- Промежуточный мозг

Содержание
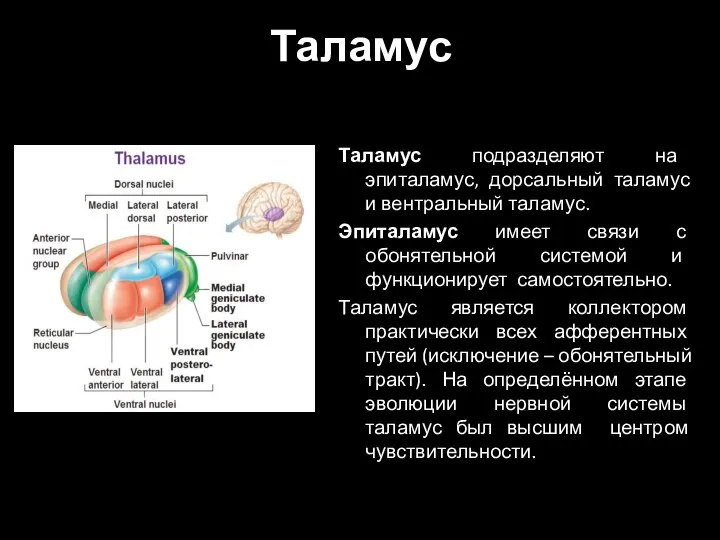
- 2. Таламус Таламус подразделяют на эпиталамус, дорсальный таламус и вентральный таламус. Эпиталамус имеет связи с обонятельной системой
- 3. С развитием коры головного мозга, он утратил эту функцию. Однако, остался высшим центром болевой чувствительности. Принимает
- 4. Основные ядра таламуса Специфические ядра переключающие ассоциативные моторные Неспецифические ядра
- 5. Специфические ядра таламуса ПЕРЕКЛЮЧАЮЩИЕ АССОЦИАТИВНЫЕ МОТОРНЫЕ Вентробазальный Медиодорсальное Переднецентральное комплекс ядро и Вентролатеральное (тактильная, проприоцеп- (Проекция
- 6. 1. Специфические ядра (релейные) а) получают афференты от всех трактов (исключая обонятельный); б) имеют прямой выход
- 7. 2. Ассоциативные ядра а) не имеет прямого афферентного входа с периферии; б) афференты этих ядер берут
- 8. 3. Моторные ядра а) к моторным ядрам таламуса относится передне-центральное и вентролатеральные ядра, которые имеют вход
- 9. Неспецифические ядра таламуса Филогенетически наиболее древние. Получают афференты от различных сенсорных систем. Нейроны этих ядер полимодальны.
