- Микрогаметофит. Созревание, покой и прорастание. Лекция 3

Содержание
- 2. Метаболизм мужского гаметофита На протяжении развития пыльцевого зерна идет интенсивный рост объема и сухой массы вегетативной
- 3. Биосинтетическая активность Синтетическая активность вегетативной клетки существенно изменяется на протяжении развития мужского гаметофита. Скорость синтеза РНК
- 4. Специфичные и обычные 1. Гены «домашнего хозяйства»(housekeeping genes), которые экспрессируются также в соматических клетках растений. 2.
- 5. Концепция 3-х стадий (протеом) 1 стадия – от микроспороцита до молодой микроспоры 2 стадия – поляризованная
- 6. Относительное содержание элементов на разных стадиях гаметофитогенеза Андреюк ДС, Богданов АГ, Матвеева НП, Тукеева МИ, Ермаков
- 7. рН и поглощение кислорода в процессе гаметофитогенеза рН Стадия 1 Стадия 2 Стадия 3 Стадия 3
- 8. Поляризация и деления микроспоры, формирование оболочки Метаболизм, транскриптом и протеом мужского гаметофита Дегидратация пыльцевых зерен Адгезия
- 9. Изменение оводнённости Степень гидратации мужского гаметофита изменяется в ходе его развития. 1) Формирование пыльцевого зерна, когда
- 10. Дегидратация Пыльцевые зёрна подразделяют на частично дегидратированные ( 20-30%). Первая сохраняется дольше, прорастает медленнее.
- 11. Дессиком Это набор генов, белков и метаболитов, обеспечивающих устойчивость клеток к дегидратации Включает регуляторные механизмы и
- 12. Гены, входящие в дессиком Стабилизация мембран и белков с помощью невосстанавливающих сахаров, белков теплового шока и
- 13. Поляризация и деления микроспоры, формирование оболочки Метаболизм, транскриптом и протеом мужского гаметофита Дегидратация пыльцевых зерен Адгезия
- 14. Активация пыльцевого зерна Активация дыхания (скорость процесса зависит от состояния митохондрий) Сдвиг рН в щелочную сторону
- 15. Динамика рН и скорости поглощения кислорода в процессе активации зрелого ПЗ in vitro 10 мин 20
- 16. Два оптических метода оценки Em Ратиометрический быстрый краситель Di-4-ANEPPS: Съемка в двух каналах при возбуждении синим
- 17. H+-АТФаза плазмалеммы Этот фермент – один из важнейших в растительной клетке. Выкачивая протоны, он влияет на
- 18. Всему голова – Са2+ Концентрация кальция возрастает в месте выхода пыльцевой трубки Ингибиторы подавляют прорастание. Iwano
- 19. Вблизи апертуры, через которую выйдет пыльцевая трубка, собираются везикулы Гольджи, митохондрии и короткие цистерны ЭПР, подготавливая
- 20. Поляризация и деления микроспоры, формирование оболочки Метаболизм, транскриптом и протеом мужского гаметофита Дегидратация пыльцевых зерен Адгезия
- 21. Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение Клеточная стенка – механическая составляющая
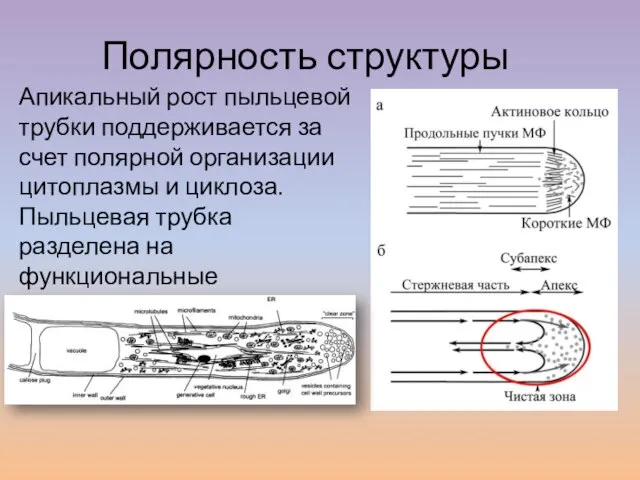
- 22. Апикальный рост пыльцевой трубки поддерживается за счет полярной организации цитоплазмы и циклоза. Пыльцевая трубка разделена на
- 23. Функции циклоза 1. Транспорт мужского гаметного модуля (MGU). 2. Транспорт в апекс сигнальных молекул и ферментов
- 24. Актиновые цитоскелет играет ключевую роль в поддержании полярного роста Продольные актиновые тяжи обеспечивают ток цитоплазмы и
- 25. Микротрубочки Играют ключевую роль в доставке мужского гаметного модуля У цветковых не участвуют в циклозе У
- 26. Везикулы аппарата Гольджи, содержащие материал для строительства апикальной мембраны и стенки, путешествуют в апекс по актиновым
- 27. Клеточная стенка: градиент жёсткости Движущей силой для роста является тургорное давление. Однако, оно не является направленной
- 28. Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза По мере удаления от кончика они деэтерифицируются
- 29. Каллоза Breygina et al., 2012 Xie et al., 2012 Отсутствует в кончике Появляется на расстоянии 10-30
- 30. Жесткий компонент, но его не очень много Отложение также неравномерное: в кончике отсутствует, далее постепенно накапливается
- 31. Фосфолипидный сигналлинг Гидратация и дегидратация Цитоскелет Экзо- и эндоцитоз ГТФазы Трансмембранный ионный транспорт Универсальные регуляторные механизмы
- 32. Регуляторные механизмы: фосфолипиды Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения актинового кольца. Фосфатидилинозитолдифосфат
- 33. Его узкая локализация обеспечивается белком phosphotidylinositol transfer protein (PITP) и фосфолипазой C (PLC), которая режет PIP2
- 34. ROP ГТФазы и рецепторные киназы В клетках растений основную роль играют мономерные (малые) G белки (20-30
- 35. Малые ГТФазы, обозначаемые как Rac/Rop или ROP (Rho of plants) выполняют функции клеточных хабов – центральных
- 36. RAC/ROPs и начальные стадии прорастания Активация пыльцы как in vitro, так и in vivo сопровождается активацией
- 37. Регуляция по принципу обратной связи Рецепторная киназа RLK активирует фактор обмена нуклеотидов GEF, ROP переводится в
- 38. ROP1 в трубке В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована преимущественно в апикальной
- 39. Эффекторные белки Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и контролирует накопление везикул, несущих
- 40. Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА, которая включает в себя
- 41. Основные закономерности прорастания и роста пыльцевого зерна: ионный статус Feijó et al. 1992; Pertl et al.
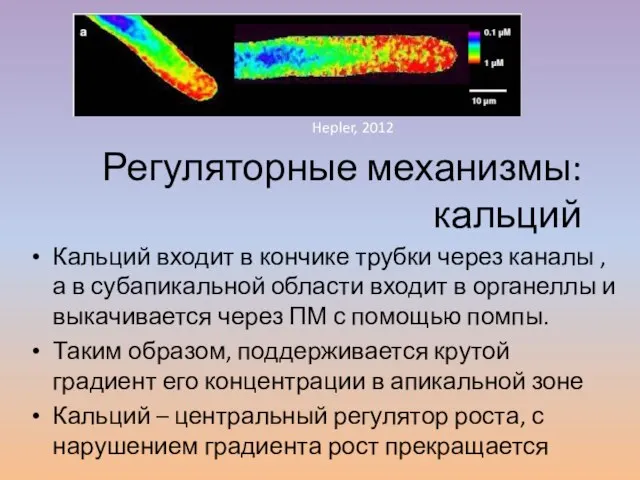
- 42. Регуляторные механизмы: кальций Hepler, 2012 Кальций входит в кончике трубки через каналы , а в субапикальной
- 43. Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает направление роста Кальций регулирует
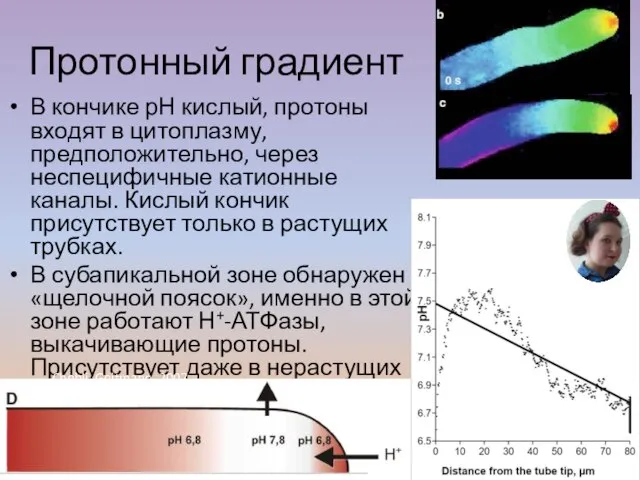
- 44. В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные каналы. Кислый кончик присутствует
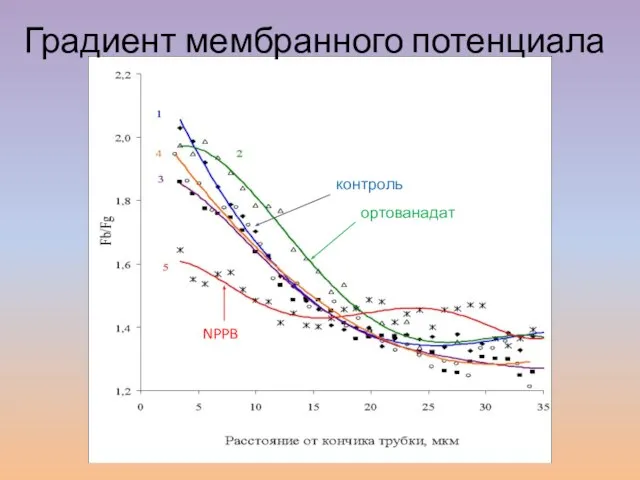
- 45. контроль ортованадат фузикокцин Брейгина и др., 2010 Градиент мембранного потенциала контроль ортованадат NPPB Брейгина и др.,
- 46. АФК ионный транспорт ? АФК в прогамной фазе оплодотворения, по-видимому, могут передавать сигнал и регулировать рост.
- 47. Активные формы кислорода (АФК) - высокореакционные, метастабильными агенты (молекулы, ионы, радикалы), обладающие избыточной энергией по отношению
- 48. АФК и пероксидазы на рыльце Пероксидазы и АФК накапливаются при подготовке к опылению.
- 49. АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму Breygina et al., 2016.
- 50. АФК как сигнал: H2O2 активирует ток K+ Сравнение средней плотности тока при (в % от контроля).
- 51. АФК как сигнал: H2O2 убирает в трубках «щелочной поясок» В растущих трубках лилии вслед за массированным
- 52. Два оптических метода оценки Em Ратиометрический быстрый краситель Di-4-ANEPPS: Съемка в двух каналах при возбуждении синим
- 53. Количественная флуоресцентная микроскопия: динамика эффекта Проточная цитометрия с красителем DiBAC4(3): исследование большой популяции протопластов АФК как
- 54. АФК как сигнал: H2O2 вызывает гиперполяризацию После 10 минут инкубации происходит достоверное смещение популяции в область
- 55. АФК как сигнал: H2O2 вызывает гиперполяризацию Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: интактные растущие трубки лилии
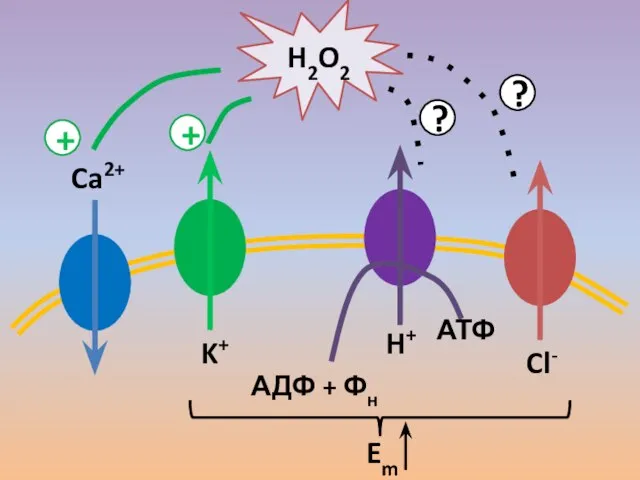
- 56. Ca2+ K+ АДФ + Фн АТФ H+ Cl- H2O2 + + ? ? Em
- 57. В трубке ион-транспортные системы расположены полярно Они формируют в цитоплазме динамичные градиенты концентраций, а на мембране
- 58. Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не было в контроле. Основные
- 59. Цветковые VS Хвойные Изученный модельный объект Быстрый рост Униполярный рост Обращенный фонтан Крутой градиент Са2+ Крутой
- 60. Градиент мембранного потенциала табак ель
- 61. Биполярное прорастание у ели Breygina et al., 2019 (в соавторстве с Полевовой С.В.)
- 62. Две трубки, а МГМ один. Куда же он пойдет? Он тоже сомневается… b a c d
- 63. Биполярное прорастание: динамика 2 часа 6 часов 9 часов Сколько разрывов – столько и трубок Breygina
- 65. Скачать презентацию
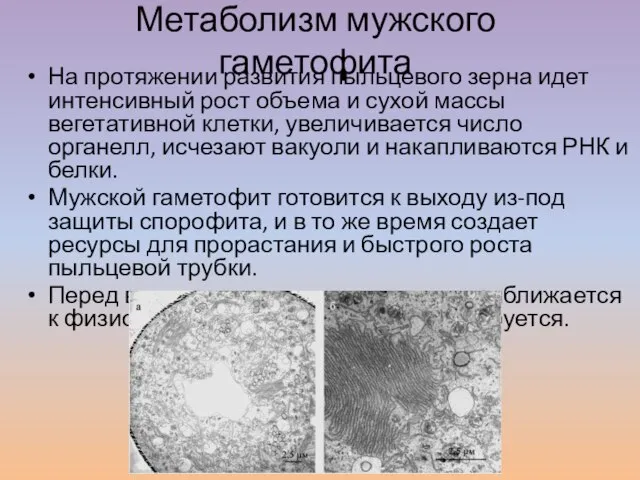
Слайд 2Метаболизм мужского гаметофита
На протяжении развития пыльцевого зерна идет интенсивный рост объема и
Метаболизм мужского гаметофита
На протяжении развития пыльцевого зерна идет интенсивный рост объема и
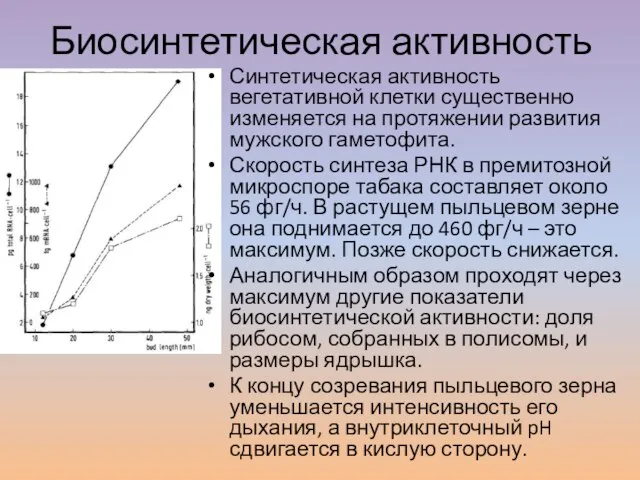
Слайд 3Биосинтетическая активность
Синтетическая активность вегетативной клетки существенно изменяется на протяжении развития мужского гаметофита.
Биосинтетическая активность
Синтетическая активность вегетативной клетки существенно изменяется на протяжении развития мужского гаметофита.
Слайд 4Специфичные и обычные
1. Гены «домашнего хозяйства»(housekeeping genes), которые экспрессируются также в соматических
Специфичные и обычные
1. Гены «домашнего хозяйства»(housekeeping genes), которые экспрессируются также в соматических

