- Макростехиометрические характеристики биотехнологического процесса

Содержание
- 2. Важное значение имеют макростехиометрические характеристики биотехнологического процесса. Они выражают взаимосвязь между приростом биомассы, продукта и расходованием
- 3. выход по субстрату, или экономический коэффициент (или коэффициент выхода). определяют, сравнивая количество выросшей за весь цикл
- 4. Выход по продукту метаболизма: Yps=Рк/S0 Различия в обозначении коэффициентов делают с помощью индексов: по биомассе обозначают
- 5. дополнения коэффициент Yxs определен не совсем точно. В начале процесса уже существует некоторое количество биомассы, определяемое
- 6. Экономические коэффициенты: Yxs = (Xk – X0)/ (S0 – Sk) = ΔX/ΔS [г. биомассы/ г. субстрата]
- 7. Метаболические, или трофические, коэффициенты: Ysx = 1/Yxs = (S0 – Sk)/ (Xk – X0) = ΔS/ΔX
- 8. В пределе можно рассматривать промежуток ∆t сколь угодно малым — вплоть до бесконечно малого dt, и
- 9. Стехиометрия процессов культивирования микроорганизмов
- 10. Основные принципы стехиометрии Экономический, метаболический коэффициенты выхода продукта по субстрату и биомассе — это в простейшем
- 11. Если в результате взаимодействия реагентов А и В получаются продукты С и D и выделяется тепло
- 12. Стехиометрические коэффициенты подбирают таким образом, чтобы выразить фундаментальный закон природы — закон сохранения материи. Количество атомов
- 13. В биологии также действует закон сохранения материи. В ходе биологических превращений в клетке перегруппировываются атомы углерода,
- 14. Процесс ферментации можно представить как систему, в которой происходит преобразование исходных реагентов (субстратов) в продукты (клетки
- 15. по аналогии со стехиометрией в микробиологическом процессе ʋc [углеродный субстрат] + +ʋN [азотный субстрат] + +ʋф
- 16. ВЫВОД «ФОРМУЛЫ» БИОМАССЫ МИКРООРГАНИЗМОВ биомасса, состоящая из множества индивидуальных веществ — белков, нуклеиновых кислот, липидов и
- 17. какой-то существующей в природе истинной «формулы» биомассы нет. нужен фиктивный, эмпирический вид «формулы» биомассы За основу
- 18. элементный состав микроорганизмов
- 19. Как теперь из этого элементного состава получить «формулу» биомассы? Если принять сухую массу биомассы равной 100
- 20. Расчет числа грамм-атомов элементов в 100 г сухой биомассы тип м/о С Н О N P
- 21. в первом приближении «формулу» дрожжей можно записать как С3,92Н6,5О1,88N0,54P0,05S0,03 бактерий — С4,42Н7О1,25N0,86P0,1S0,03 «усредненной» биомассы — С4,17Н8О1,25N1P0,1S0,03
- 22. чтобы придать видимость правдоподобия «формуле», ее умножают на какое-то очень большое число, чтобы все коэффициенты оказались
- 23. в стехиометрических расчетах обычно пренебрегают элементами, составляющими малую часть состава биомассы. В «формуле» отбрасывают фосфор и
- 24. в стехиометрическом уравнении все члены можно умножить или разделить на одно и то же число. без
- 25. Удобно принимать такую «молекулярную массу», чтобы в ней оказался только один атом (грамм-атом) углерода. Для этого
- 26. дрожжи — CH1,66O0,48N 0,14, бактерии — CH1,58O0,28N0,19; «усредненная» биомасса — CH1,92O0,30N0,24;
- 27. для «усредненной» биомассы используется формула, предложенная Стоутхамером для С-моля: CH1,8O0,5N0,2 Из-за простоты эту формулу применяют в
- 28. Рассчет «молекулярной массы» С-моля: М= 1·12 + 1,8·1 + 0,5·16 + 0,2·14 = 24,6 по брутто-формуле
- 29. РАСЧЕТ ВЫХОДА БИОМАССЫ НА УГЛЕРОДНЫЙ СУБСТРАТ наибольший интерес с учетом выхода составляет самый дорогой субстрат —