- 10. Генерирование α–ритма таламусом.
- 11. Основные структуры гипоталамуса Гипоталамус осуществляет связь между нервной и гуморальной системами регуляции. В нём выделяют 48
- 12. Локализация центров в гипоталамусе регулирующих внутренний гомеостаз Задний гипоталамус – центр преимущественно химической терморегуляции (за счёт
- 13. ФУНКЦИИ ГИПОТАЛАМУСА Высший центр регуляции вегетативной нервной системы Высший центр регуляции эндокринных функций Регуляция мотиваций пищевого
- 14. Нейроны гипоталамуса имеют следующие особенности: Отсутствует гематоэнцефалический барьер между нейронами и кровью. Чувствительны к составу омывающей
- 15. Опыт с самораздражением Олдса
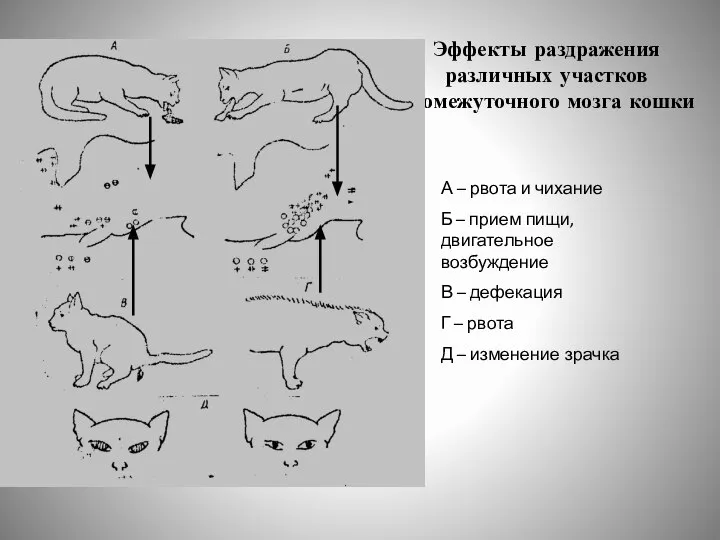
- 16. Эффекты раздражения различных участков промежуточного мозга кошки А – рвота и чихание Б – прием пищи,
- 17. Физиология мозжечка
- 18. мозжечок На долю мозжечка приходится от 8 до 12% веса головного мозга. Общая поверхность коры мозжечка
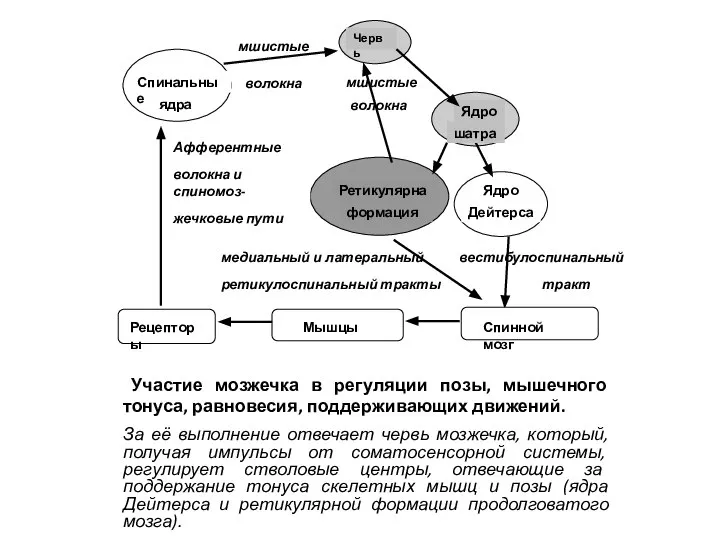
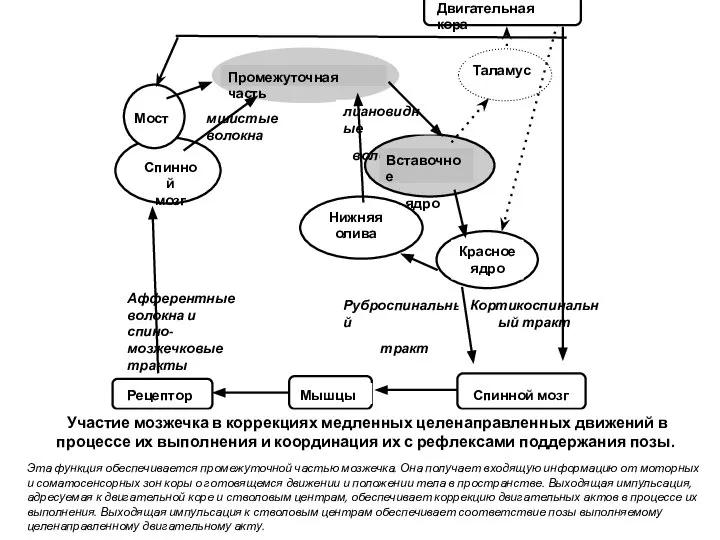
- 19. ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ И ИХ КООРДИНАЦИЯ С