Слайд 5Концепция 3-х стадий (протеом)
1 стадия – от микроспороцита до молодой микроспоры
2 стадия
Концепция 3-х стадий (протеом)
1 стадия – от микроспороцита до молодой микроспоры
2 стадия
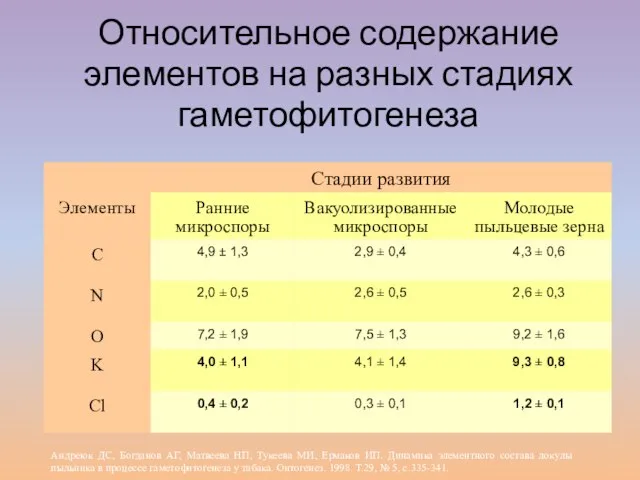
Слайд 6
Относительное содержание элементов на разных стадиях гаметофитогенеза
Андреюк ДС, Богданов АГ, Матвеева НП,
Относительное содержание элементов на разных стадиях гаметофитогенеза
Андреюк ДС, Богданов АГ, Матвеева НП,
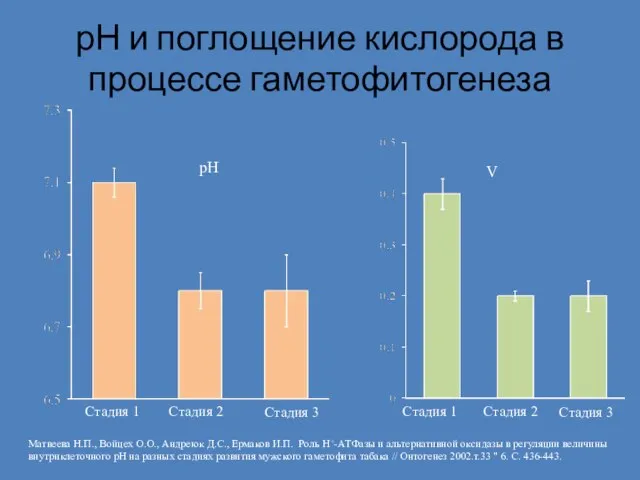
Слайд 7рН и поглощение кислорода в процессе гаметофитогенеза
рН
Стадия 1
Стадия 2
Стадия 3
Стадия 3
Стадия 2
Стадия
рН и поглощение кислорода в процессе гаметофитогенеза
рН
Стадия 1
Стадия 2
Стадия 3
Стадия 3
Стадия 2
Стадия
Слайд 8Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых
Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых

Слайд 9Изменение оводнённости
Степень гидратации мужского гаметофита изменяется в ходе его развития.
1) Формирование
Изменение оводнённости
Степень гидратации мужского гаметофита изменяется в ходе его развития.
1) Формирование

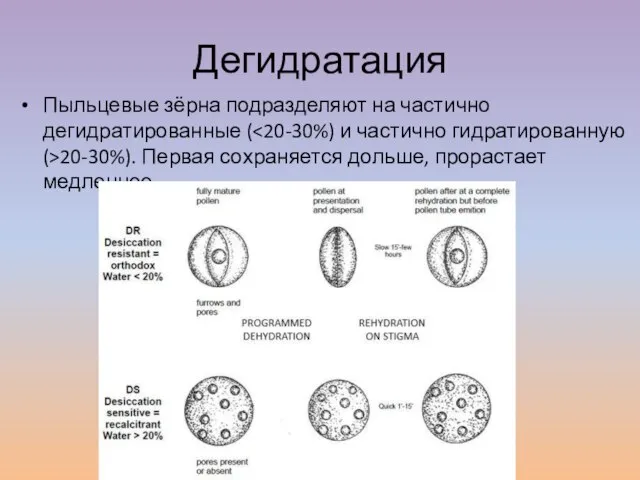
Слайд 10Дегидратация
Пыльцевые зёрна подразделяют на частично дегидратированные (<20-30%) и частично гидратированную (>20-30%). Первая
Дегидратация
Пыльцевые зёрна подразделяют на частично дегидратированные (<20-30%) и частично гидратированную (>20-30%). Первая
Слайд 11Дессиком
Это набор генов, белков и метаболитов, обеспечивающих устойчивость клеток к дегидратации
Включает регуляторные
Дессиком
Это набор генов, белков и метаболитов, обеспечивающих устойчивость клеток к дегидратации
Включает регуляторные
Слайд 12Гены, входящие в дессиком
Стабилизация мембран и белков с помощью невосстанавливающих сахаров, белков
Гены, входящие в дессиком
Стабилизация мембран и белков с помощью невосстанавливающих сахаров, белков

Слайд 13Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых
Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых

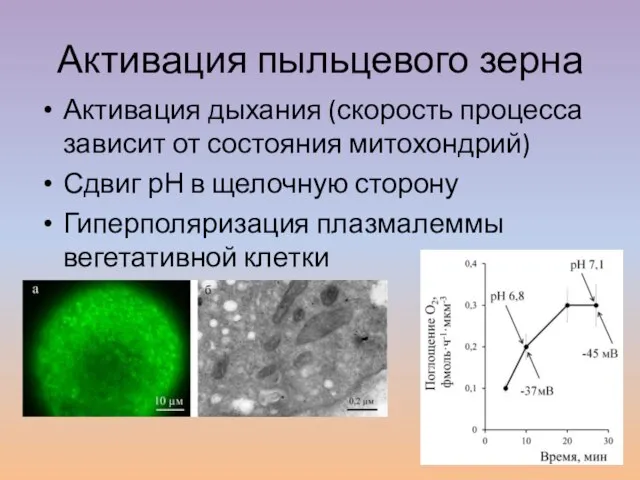
Слайд 14Активация пыльцевого зерна
Активация дыхания (скорость процесса зависит от состояния митохондрий)
Сдвиг рН в
Активация пыльцевого зерна
Активация дыхания (скорость процесса зависит от состояния митохондрий)
Сдвиг рН в
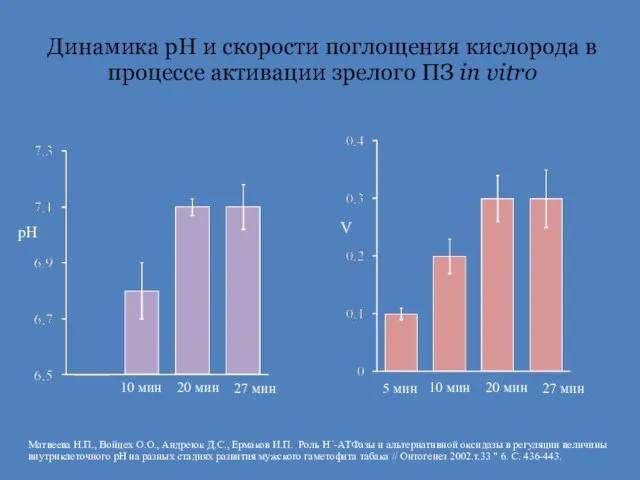
Слайд 15Динамика рН и скорости поглощения кислорода в процессе активации зрелого ПЗ in
Динамика рН и скорости поглощения кислорода в процессе активации зрелого ПЗ in
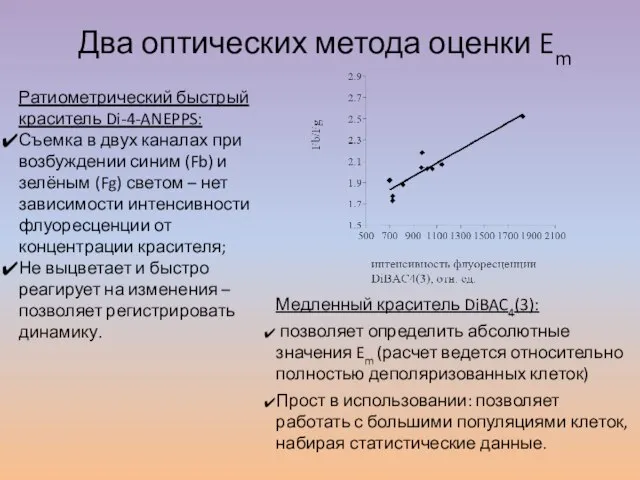
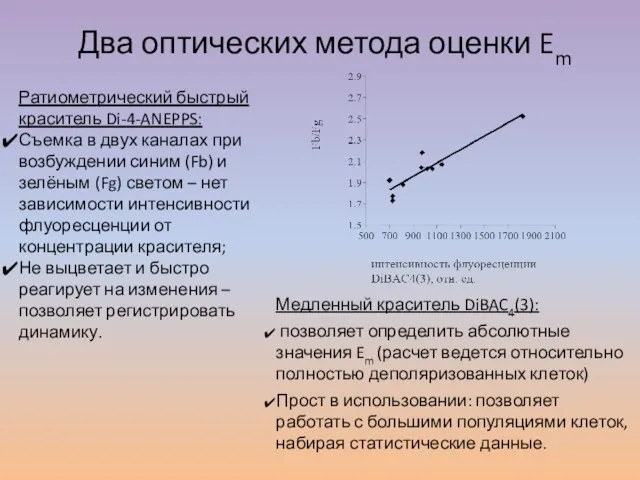
Слайд 16Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при
Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при
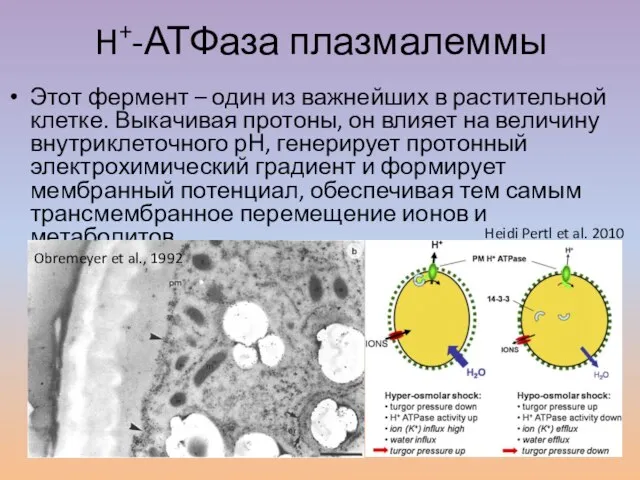
Слайд 17H+-АТФаза плазмалеммы
Этот фермент – один из важнейших в растительной клетке. Выкачивая протоны,
H+-АТФаза плазмалеммы
Этот фермент – один из важнейших в растительной клетке. Выкачивая протоны,
Слайд 18Всему голова – Са2+
Концентрация кальция возрастает в месте выхода пыльцевой трубки
Ингибиторы подавляют
Всему голова – Са2+
Концентрация кальция возрастает в месте выхода пыльцевой трубки
Ингибиторы подавляют