- 30. Эти вещества можно также пересчитать на С-моль (т.е. оставить в молекуле только один атом углерода). Например,
- 31. разные субстраты дают различный стехиометрический выход по биомассе. фактически измеренные данные выхода биомассы отличаются от теоретически
- 32. Чтобы объяснить эти расхождения, ввели понятие «энергетический выход» биомассы. - в каждом субстрате заключена энергия, которая
- 33. Степень восстановленности зависит от числа так называемых «доступных электронов», или «редоксонов». Принимают, что один атом углерода
- 34. степень восстановленности субстрата CmHnOpNq рассчитывают по формуле: γs = 4m + n - 2р - 3q
- 35. Оценка биомассы по степени восстановленности (обозначение γх) Биомасса (CHl,8O0,5N0,2): γX =4 х 1 + 1,8 –
- 36. В.К. Ерошиным введено понятие энергетический выход биомассы, представляющий собой отношение энергии в субстрате к энергии, заключенной
- 37. метан η = 8,0/4,2 = 1,9 С-моль биомассы/C-моль субстрата; гексан η = 6,3/4,2 = 1,5 метанол
- 38. В формуле для η выход выражен в С-молях биомассы на С-моль субстрата. От него можно перейти
- 39. метан YXS = 1,9 х 24,6/16 = 2,92 г биомассы/ г субстрата; гексан YXS = 1,5
- 40. для глюкозы энергетический выход биомассы дает значения чуть ниже стехиометрического (0,78 против 0,82), но при этом
- 41. ОПРЕДЕЛЕНИЕ СТЕХИОМЕТРИЧЕСКИХ СООТНОШЕНИЙ В РЕАЛЬНЫХ ПРОЦЕССАХ ФЕРМЕНТАЦИИ исходные предпосылки «энергетической» теории материального баланса неверны. Они не
- 42. общее стехиометрическое соотношение для объединенного процесса: ʋs S + ʋo2 [02] + ʋN [NH3] → X
- 43. Необходимо знать, количество израсходованного субстрата (в реальных мерах — граммы, килограммы и т. д.) и количество
- 44. Разделив эти величины на молекулярные массы соответственно субстрата, биомассы или продукта, получаем для этих веществ количества
- 45. Первый из этих расчетов — приведение всех количеств к одному С-молю биомассы. обычно находят такие стехиометрические
- 46. Таким образом сразу находим ʋs и ʋР. Для определения коэффициентов по другим веществам (О2, СО2, NH3
- 47. При этом индексы в формуле субстрата обозначим как ms, ns, ps, qs, в формуле продукта —
- 48. уравнение баланса по углероду: ʋSmS = 1 + ʋpmp + ʋCO2 То же для водорода: ʋSnS
- 49. Уравнение баланса по кислороду: ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp + ʋCO2 ·
- 50. Решение системы уравнений позволит найти все стехиометрические коэффициенты для данного процесса ферментации. Для примера рассмотрим конкретный
- 51. Переведем все величины в г моли соответствующих веществ Молекулярная масса (г/моль): сахарозы (С12Н22О11) равна 342, лимонной
- 52. Пересчитаем все стехиометрические коэффициенты на С-моль биомассы, т.е. разделим обе части равенства на 12,2. 0,24 [С12Н22О11]
- 53. Для нахождения оставшихся коэффи-циентов записываем систему уравнений элементного баланса для данного процесса При этом для субстрата
- 54. Баланс по водороду: ʋSnS + ʋNH3 · 3 = 1,8 + ʋpnp + ʋH2O · 2
- 55. Баланс по кислороду: ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp + ʋCO2 · 2
- 56. Баланс по азоту: ʋsqs + ʋNH3 = 0,2 + ʋpqp 0 + ʋNH3 = 0,2 +
- 58. Скачать презентацию
Слайд 2Важное значение имеют макростехиометрические характеристики биотехнологического процесса.
Они выражают взаимосвязь между приростом биомассы,
Важное значение имеют макростехиометрические характеристики биотехнологического процесса.
Они выражают взаимосвязь между приростом биомассы,