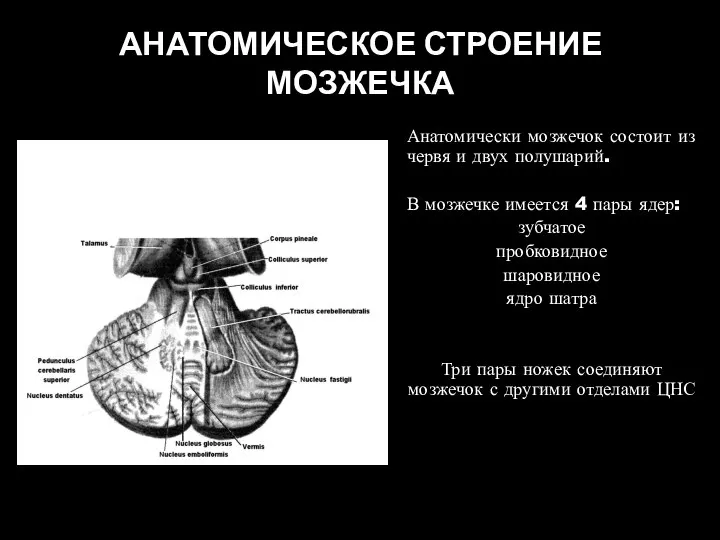
- 20. АНАТОМИЧЕСКОЕ СТРОЕНИЕ МОЗЖЕЧКА Анатомически мозжечок состоит из червя и двух полушарий. В мозжечке имеется 4 пары
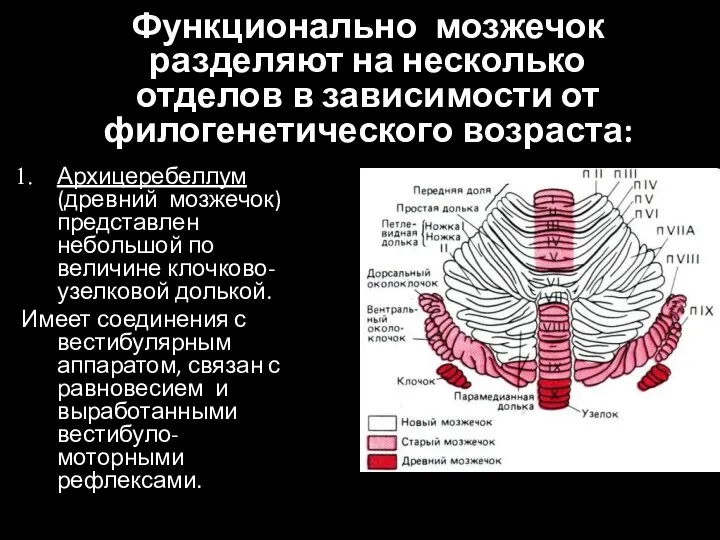
- 21. Функционально мозжечок разделяют на несколько отделов в зависимости от филогенетического возраста: Архицеребеллум (древний мозжечок) представлен небольшой
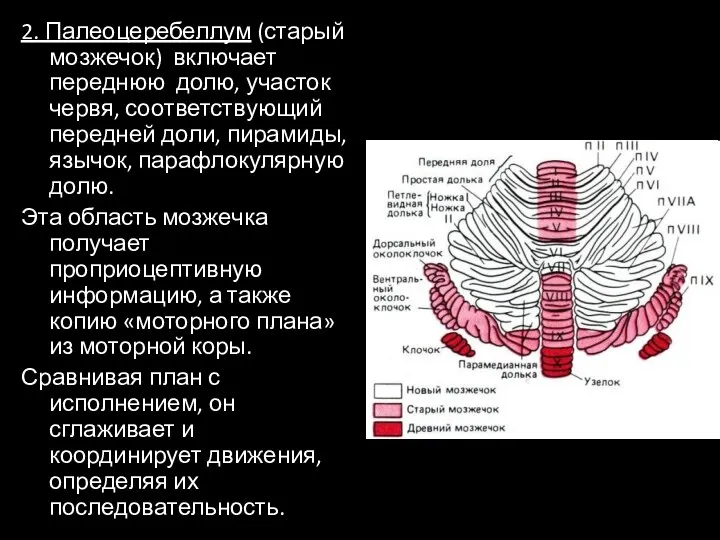
- 22. 2. Палеоцеребеллум (старый мозжечок) включает переднюю долю, участок червя, соответствующий передней доли, пирамиды, язычок, парафлокулярную долю.
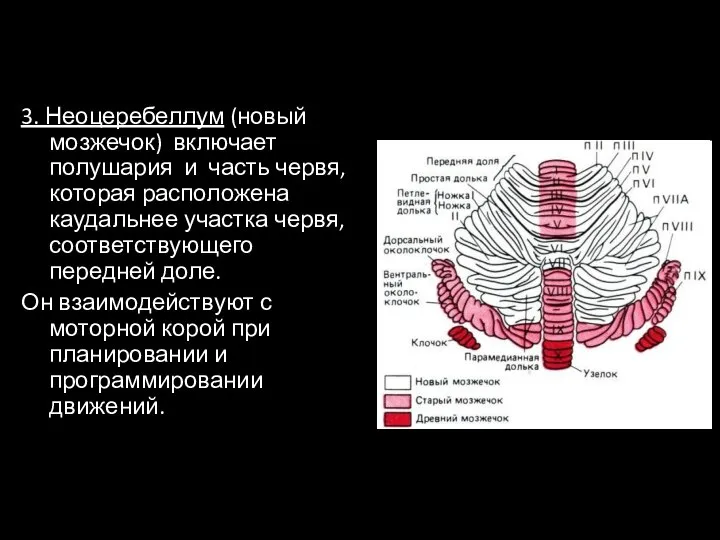
- 23. 3. Неоцеребеллум (новый мозжечок) включает полушария и часть червя, которая расположена каудальнее участка червя, соответствующего передней
- 24. СВЯЗИ КОРЫ МОЗЖЕЧКА АФФЕРЕНТНЫЕ СВЯЗИ МОХОВИДНЫЕ ВОЛОКНА: от 1) Вестибулярных ядер - вестибулоцеребеллярные тракты 2) Спинного
- 25. Связи ядер мозжечка АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА ЗУБЧАТЫЕ ЯДРА: от коры полушарий
- 30. Последствия удаления мозжечка Фаза раздражения. Длится несколько суток . Причиной является: кровоизлияние, отёк тканей, раздражение мозга.
