обеспечивают
инициацию сокращения в поперечно-полосатых и
гладких мышцах
EF-hand-белки
Слайд 24Самые простые по строению кальций-связывающие белки содержат в своем составе только 2
Самые простые по строению кальций-связывающие белки содержат в своем составе только 2

Слайд 25К EF-hand-белкам относятся также:
альфа-актинин, дистрофин!
Группа белков, содержащая 6 и более
К EF-hand-белкам относятся также:
альфа-актинин, дистрофин!
Группа белков, содержащая 6 и более

Слайд 26Ионы магния имеют меньший радиус, чем ионы кальция. Вследствие этого они менее
Ионы магния имеют меньший радиус, чем ионы кальция. Вследствие этого они менее

Слайд 27EF-hand-белки участвуют в регуляции сократительной активности поперечно-полосатых и гладких мышц!
Эти белки связаны
EF-hand-белки участвуют в регуляции сократительной активности поперечно-полосатых и гладких мышц!
Эти белки связаны

Слайд 282 Тип миозиновой регуляции – конформационные изменения в области шейки головки формируются
2 Тип миозиновой регуляции – конформационные изменения в области шейки головки формируются

Слайд 29Актиновый тип регуляции - основной тип регуляции мышечного сокращения в поперечно-полосатых мышцах
Актиновый тип регуляции - основной тип регуляции мышечного сокращения в поперечно-полосатых мышцах

Слайд 30Актиновый тип регуляции - свойственен и для гладких мышц!
Только у них вместо
Актиновый тип регуляции - свойственен и для гладких мышц!
Только у них вместо


Слайд 34Т.о., актиновый и миозиновый типы регуляции свойственны для всех типов мышц!
Т.о., актиновый и миозиновый типы регуляции свойственны для всех типов мышц!

Слайд 35Следующая лекция
Следующая лекция

 Многообразие цветковых растений
Многообразие цветковых растений Создание гербария Травки-муравки
Создание гербария Травки-муравки Морской мир
Морской мир Вегетативная нервная система человека. (Лекция 5)
Вегетативная нервная система человека. (Лекция 5) Проращивание овса
Проращивание овса Животный мир
Животный мир Презентация на тему Ядовитые животные
Презентация на тему Ядовитые животные  Павлов Иван Петрович (фотографии)
Павлов Иван Петрович (фотографии) Koala as an endangered species
Koala as an endangered species Рыбы - обитатели водных пространств
Рыбы - обитатели водных пространств Экологические группы водорослей
Экологические группы водорослей Ткани человека
Ткани человека Анатомия собаки. Скелет, зубная система
Анатомия собаки. Скелет, зубная система Земноводные или амфибии
Земноводные или амфибии Тля и белокрылка
Тля и белокрылка Дисграфия. Методы коррекции
Дисграфия. Методы коррекции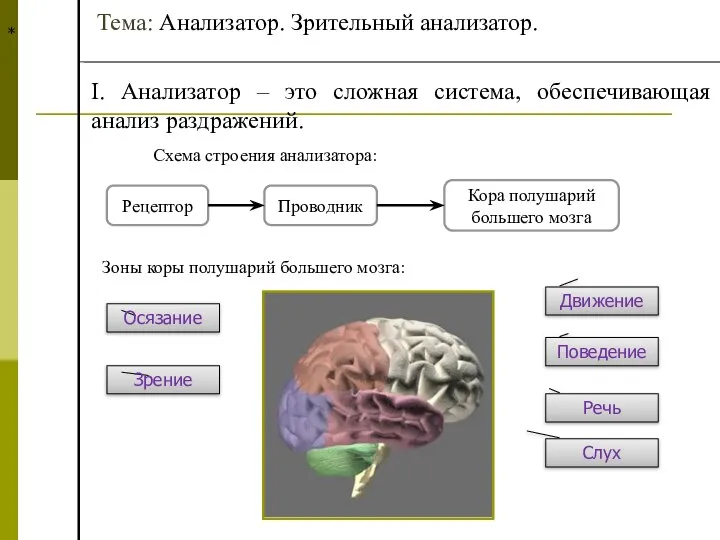 Анализатор. Зрительный анализатор
Анализатор. Зрительный анализатор Красная книга России
Красная книга России Применение водорослей в медицине
Применение водорослей в медицине Вирусы. Особенности строения и размножения. Значение в природе и жизни человека
Вирусы. Особенности строения и размножения. Значение в природе и жизни человека Воздушное питание, хлоропласты, фотосинтез, растения-паразиты. Питание животных
Воздушное питание, хлоропласты, фотосинтез, растения-паразиты. Питание животных Презентация на тему РЕКОРДЫ СОБАЧЬЕГО МИРА
Презентация на тему РЕКОРДЫ СОБАЧЬЕГО МИРА  Презентация на тему Взаимодействие неаллельных генов
Презентация на тему Взаимодействие неаллельных генов  Екосистеми, їх структура та властивості
Екосистеми, їх структура та властивості Эффект кворума
Эффект кворума Звуки в лесу
Звуки в лесу Вид. Критерии вида
Вид. Критерии вида Человек и биосфера. Деятельность человека и эволюция биосферы
Человек и биосфера. Деятельность человека и эволюция биосферы