Слайд 3выход по субстрату, или экономический коэффициент (или коэффициент выхода).
определяют, сравнивая количество
выход по субстрату, или экономический коэффициент (или коэффициент выхода).
определяют, сравнивая количество

Слайд 4Выход по продукту метаболизма:
Yps=Рк/S0
Различия в обозначении коэффициентов делают с помощью
Выход по продукту метаболизма:
Yps=Рк/S0
Различия в обозначении коэффициентов делают с помощью

Слайд 5
дополнения
коэффициент Yxs определен не совсем точно. В начале процесса уже
дополнения
коэффициент Yxs определен не совсем точно. В начале процесса уже

Слайд 6Экономические коэффициенты:
Yxs = (Xk – X0)/ (S0 – Sk) = ΔX/ΔS
[г.
Экономические коэффициенты:
Yxs = (Xk – X0)/ (S0 – Sk) = ΔX/ΔS
[г.

Слайд 7Метаболические, или трофические, коэффициенты:
Ysx = 1/Yxs = (S0 – Sk)/ (Xk –
Метаболические, или трофические, коэффициенты:
Ysx = 1/Yxs = (S0 – Sk)/ (Xk –

Слайд 8 В пределе можно рассматривать промежуток ∆t сколь угодно малым — вплоть
В пределе можно рассматривать промежуток ∆t сколь угодно малым — вплоть

Слайд 9Стехиометрия процессов культивирования
микроорганизмов
Стехиометрия процессов культивирования
микроорганизмов

Слайд 10Основные принципы
стехиометрии
Экономический, метаболический коэффициенты выхода продукта по субстрату и биомассе
Основные принципы
стехиометрии
Экономический, метаболический коэффициенты выхода продукта по субстрату и биомассе

Слайд 11
Если в результате взаимодействия реагентов А и В получаются продукты С
Если в результате взаимодействия реагентов А и В получаются продукты С

Слайд 12 Стехиометрические коэффициенты подбирают таким образом, чтобы выразить фундаментальный закон природы —
Стехиометрические коэффициенты подбирают таким образом, чтобы выразить фундаментальный закон природы —

Слайд 13В биологии также действует закон сохранения материи.
В ходе биологических превращений
В биологии также действует закон сохранения материи.
В ходе биологических превращений

Слайд 14 Процесс ферментации можно представить как систему, в которой происходит преобразование исходных
Процесс ферментации можно представить как систему, в которой происходит преобразование исходных

Слайд 15по аналогии со стехиометрией в микробиологическом процессе
ʋc [углеродный субстрат] +
+ʋN [азотный
по аналогии со стехиометрией в микробиологическом процессе
ʋc [углеродный субстрат] +
+ʋN [азотный
![по аналогии со стехиометрией в микробиологическом процессе ʋc [углеродный субстрат] + +ʋN](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/975173/slide-14.jpg)
Слайд 16ВЫВОД «ФОРМУЛЫ» БИОМАССЫ МИКРООРГАНИЗМОВ
биомасса, состоящая из множества индивидуальных веществ — белков,
ВЫВОД «ФОРМУЛЫ» БИОМАССЫ МИКРООРГАНИЗМОВ
биомасса, состоящая из множества индивидуальных веществ — белков,

Слайд 17 какой-то существующей в природе истинной «формулы» биомассы нет.
нужен фиктивный, эмпирический
какой-то существующей в природе истинной «формулы» биомассы нет.
нужен фиктивный, эмпирический

Слайд 18элементный состав микроорганизмов
элементный состав микроорганизмов
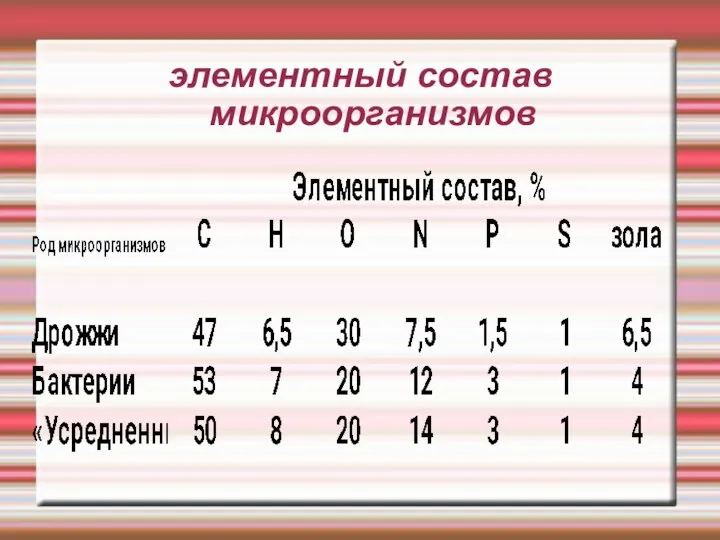
Слайд 19Как теперь из этого элементного состава получить «формулу» биомассы?
Если принять сухую
Как теперь из этого элементного состава получить «формулу» биомассы?
Если принять сухую