- 31. ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА ТРИАДА ЛЮЧИАНИ: атония, астазия, астения ТРИАДА ШАРКО: нистагм, тремор, скандированная речь АТАКСИЯ (пьяная
- 32. Характер движений после удаления мозжечка
- 33. ВЕГЕТАТИВНЫЕ ФУНКЦИИ МОЗЖЕЧКА Как и симпатическая система, мозжечок выполняет адаптационно-трофическую роль; способствуя активации резервов организма для
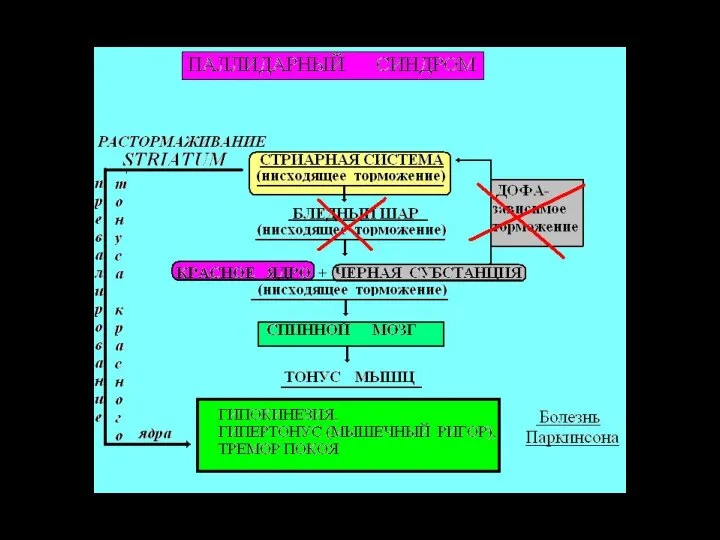
- 34. БАЗАЛЬНЫЕ ЯДРА Хвостатое ядро Скорлупа Бледный шар (наружный сегмент) Бледный шар (внутренний сегмент) Субталамическое ядро Черная
- 36. Афференты к стриатуму: от всех областей коры от таламуса от черной субстанции; Эфференты от стриатума: к
- 37. ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ 1. Центры координации сочетанных двигательных актов 2. Центры контроля координации тонуса мышц и
- 38. Циркуляция возбуждений между базальными ганглиями и корой
- 39. Концепция множественных раздельных, параллельно действующих функциональных петель. Модульная организация.
- 40. Функциональные петли, проходящие через базальные ганглии ПМК–премоторная кора; ДДК–дополнительная двигательная область коры; ДК двигательная кора; ССК–соматосенсорная
- 43. Красное ядро – стимуляция сгибателей, торможение разгибателей В случае перерезки головного мозга ниже красного ядра возникает
- 44. Болезнь Паркинсона: ригидность, тремор покоя, акинезия, эмоциональная тупость
- 45. Паркинсонизм Причина - ↓ меланина (предшественника дофамина) в черной субстанции. Гипокинетические и гиперкинетические признаки: тремор возникает
- 47. Патология базальных ядер Базальные ядра совместно с корой больших полушарий контролируют два важных показателя — амплитуду
- 48. Патология ядер Атетоз – непроизвольные медленные стереотипные, вычурные движения небольшого объёма в дистальных отделах конечностей, нередко
- 49. Атетоз - наблюдается в виде медленных, червеобразных, вычурных движений в кистях, пальца рук и стоп.
- 50. Торсионный спазм
- 51. Лицевой гемиспазм
- 52. Гемибаллизм — производимые с большой силой крупные, размашистые бросковые движения конечностей
- 53. Хорея Хантингтона (Гентингтона) – характеризуется нарастающим гиперкинезом (быстрые беспорядочные нерегулярные движения) пока пациент не упадёт. Речь
- 54. Хорея Хантингтона
- 55. ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ Поражения хвостатого ядра: гиперкинезы- атетозы и хорея (пляска святого Витта) Поражения паллидум:
- 56. Особенности двигательных программ мозжечка и базальных ганглиев Мозжечок Двигательные программы приобретённые Регуляция быстрых движений Поражение мозжечка
- 57. ЭКСТРАПИРАМИДНАЯ СИСТЕМА совокупность структур мозга, включающая: часть коры головного мозга, базальные ганглии, ретикулярную формацию ствола, красное
- 59. ЛИМБИЧЕСКАЯ СИСТЕМА
- 60. Расположение лимбической системы в виде кольца по краю неокортекса (А); афферентные и эфферентные связи лимбической системы
- 62. 2 ЛИМБИЧЕСКИХ КРУГА БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела - мамиллярно-таламический пучок - таламус
- 63. Основные функции лимбики: 1. Организация вегетативно-соматических компонентов эмоций. 2. Организация кратковременной и долговременной памяти. 3. Участие
- 64. Функциональная структура лимбики по МакЛину (1970) 1. Нижний отдел - миндалина и гиппокамп - центры эмоций
- 65. ГИППОКАМП Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными между собой и другими структурами. Модуль включает специализированные
- 66. Здесь находятся нейроны, которые позволяют человеку ориентироваться в пространстве, точно определять местоположение, запоминать и прокладывать маршруты.