Слайд 19Вблизи апертуры, через которую выйдет пыльцевая трубка, собираются везикулы Гольджи, митохондрии и
Вблизи апертуры, через которую выйдет пыльцевая трубка, собираются везикулы Гольджи, митохондрии и
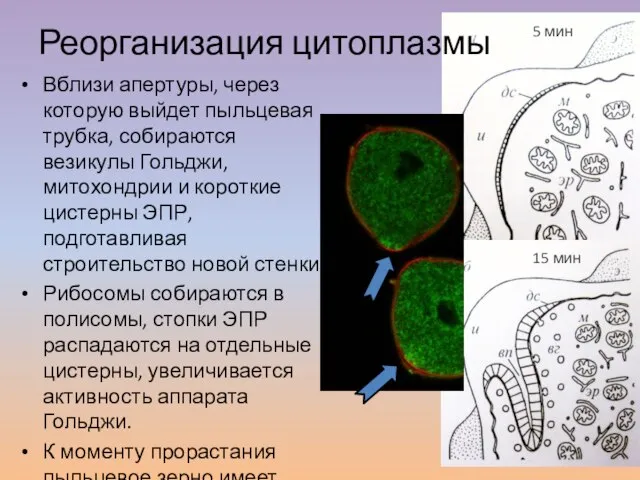
Слайд 20Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых
Поляризация и деления микроспоры, формирование оболочки
Метаболизм, транскриптом и протеом мужского гаметофита
Дегидратация пыльцевых

Слайд 21Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение
Клеточная стенка
Везикулярный транспорт и движение органелл по цитоскелету: доставка стройматериалов и энергообеспечение
Клеточная стенка
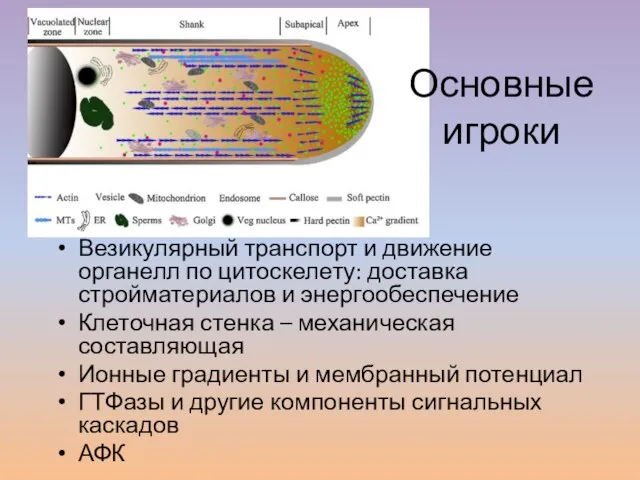
Слайд 22Апикальный рост пыльцевой трубки поддерживается за счет полярной организации цитоплазмы и циклоза.
Апикальный рост пыльцевой трубки поддерживается за счет полярной организации цитоплазмы и циклоза.
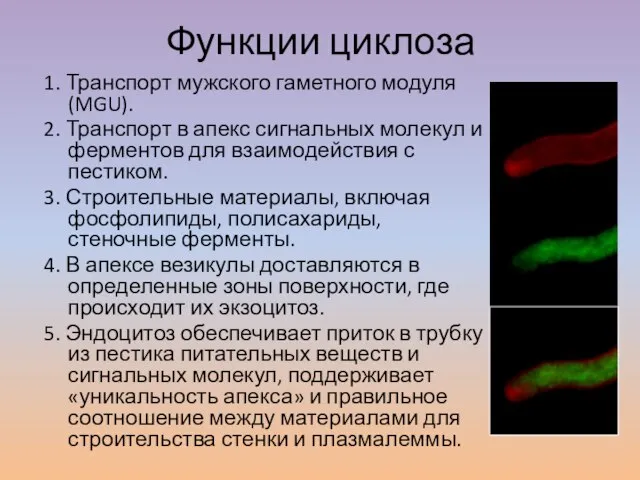
Слайд 23Функции циклоза
1. Транспорт мужского гаметного модуля (MGU).
2. Транспорт в апекс сигнальных молекул
Функции циклоза
1. Транспорт мужского гаметного модуля (MGU).
2. Транспорт в апекс сигнальных молекул
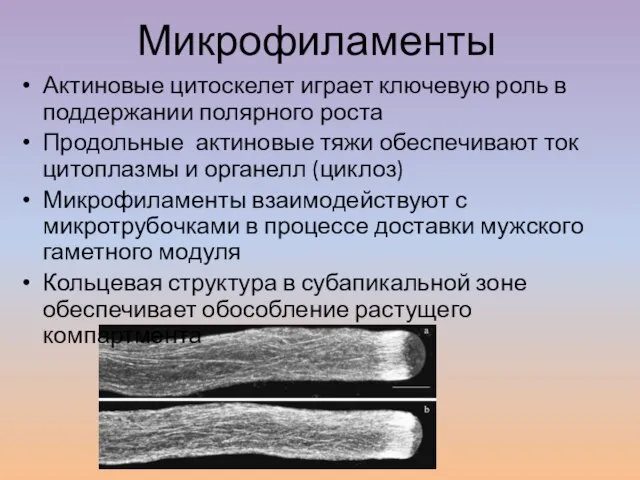
Слайд 24Актиновые цитоскелет играет ключевую роль в поддержании полярного роста
Продольные актиновые тяжи обеспечивают
Актиновые цитоскелет играет ключевую роль в поддержании полярного роста
Продольные актиновые тяжи обеспечивают
Слайд 25Микротрубочки
Играют ключевую роль в доставке мужского гаметного модуля
У цветковых не участвуют в
Микротрубочки
Играют ключевую роль в доставке мужского гаметного модуля
У цветковых не участвуют в

Слайд 26Везикулы аппарата Гольджи, содержащие материал для строительства апикальной мембраны и стенки, путешествуют
Везикулы аппарата Гольджи, содержащие материал для строительства апикальной мембраны и стенки, путешествуют

Слайд 27Клеточная стенка: градиент жёсткости
Движущей силой для роста является тургорное давление. Однако, оно
Клеточная стенка: градиент жёсткости
Движущей силой для роста является тургорное давление. Однако, оно