Слайд 20Расчет числа грамм-атомов элементов в 100 г сухой биомассы
тип м/о С Н
Расчет числа грамм-атомов элементов в 100 г сухой биомассы
тип м/о С Н
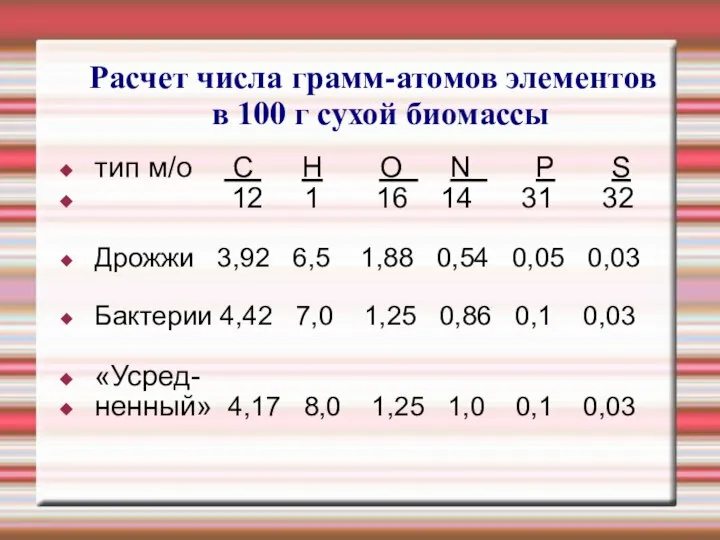
Слайд 21 в первом приближении «формулу» дрожжей можно записать как
С3,92Н6,5О1,88N0,54P0,05S0,03
бактерий —
в первом приближении «формулу» дрожжей можно записать как
С3,92Н6,5О1,88N0,54P0,05S0,03
бактерий —

Слайд 22чтобы придать видимость правдоподобия «формуле», ее умножают на какое-то очень большое число,
чтобы придать видимость правдоподобия «формуле», ее умножают на какое-то очень большое число,

Слайд 23 в стехиометрических расчетах обычно пренебрегают элементами, составляющими малую часть состава биомассы.
в стехиометрических расчетах обычно пренебрегают элементами, составляющими малую часть состава биомассы.

Слайд 24 в стехиометрическом уравнении все члены можно умножить или разделить на одно
в стехиометрическом уравнении все члены можно умножить или разделить на одно

Слайд 25Удобно принимать такую «молекулярную массу», чтобы в ней оказался только один атом
Удобно принимать такую «молекулярную массу», чтобы в ней оказался только один атом

Слайд 26 дрожжи — CH1,66O0,48N 0,14,
бактерии — CH1,58O0,28N0,19;
«усредненная» биомасса — CH1,92O0,30N0,24;
дрожжи — CH1,66O0,48N 0,14,
бактерии — CH1,58O0,28N0,19;
«усредненная» биомасса — CH1,92O0,30N0,24;

Слайд 27для «усредненной» биомассы используется формула, предложенная Стоутхамером для С-моля:
CH1,8O0,5N0,2
Из-за простоты эту формулу
для «усредненной» биомассы используется формула, предложенная Стоутхамером для С-моля:
CH1,8O0,5N0,2
Из-за простоты эту формулу

Слайд 28Рассчет «молекулярной массы»
С-моля:
М= 1·12 + 1,8·1 + 0,5·16 + 0,2·14 =
Рассчет «молекулярной массы»
С-моля:
М= 1·12 + 1,8·1 + 0,5·16 + 0,2·14 =

Слайд 29 РАСЧЕТ ВЫХОДА БИОМАССЫ НА УГЛЕРОДНЫЙ СУБСТРАТ
наибольший интерес с учетом
РАСЧЕТ ВЫХОДА БИОМАССЫ НА УГЛЕРОДНЫЙ СУБСТРАТ
наибольший интерес с учетом

Слайд 30Эти вещества можно также пересчитать на С-моль (т.е. оставить в молекуле только
Эти вещества можно также пересчитать на С-моль (т.е. оставить в молекуле только

Слайд 31разные субстраты дают различный стехиометрический выход по биомассе.
фактически измеренные данные выхода биомассы
разные субстраты дают различный стехиометрический выход по биомассе.
фактически измеренные данные выхода биомассы

Слайд 32 Чтобы объяснить эти расхождения, ввели понятие «энергетический выход» биомассы.
- в
Чтобы объяснить эти расхождения, ввели понятие «энергетический выход» биомассы.
- в

Слайд 33 Степень восстановленности зависит от числа так называемых «доступных электронов», или «редоксонов».
Степень восстановленности зависит от числа так называемых «доступных электронов», или «редоксонов».