- 67. Связь модулей создаёт условие циркулирования активности в гиппокампе при обучении. При этом возрастает нейросекреция и увеличивается
- 68. Реактивность гиппокампа. Усиление - θ-ритма у животных коррелирует с высоким уровнем эмоционального напряжения: страх, агрессия, пищевое
- 69. Консолидация памяти. После двустороннего удаления части гиппокампа, пациенты могли удовлетворительно вспоминать полученные прежде знания. Однако, они
- 70. По своему происхождению гиппокамп является частью обонятельной коры, и на эволюционно ранних стадиях развития мозга ему
- 71. Миндалевидное тело Комплекс ядер, имеющих обширные двусторонние связи с гипоталамусом и всеми частями лимбической системы, а
- 72. Стимуляция миндалевидного тела вызывает те же эффекты, что и стимуляция гипоталамуса: увеличение или уменьшение АД, увеличение
- 73. Кроме того, стимуляция некоторых ядер может порождать реакции ярости, избегания опасности, тяжёлой боли и страха. Наконец,
- 74. До операции После удаления миндалин у Дейва Влияние миндалин на иерархические отношения в стае
- 75. Миндалина образует также обширные связи с обонятельной луковицей. Благодаря этим связям обоняние у животных участвует в
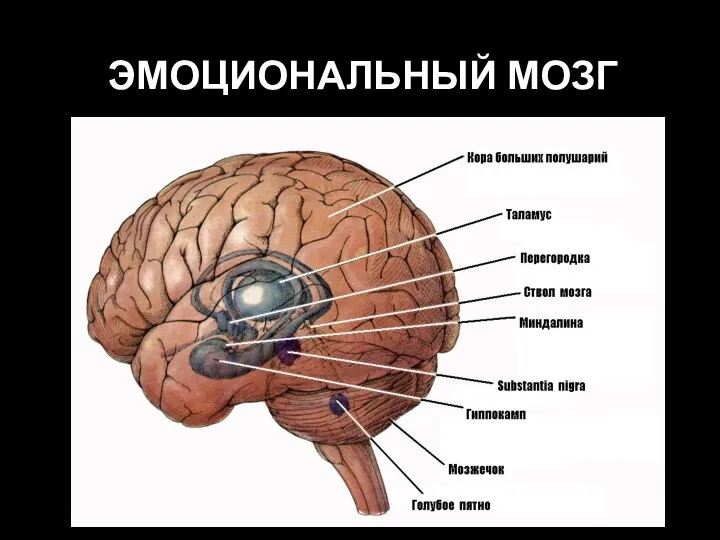
- 76. ЭМОЦИОНАЛЬНЫЙ МОЗГ
- 77. цитаты Некоторые из эмоциональных выражений, свойственных лицу человека, очень сходны с тем, которые были свойственны нашим
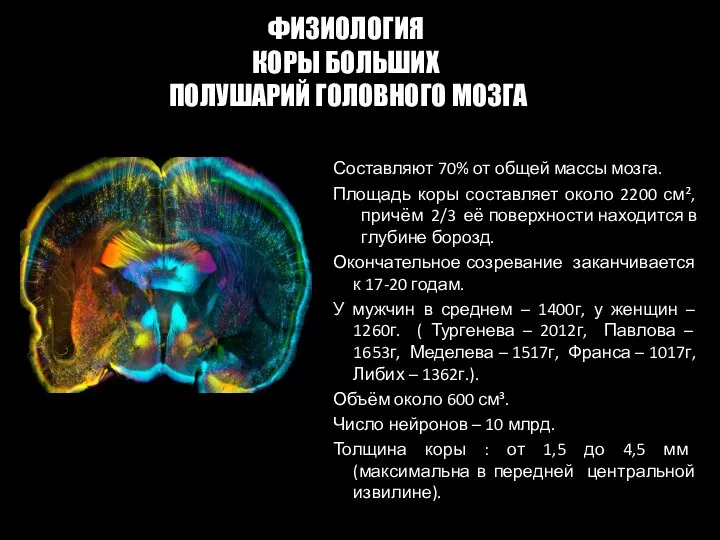
- 78. Составляют 70% от общей массы мозга. Площадь коры составляет около 2200 см², причём 2/3 её поверхности
- 79. ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.) МОТОРНЫЕ ( первичные, вторичные, комплексные) АССОЦИАТИВНЫЕ
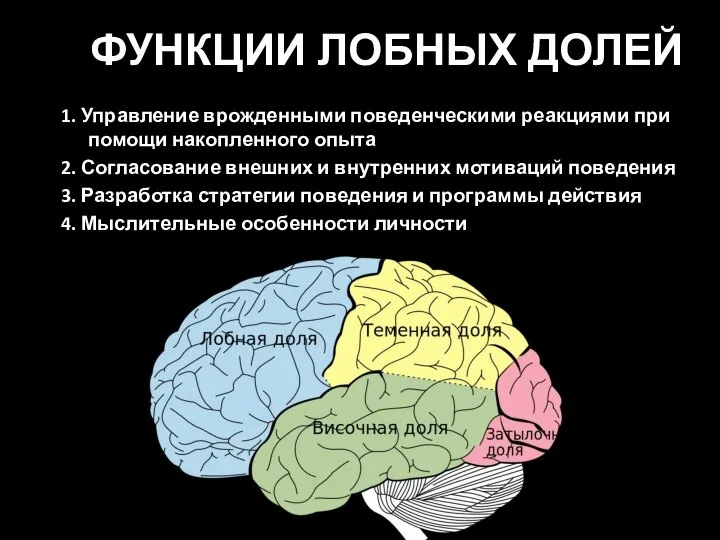
- 80. ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ 1. Управление врожденными поведенческими реакциями при помощи накопленного опыта 2. Согласование внешних и
- 81. Функции теменных долей Восприятие информации от болевых, тактильных и температурных рецепторов. Интеграция зрительных и тактильных восприятий
- 82. Функции височных долей Восприятие и анализ слуховых раздражений. Слуховой и зрительный контроль речи. Восприятие вестибулярной информации.