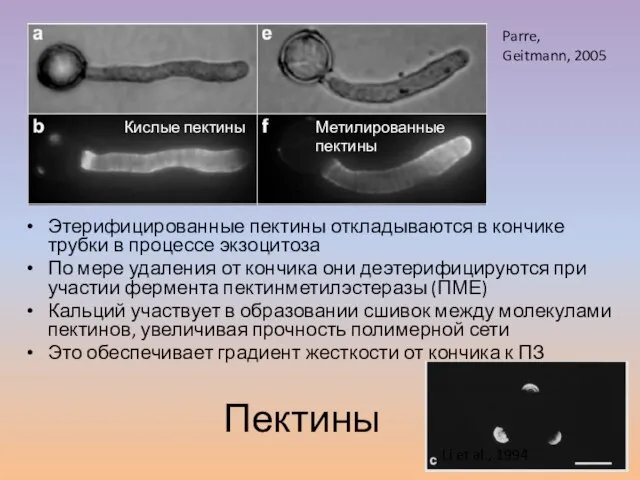
Слайд 28Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза
По мере удаления от
Этерифицированные пектины откладываются в кончике трубки в процессе экзоцитоза
По мере удаления от
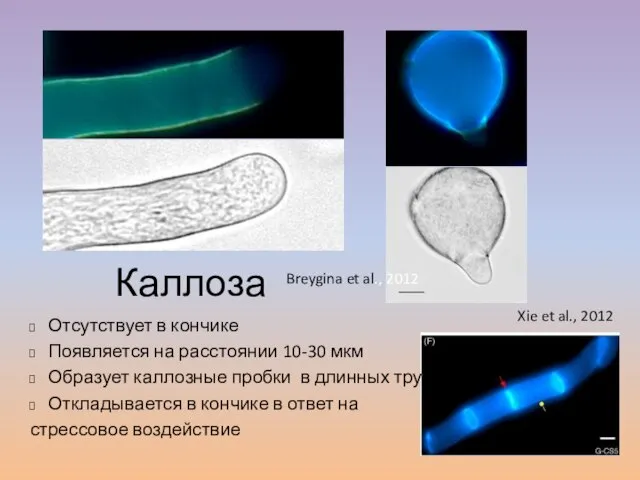
Слайд 29Каллоза
Breygina et al., 2012
Xie et al., 2012
Отсутствует в кончике
Появляется на расстоянии 10-30
Каллоза
Breygina et al., 2012
Xie et al., 2012
Отсутствует в кончике
Появляется на расстоянии 10-30
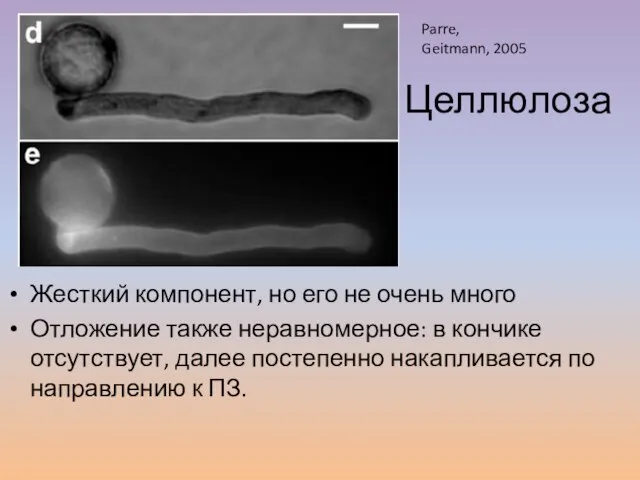
Слайд 30Жесткий компонент, но его не очень много
Отложение также неравномерное: в кончике отсутствует,
Жесткий компонент, но его не очень много
Отложение также неравномерное: в кончике отсутствует,
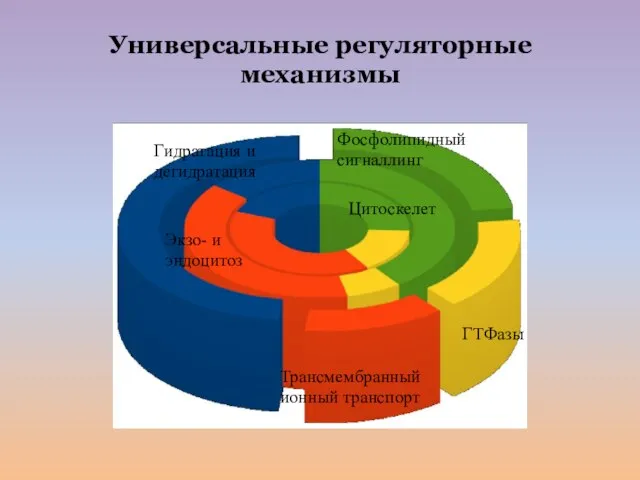
Слайд 31Фосфолипидный сигналлинг
Гидратация и дегидратация
Цитоскелет
Экзо- и эндоцитоз
ГТФазы
Трансмембранный ионный транспорт
Универсальные регуляторные механизмы
Фосфолипидный сигналлинг
Гидратация и дегидратация
Цитоскелет
Экзо- и эндоцитоз
ГТФазы
Трансмембранный ионный транспорт
Универсальные регуляторные механизмы
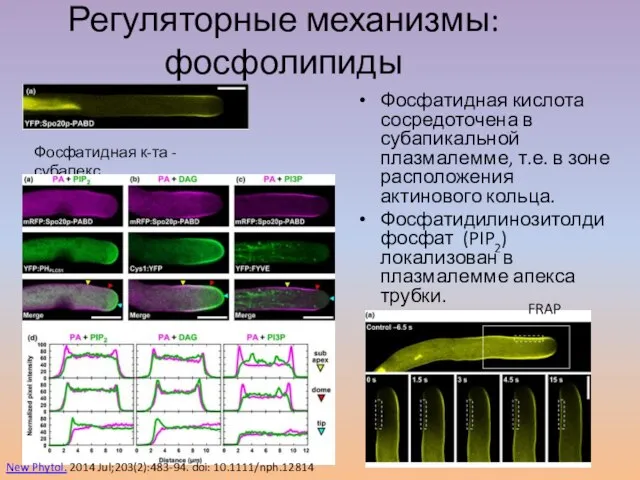
Слайд 32Регуляторные механизмы: фосфолипиды
Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения
Регуляторные механизмы: фосфолипиды
Фосфатидная кислота сосредоточена в субапикальной плазмалемме, т.е. в зоне расположения
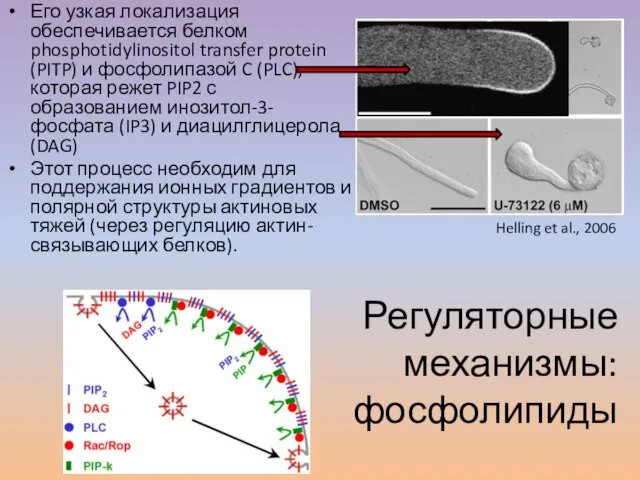
Слайд 33Его узкая локализация обеспечивается белком phosphotidylinositol transfer protein (PITP) и фосфолипазой C
Его узкая локализация обеспечивается белком phosphotidylinositol transfer protein (PITP) и фосфолипазой C
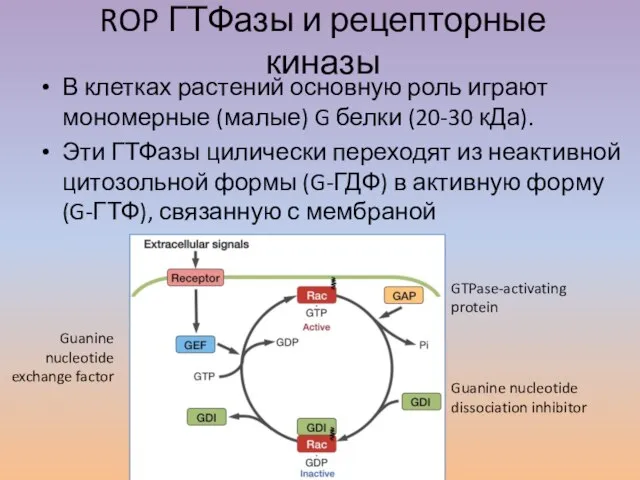
Слайд 34ROP ГТФазы и рецепторные киназы
В клетках растений основную роль играют мономерные (малые)
ROP ГТФазы и рецепторные киназы
В клетках растений основную роль играют мономерные (малые)
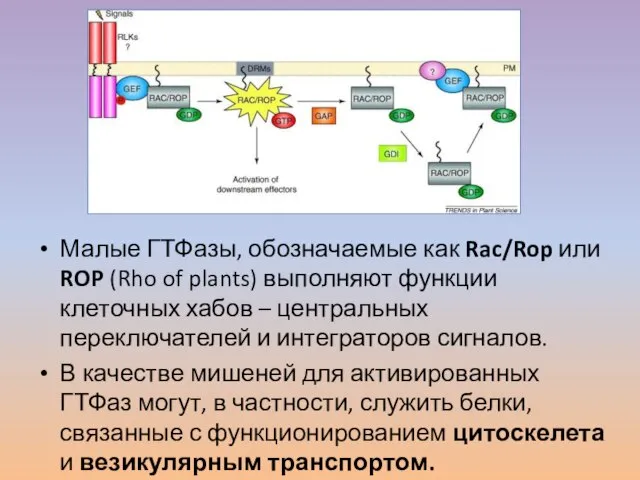
Слайд 35Малые ГТФазы, обозначаемые как Rac/Rop или ROP (Rho of plants) выполняют функции
Малые ГТФазы, обозначаемые как Rac/Rop или ROP (Rho of plants) выполняют функции
Слайд 36RAC/ROPs и начальные стадии прорастания
Активация пыльцы как in vitro, так и in
RAC/ROPs и начальные стадии прорастания
Активация пыльцы как in vitro, так и in