Слайд 34степень восстановленности субстрата CmHnOpNq рассчитывают по формуле:
γs = 4m + n -
степень восстановленности субстрата CmHnOpNq рассчитывают по формуле:
γs = 4m + n -

Слайд 35Оценка биомассы по степени восстановленности (обозначение γх)
Биомасса (CHl,8O0,5N0,2):
γX =4 х 1 +
Оценка биомассы по степени восстановленности (обозначение γх)
Биомасса (CHl,8O0,5N0,2):
γX =4 х 1 +

Слайд 36 В.К. Ерошиным введено понятие энергетический выход биомассы, представляющий собой отношение энергии
В.К. Ерошиным введено понятие энергетический выход биомассы, представляющий собой отношение энергии

Слайд 37метан η = 8,0/4,2 = 1,9 С-моль биомассы/C-моль субстрата;
гексан η = 6,3/4,2
метан η = 8,0/4,2 = 1,9 С-моль биомассы/C-моль субстрата;
гексан η = 6,3/4,2

Слайд 38 В формуле для η выход выражен в С-молях биомассы на С-моль
В формуле для η выход выражен в С-молях биомассы на С-моль

Слайд 39метан YXS = 1,9 х 24,6/16 = 2,92 г биомассы/ г субстрата;

Слайд 40для глюкозы энергетический выход биомассы дает значения чуть ниже стехиометрического (0,78 против
для глюкозы энергетический выход биомассы дает значения чуть ниже стехиометрического (0,78 против

Слайд 41ОПРЕДЕЛЕНИЕ СТЕХИОМЕТРИЧЕСКИХ СООТНОШЕНИЙ В РЕАЛЬНЫХ ПРОЦЕССАХ ФЕРМЕНТАЦИИ
исходные предпосылки «энергетической» теории материального
ОПРЕДЕЛЕНИЕ СТЕХИОМЕТРИЧЕСКИХ СООТНОШЕНИЙ В РЕАЛЬНЫХ ПРОЦЕССАХ ФЕРМЕНТАЦИИ
исходные предпосылки «энергетической» теории материального

Слайд 42общее стехиометрическое соотношение для объединенного процесса:
ʋs S + ʋo2 [02] + ʋN
общее стехиометрическое соотношение для объединенного процесса:
ʋs S + ʋo2 [02] + ʋN
![общее стехиометрическое соотношение для объединенного процесса: ʋs S + ʋo2 [02] +](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/975173/slide-41.jpg)
Слайд 43Необходимо знать, количество израсходованного субстрата (в реальных мерах — граммы, килограммы и

Слайд 44 Разделив эти величины на молекулярные массы соответственно субстрата, биомассы или продукта,
Разделив эти величины на молекулярные массы соответственно субстрата, биомассы или продукта,

Слайд 45 Первый из этих расчетов — приведение всех количеств к одному С-молю
Первый из этих расчетов — приведение всех количеств к одному С-молю

Слайд 46Таким образом сразу находим ʋs и ʋР.
Для определения коэффициентов по другим
Таким образом сразу находим ʋs и ʋР.
Для определения коэффициентов по другим

Слайд 47 При этом индексы в формуле субстрата обозначим как ms, ns, ps,
При этом индексы в формуле субстрата обозначим как ms, ns, ps,

Слайд 48 уравнение баланса по углероду:
ʋSmS = 1 + ʋpmp + ʋCO2
То же
уравнение баланса по углероду:
ʋSmS = 1 + ʋpmp + ʋCO2
То же

Слайд 49Уравнение баланса по кислороду:
ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp
Уравнение баланса по кислороду:
ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp

Слайд 50 Решение системы уравнений позволит найти все стехиометрические коэффициенты для данного процесса
Решение системы уравнений позволит найти все стехиометрические коэффициенты для данного процесса