- 83. Функции затылочных долей Восприятие зрительной информации: Наличие и интенсивность зрительного сигнала. Цвет, форма размеры и качество.
- 84. Слои коры больших полушарий 1 - молекулярный. 2 - наружный зернистый. 3 - наружных пирамид. 4
- 85. Функциональная организация коры Функциональная единица коры - вертикальная колонка диаметром около 500 мкм – макромодуль. Колонка
- 86. Структурный микромодуль сенсомоторной коры III—VI — корковые слои; 1 — интернейроны, 2 — афферентный вход, контактирующий
- 87. Колонкам корковых нейронов присуща функциональная специализация. Так, в соматосенсорной коре каждая колонка иннервирует только одно спинальное
- 88. Колончатая организация корковых нейронов. А. Сагиттальный разрез через постцентральную извилину. Б. Рецептивные поля (ладонь обезьяны) пяти
- 89. Локализационная карта Клейста. Наружная поверхность мозга
- 90. Первичная моторная кора. Грубая топография представительства различных мышц начиная от мышц лица (вблизи от сильвиевой борозды)
- 91. Премоторная кора Располагается кпереди от первичной моторной коры, её топографическая организация подобна организации первичной коры. В
- 92. Дополнительная двигательная кора Располагается в продольной щели и функционирует совместно с премоторной областью. Обеспечивает движения, поддерживающие
- 93. Схема связей в двигательной системе.
- 94. Центральная регуляция двигательной активности
- 95. Нарушение функций моторной коры Повреждение пирамидных путей приводит к гемипарезу — мышечный спазм поражённых мышц на
- 96. Асимметрия полушарий мозга
- 97. Анатомические различия между двумя полушариями Правая лобная доля в норме толще, чем левая, а левая затылочная
- 98. Химические различия между двумя полушариями В путях между полосатым телом и чёрным веществом выше содержание дофамина:
- 99. СПЕЦИАЛИЗАЦИЯ И ДОМИНИРОВАНИЕ ПОЛУШАРИЙ. Считается, что ассиметрия мозга формируется при созревании мозолистого тела. Функции устной и
- 100. Межполушарные различия
- 101. Левое полушарие Играет преимущественную роль в экспрессивной и импрессивной речи, в чтении, письме, вербальной памяти и
- 102. Правое полушарие Выступает ведущим для неречевого, например, музыкального слуха, зрительно-пространственной ориентации, невербальной памяти, критичности. Правое полушарие
- 104. Скачать презентацию
Слайд 2Таламус
Таламус подразделяют на эпиталамус, дорсальный таламус и вентральный таламус.
Эпиталамус имеет связи
Таламус
Таламус подразделяют на эпиталамус, дорсальный таламус и вентральный таламус.