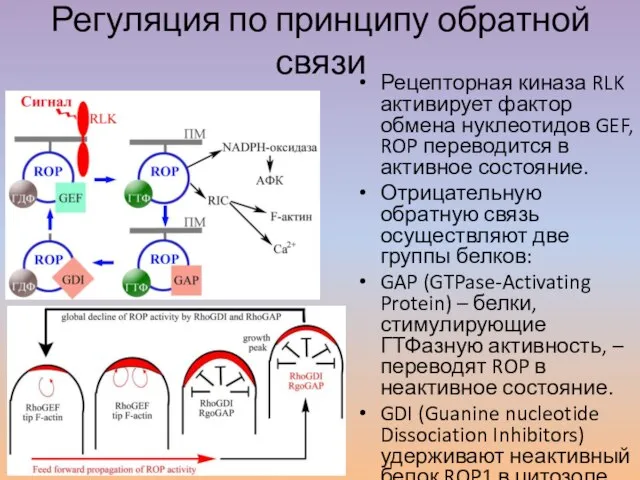
Слайд 37Регуляция по принципу обратной связи
Рецепторная киназа RLK активирует фактор обмена нуклеотидов GEF,
Регуляция по принципу обратной связи
Рецепторная киназа RLK активирует фактор обмена нуклеотидов GEF,
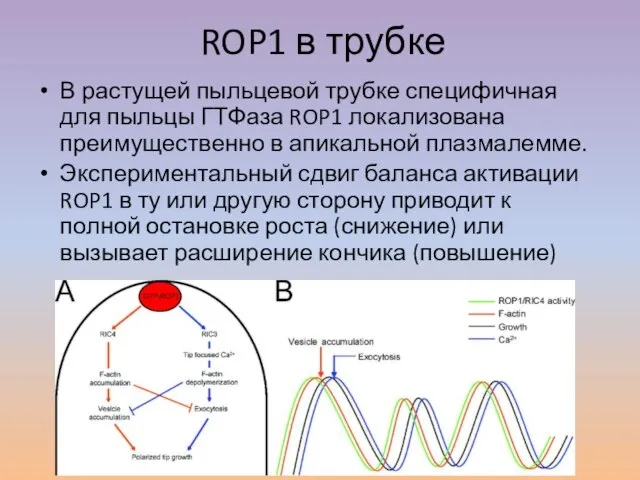
Слайд 38ROP1 в трубке
В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована
ROP1 в трубке
В растущей пыльцевой трубке специфичная для пыльцы ГТФаза ROP1 локализована
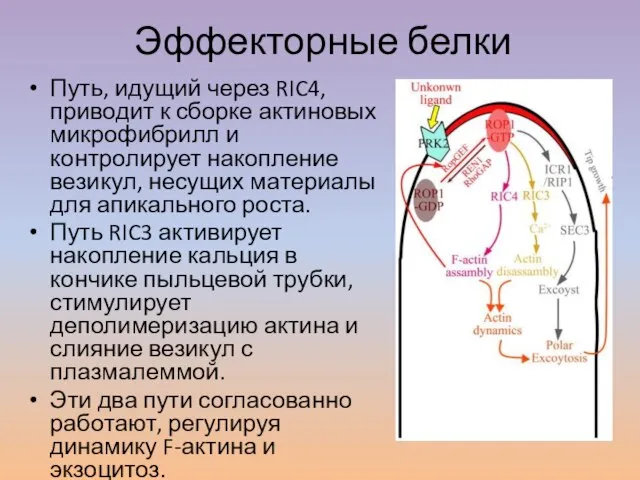
Слайд 39Эффекторные белки
Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и контролирует
Эффекторные белки
Путь, идущий через RIC4, приводит к сборке актиновых микрофибрилл и контролирует
Слайд 40Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА,
Таким образом, пыльцевая трубка – прекрасная модель для изучения ИОННОЙ РЕГУЛЯЦИИ РОСТА,

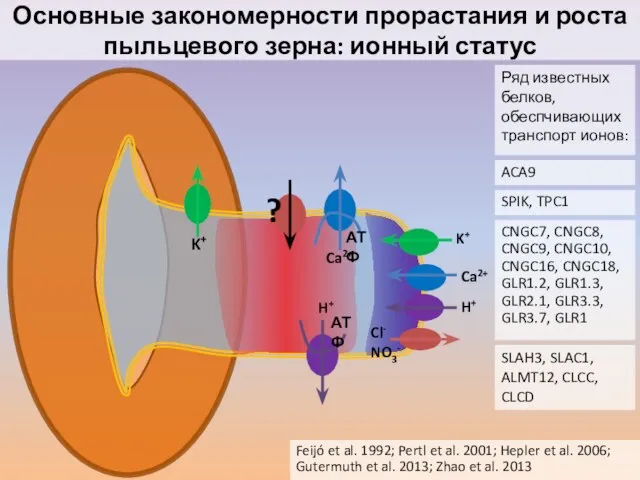
Слайд 41Основные закономерности прорастания и роста пыльцевого зерна: ионный статус
Feijó et al. 1992;
Основные закономерности прорастания и роста пыльцевого зерна: ионный статус
Feijó et al. 1992;
Слайд 42Регуляторные механизмы: кальций
Hepler, 2012
Кальций входит в кончике трубки через каналы , а
Регуляторные механизмы: кальций
Hepler, 2012
Кальций входит в кончике трубки через каналы , а
Слайд 43Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает
Кальциевый градиент определяет место слияния везикул с ПМ и, таким образом, задает

Слайд 44В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные
В кончике рН кислый, протоны входят в цитоплазму, предположительно, через неспецифичные катионные
Слайд 45контроль
ортованадат
фузикокцин
Брейгина и др., 2010
Градиент мембранного потенциала
контроль
ортованадат
NPPB
Брейгина и др., 2010
контроль
ортованадат
фузикокцин
Брейгина и др., 2010
Градиент мембранного потенциала
контроль
ортованадат
NPPB
Брейгина и др., 2010
Слайд 46АФК ионный транспорт ?
АФК в прогамной фазе оплодотворения, по-видимому, могут передавать сигнал
АФК ионный транспорт ?
АФК в прогамной фазе оплодотворения, по-видимому, могут передавать сигнал
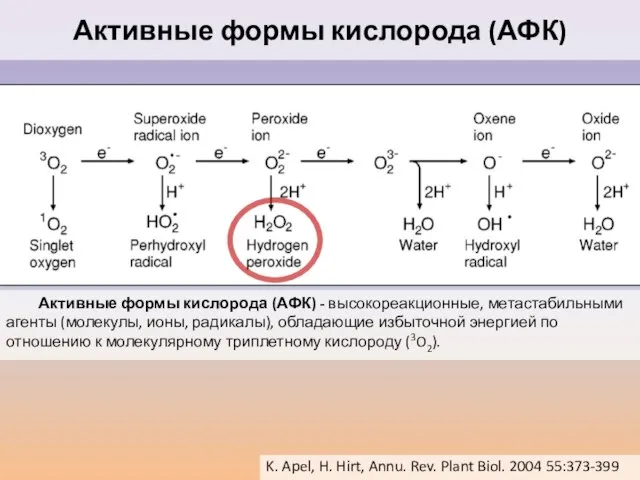
Слайд 47 Активные формы кислорода (АФК) - высокореакционные, метастабильными агенты (молекулы, ионы, радикалы), обладающие
Активные формы кислорода (АФК) - высокореакционные, метастабильными агенты (молекулы, ионы, радикалы), обладающие
Слайд 48АФК и пероксидазы на рыльце
Пероксидазы и АФК накапливаются при подготовке к
АФК и пероксидазы на рыльце
Пероксидазы и АФК накапливаются при подготовке к