Слайд 51Переведем все величины в г моли соответствующих веществ
Молекулярная масса (г/моль): сахарозы
Переведем все величины в г моли соответствующих веществ
Молекулярная масса (г/моль): сахарозы

Слайд 52 Пересчитаем все стехиометрические коэффициенты на С-моль биомассы, т.е. разделим обе части
Пересчитаем все стехиометрические коэффициенты на С-моль биомассы, т.е. разделим обе части

Слайд 53Для нахождения оставшихся коэффи-циентов записываем систему уравнений элементного баланса для данного процесса
При
Для нахождения оставшихся коэффи-циентов записываем систему уравнений элементного баланса для данного процесса
При

Слайд 54Баланс по водороду: ʋSnS + ʋNH3 · 3 = 1,8 + ʋpnp
Баланс по водороду: ʋSnS + ʋNH3 · 3 = 1,8 + ʋpnp

Слайд 55Баланс по кислороду: ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp
Баланс по кислороду: ʋSpS + ʋO2 · 2 = 0,5 + ʋpPp

Слайд 56 Баланс по азоту: ʋsqs + ʋNH3 = 0,2 + ʋpqp
0 +
Баланс по азоту: ʋsqs + ʋNH3 = 0,2 + ʋpqp
0 +

 Презентация на тему Кто такие рыбы (1 класс)
Презентация на тему Кто такие рыбы (1 класс)  Первоцеты в природе
Первоцеты в природе Флоэма и ксилема. Их гистологический состав
Флоэма и ксилема. Их гистологический состав Антропогенез и этногенез. Лекция 4
Антропогенез и этногенез. Лекция 4 Пищеварительная система человека
Пищеварительная система человека СТРОЕНИЕ. ФУНКЦИОНИРОВАНИЕ И СВОЙСТВА ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ. ШИРЯКОВА ЭВЕЛИНА. КЛЫПЕНКОВА НАДЕЖДА. ЛЕОНТЬЕВА ДИАНА
СТРОЕНИЕ. ФУНКЦИОНИРОВАНИЕ И СВОЙСТВА ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ. ШИРЯКОВА ЭВЕЛИНА. КЛЫПЕНКОВА НАДЕЖДА. ЛЕОНТЬЕВА ДИАНА Презентация на тему Путешествие в мир Простейших
Презентация на тему Путешествие в мир Простейших  Свойства живого
Свойства живого Строение одноклеточной водоросли хламидомонады
Строение одноклеточной водоросли хламидомонады Экологические группы птиц
Экологические группы птиц Цветок, его строение и значение
Цветок, его строение и значение Моногибридное скрещивание Законы Менделя
Моногибридное скрещивание Законы Менделя Цитология. Строение клетки
Цитология. Строение клетки Хищники и их жертвы
Хищники и их жертвы Помидоры. Выполнила: Лагунова Елизавета 5 Б класс
Помидоры. Выполнила: Лагунова Елизавета 5 Б класс Презентация на тему Клонирование за и против
Презентация на тему Клонирование за и против  Презентация на тему ВОЗДУШНОЕ ПИТАНИЕ РАСТЕНИЙ
Презентация на тему ВОЗДУШНОЕ ПИТАНИЕ РАСТЕНИЙ  Ультраструктура поперечнополосатых скелетных мышечных волокон. Биомеханика мышцы
Ультраструктура поперечнополосатых скелетных мышечных волокон. Биомеханика мышцы Геоботаник и флорист А.А. Дедов, интиолог Е.С. Кучина
Геоботаник и флорист А.А. Дедов, интиолог Е.С. Кучина Насекомые-паразиты
Насекомые-паразиты Презентация на тему ВНЕШНЕЕ СТРОЕНИЕ ЛИСТА
Презентация на тему ВНЕШНЕЕ СТРОЕНИЕ ЛИСТА  Пищеварительная система. Зубы, строение и функция
Пищеварительная система. Зубы, строение и функция История генетики. Основные понятия генетики (урок 19)
История генетики. Основные понятия генетики (урок 19) Анатомия черепа
Анатомия черепа Эмбриология. Эмбриональный гистогенез
Эмбриология. Эмбриональный гистогенез Липиды. Группы липидов. Функции. Витамины. Холестерин
Липиды. Группы липидов. Функции. Витамины. Холестерин Обмен веществ и энергии
Обмен веществ и энергии Грибы. Строение, физиология, методы изучения
Грибы. Строение, физиология, методы изучения