Эпиталамус имеет связи
Слайд 3С развитием коры головного мозга, он утратил эту функцию. Однако, остался высшим
С развитием коры головного мозга, он утратил эту функцию. Однако, остался высшим

Слайд 4Основные ядра таламуса
Специфические ядра переключающие
ассоциативные
моторные
Неспецифические ядра
Основные ядра таламуса
Специфические ядра переключающие
ассоциативные
моторные
Неспецифические ядра

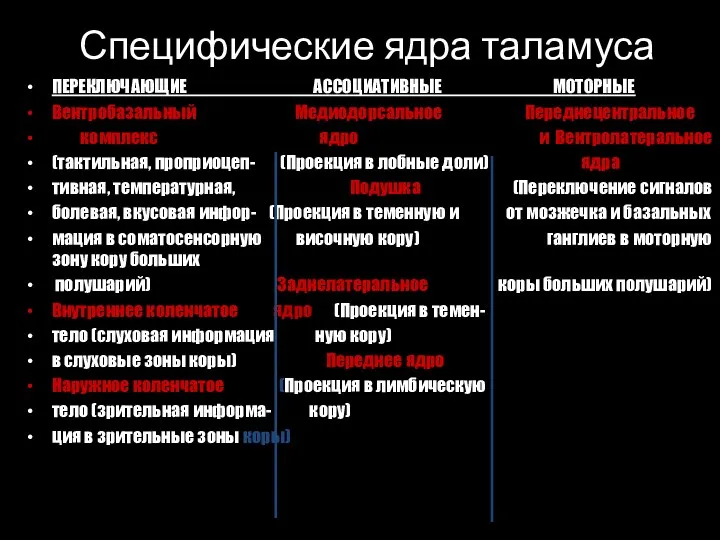
Слайд 5Специфические ядра таламуса
ПЕРЕКЛЮЧАЮЩИЕ АССОЦИАТИВНЫЕ МОТОРНЫЕ
Вентробазальный Медиодорсальное Переднецентральное
комплекс ядро и Вентролатеральное
(тактильная, проприоцеп-
Специфические ядра таламуса
ПЕРЕКЛЮЧАЮЩИЕ АССОЦИАТИВНЫЕ МОТОРНЫЕ
Вентробазальный Медиодорсальное Переднецентральное
комплекс ядро и Вентролатеральное
(тактильная, проприоцеп-
Слайд 61. Специфические ядра (релейные)
а) получают афференты от всех трактов (исключая обонятельный);
б) имеют
1. Специфические ядра (релейные)
а) получают афференты от всех трактов (исключая обонятельный);
б) имеют

Слайд 72. Ассоциативные ядра
а) не имеет прямого афферентного входа с периферии;
б)
2. Ассоциативные ядра
а) не имеет прямого афферентного входа с периферии;
б)

Слайд 83. Моторные ядра
а) к моторным ядрам таламуса относится передне-центральное и вентролатеральные
3. Моторные ядра
а) к моторным ядрам таламуса относится передне-центральное и вентролатеральные

Слайд 9Неспецифические ядра таламуса
Филогенетически наиболее древние. Получают афференты от различных сенсорных систем. Нейроны
Неспецифические ядра таламуса
Филогенетически наиболее древние. Получают афференты от различных сенсорных систем. Нейроны

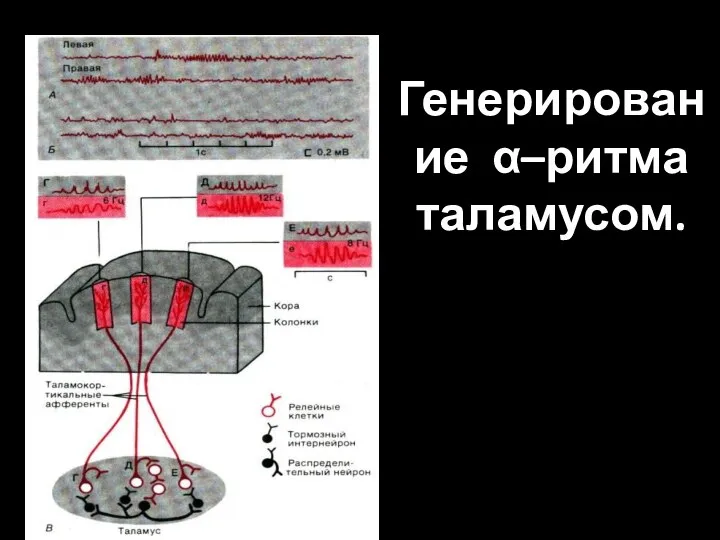
Слайд 10Генерирование α–ритма таламусом.
Генерирование α–ритма таламусом.
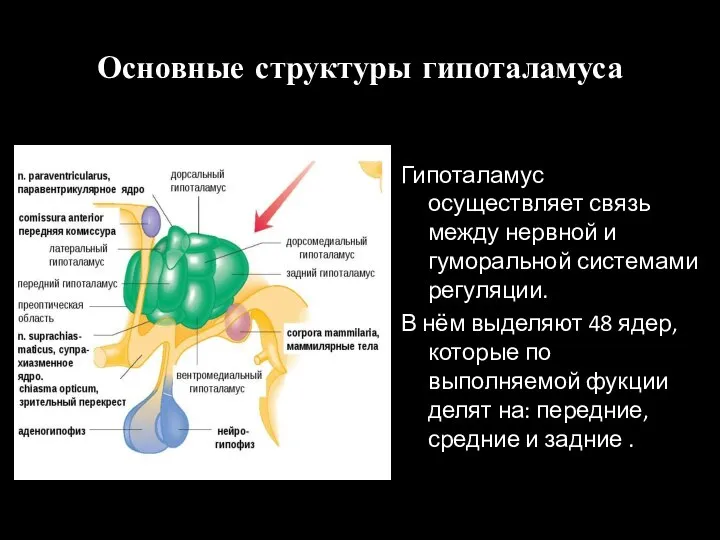
Слайд 11Основные структуры гипоталамуса
Гипоталамус осуществляет связь между нервной и гуморальной системами регуляции.