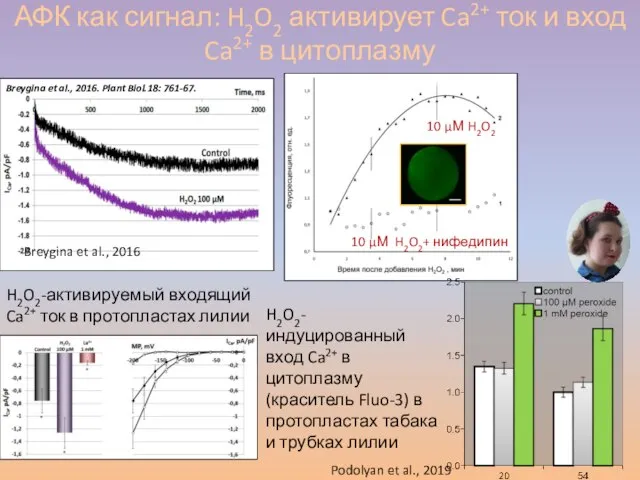
Слайд 49АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму
АФК как сигнал: H2O2 активирует Ca2+ ток и вход Ca2+ в цитоплазму
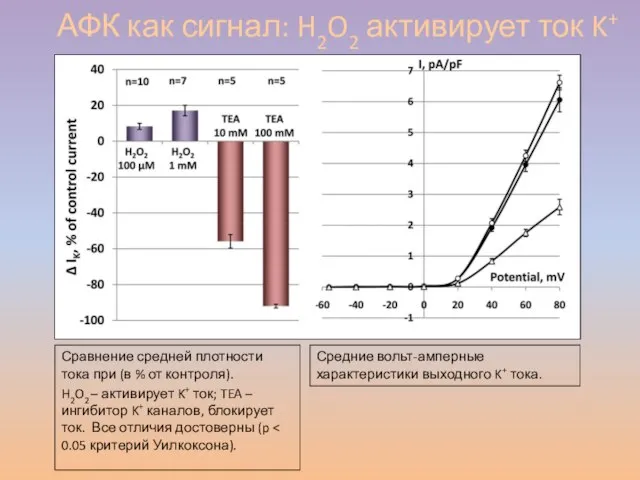
Слайд 50АФК как сигнал: H2O2 активирует ток K+
Сравнение средней плотности тока при (в
АФК как сигнал: H2O2 активирует ток K+
Сравнение средней плотности тока при (в
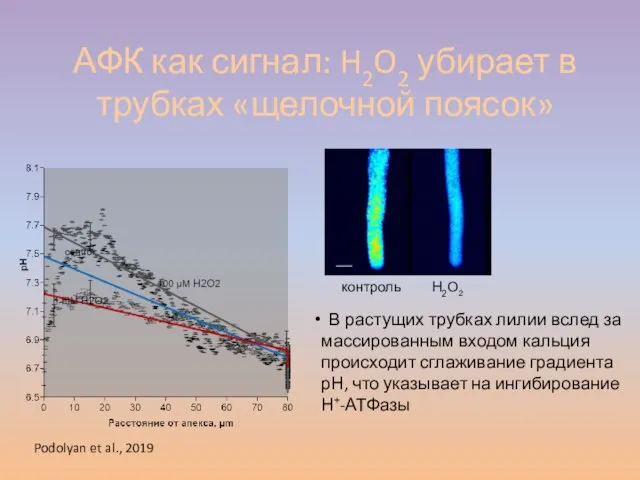
Слайд 51АФК как сигнал: H2O2 убирает в трубках «щелочной поясок»
В растущих трубках
АФК как сигнал: H2O2 убирает в трубках «щелочной поясок»
В растущих трубках
Слайд 52Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при
Два оптических метода оценки Em
Ратиометрический быстрый краситель Di-4-ANEPPS:
Съемка в двух каналах при
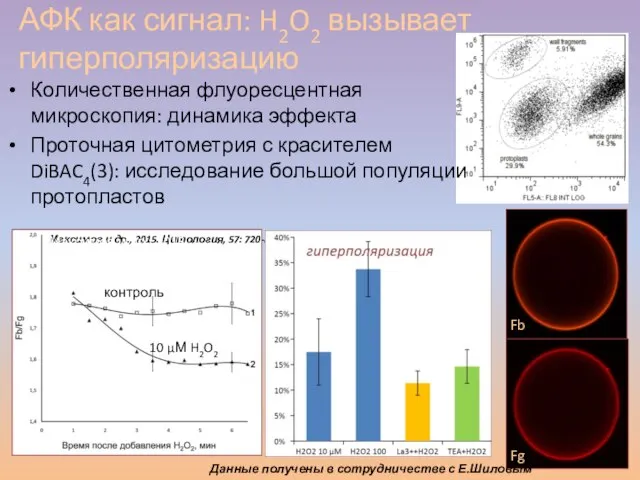
Слайд 53Количественная флуоресцентная микроскопия: динамика эффекта
Проточная цитометрия с красителем DiBAC4(3): исследование большой
Количественная флуоресцентная микроскопия: динамика эффекта
Проточная цитометрия с красителем DiBAC4(3): исследование большой
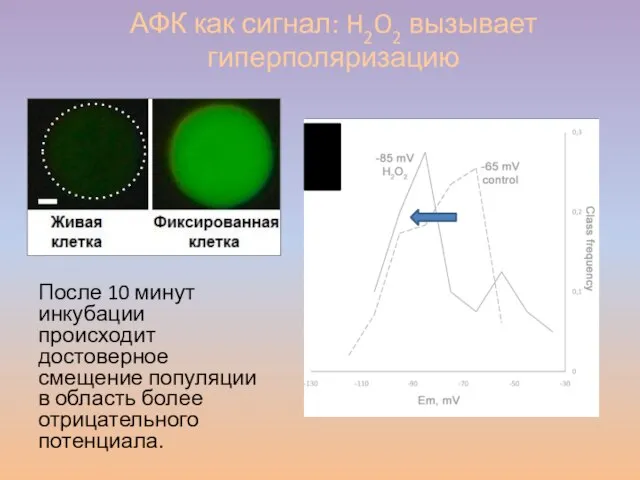
Слайд 54АФК как сигнал: H2O2 вызывает гиперполяризацию
После 10 минут инкубации происходит достоверное смещение
АФК как сигнал: H2O2 вызывает гиперполяризацию
После 10 минут инкубации происходит достоверное смещение
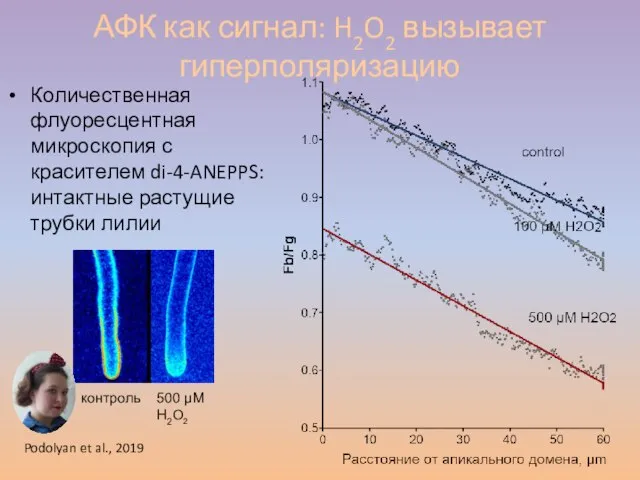
Слайд 55АФК как сигнал: H2O2 вызывает гиперполяризацию
Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: интактные
АФК как сигнал: H2O2 вызывает гиперполяризацию
Количественная флуоресцентная микроскопия с красителем di-4-ANEPPS: интактные
Слайд 56Ca2+
K+
АДФ + Фн
АТФ
H+
Cl-
H2O2
+
+
?
?
Em
Ca2+
K+
АДФ + Фн
АТФ
H+
Cl-
H2O2
+
+
?
?
Em
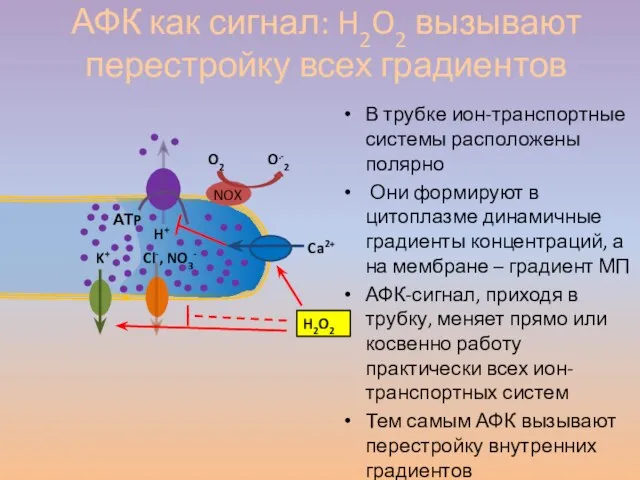
Слайд 57В трубке ион-транспортные системы расположены полярно
Они формируют в цитоплазме динамичные градиенты
В трубке ион-транспортные системы расположены полярно
Они формируют в цитоплазме динамичные градиенты
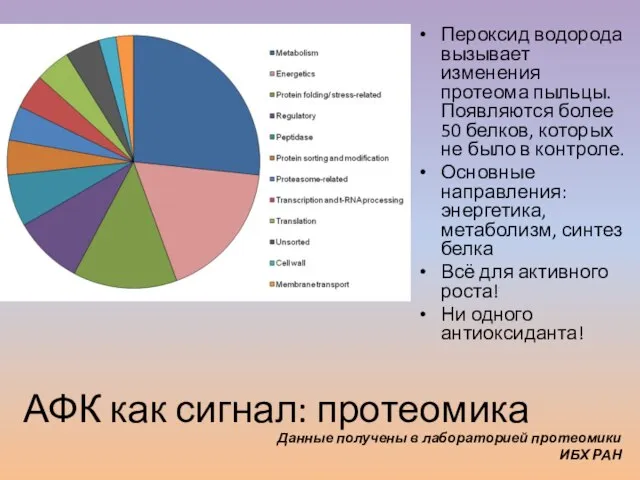
Слайд 58Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не
Пероксид водорода вызывает изменения протеома пыльцы. Появляются более 50 белков, которых не
Слайд 59Цветковые VS Хвойные
Изученный модельный объект
Быстрый рост
Униполярный рост
Обращенный фонтан
Крутой градиент Са2+
Крутой градиент МП
Зависимость
Цветковые VS Хвойные
Изученный модельный объект
Быстрый рост
Униполярный рост
Обращенный фонтан
Крутой градиент Са2+
Крутой градиент МП
Зависимость

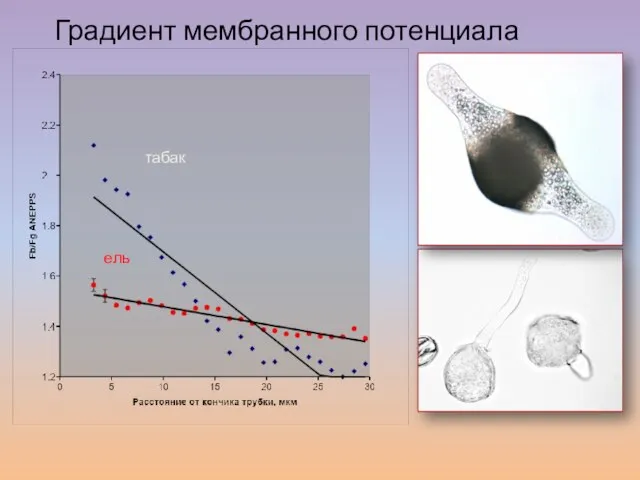
Слайд 60Градиент мембранного потенциала
табак
ель
Градиент мембранного потенциала
табак
ель
Слайд 61Биполярное прорастание у ели
Breygina et al., 2019
(в соавторстве с Полевовой С.В.)
Биполярное прорастание у ели
Breygina et al., 2019
(в соавторстве с Полевовой С.В.)
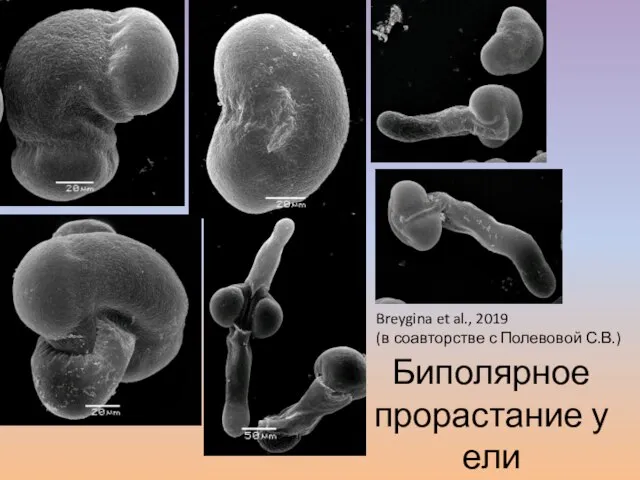
Слайд 62Две трубки, а МГМ один. Куда же он пойдет?
Он тоже сомневается…
b
a
c
d
Биполярное
Две трубки, а МГМ один. Куда же он пойдет?
Он тоже сомневается…
b
a
c
d
Биполярное
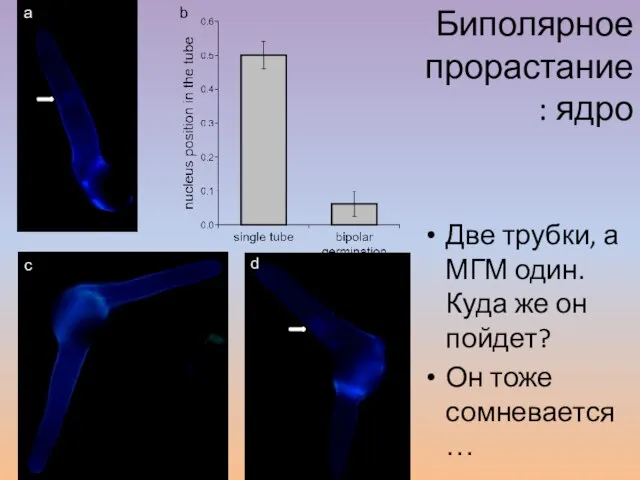
Слайд 63Биполярное прорастание: динамика
2 часа
6 часов
9 часов
Сколько разрывов – столько и трубок
Breygina et
Биполярное прорастание: динамика
2 часа
6 часов
9 часов
Сколько разрывов – столько и трубок
Breygina et

 Вегетативная нервная система
Вегетативная нервная система Центральная нервная система. Brainstem
Центральная нервная система. Brainstem Многообразие и экология грибов
Многообразие и экология грибов Прокариоты, грибы, растения
Прокариоты, грибы, растения Теорія старіння
Теорія старіння Нервная система человека
Нервная система человека Антропология
Антропология Презентация по биологии на тему _Питание и пищевар
Презентация по биологии на тему _Питание и пищевар Красная книга Самарской области
Красная книга Самарской области Лесная викторина
Лесная викторина Микрозелень. Огород на подоконнике
Микрозелень. Огород на подоконнике Гусеницы. Туловище
Гусеницы. Туловище Индивидуальное развитие. Рост и развитие животных
Индивидуальное развитие. Рост и развитие животных Пептидные биорегуляторы - доказанный эффект увеличения периода эффективности человека
Пептидные биорегуляторы - доказанный эффект увеличения периода эффективности человека Обмен веществ и энергии
Обмен веществ и энергии Изучение условий выращивания арбузов на дачном участке в условиях п.г.т. Пржевальское Смоленской области
Изучение условий выращивания арбузов на дачном участке в условиях п.г.т. Пржевальское Смоленской области Витамины. Роль витаминов в организме человека
Витамины. Роль витаминов в организме человека Клеточные культуры в получении вторичных метаболитов
Клеточные культуры в получении вторичных метаболитов История и перспективы развития микробиологии. Лекция 1
История и перспективы развития микробиологии. Лекция 1 Ткани растений. Физиология растения. Агротехнические приемы
Ткани растений. Физиология растения. Агротехнические приемы Пищевая специализация моллюска вида улитка садовая Helix aspersa
Пищевая специализация моллюска вида улитка садовая Helix aspersa Значение дыхания. Органы дыхания. 8 класс
Значение дыхания. Органы дыхания. 8 класс Тестирование по теме Строение клетки
Тестирование по теме Строение клетки Некультивируемое состояние микроорганизмов
Некультивируемое состояние микроорганизмов Гигиена кормов и кормления животных
Гигиена кормов и кормления животных Координация и регуляция. Структура и функции палочек и колбочек, волосковых клеток
Координация и регуляция. Структура и функции палочек и колбочек, волосковых клеток 9-13бесполое размножение
9-13бесполое размножение Властивості ґрунту, догляд за ґрунтом
Властивості ґрунту, догляд за ґрунтом