В
Основные структуры гипоталамуса
Гипоталамус осуществляет связь между нервной и гуморальной системами регуляции.
В
Слайд 12Локализация центров в гипоталамусе регулирующих внутренний гомеостаз
Задний гипоталамус – центр преимущественно
Локализация центров в гипоталамусе регулирующих внутренний гомеостаз
Задний гипоталамус – центр преимущественно

Слайд 13ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция мотиваций
ФУНКЦИИ ГИПОТАЛАМУСА
Высший центр регуляции вегетативной нервной системы
Высший центр регуляции эндокринных функций
Регуляция мотиваций

Слайд 14
Нейроны гипоталамуса имеют следующие особенности:
Отсутствует гематоэнцефалический барьер между нейронами и кровью.
Чувствительны к
Нейроны гипоталамуса имеют следующие особенности:
Отсутствует гематоэнцефалический барьер между нейронами и кровью.
Чувствительны к

Слайд 15Опыт с самораздражением Олдса
Опыт с самораздражением Олдса

Слайд 16Эффекты раздражения различных участков промежуточного мозга кошки
А – рвота и чихание
Б –
Эффекты раздражения различных участков промежуточного мозга кошки
А – рвота и чихание
Б –
Слайд 17Физиология мозжечка
Физиология мозжечка
Слайд 18мозжечок
На долю мозжечка приходится от 8 до 12% веса головного мозга.
Общая поверхность
мозжечок
На долю мозжечка приходится от 8 до 12% веса головного мозга.
Общая поверхность

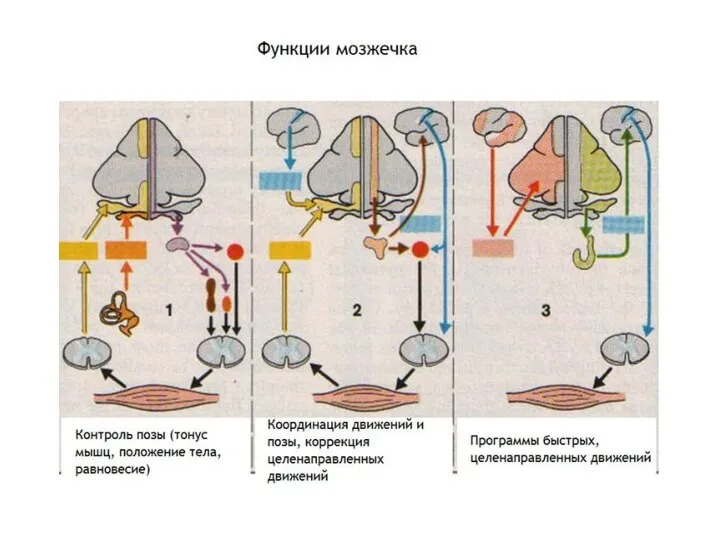
Слайд 19ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА
КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ И ИХ
ОСНОВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
РЕГУЛЯЦИЯ ПОЗЫ И МЫШЕЧНОГО ТОНУСА
КОРРЕКЦИЯ МЕДЛЕННЫХ ЦЕЛЕНАПРАВЛЕННЫХ ДВИЖЕНИЙ И ИХ

Слайд 20АНАТОМИЧЕСКОЕ СТРОЕНИЕ МОЗЖЕЧКА
Анатомически мозжечок состоит из червя и двух полушарий.
В мозжечке имеется
АНАТОМИЧЕСКОЕ СТРОЕНИЕ МОЗЖЕЧКА
Анатомически мозжечок состоит из червя и двух полушарий.
В мозжечке имеется
Слайд 21Функционально мозжечок разделяют на несколько отделов в зависимости от филогенетического возраста:
Архицеребеллум (древний
Функционально мозжечок разделяют на несколько отделов в зависимости от филогенетического возраста:
Архицеребеллум (древний
Слайд 222. Палеоцеребеллум (старый мозжечок) включает переднюю долю, участок червя, соответствующий передней доли,
2. Палеоцеребеллум (старый мозжечок) включает переднюю долю, участок червя, соответствующий передней доли,
Слайд 233. Неоцеребеллум (новый мозжечок) включает полушария и часть червя, которая расположена каудальнее
3. Неоцеребеллум (новый мозжечок) включает полушария и часть червя, которая расположена каудальнее
Слайд 24СВЯЗИ КОРЫ МОЗЖЕЧКА
АФФЕРЕНТНЫЕ СВЯЗИ
МОХОВИДНЫЕ ВОЛОКНА: от
1) Вестибулярных ядер - вестибулоцеребеллярные тракты
2) Спинного
СВЯЗИ КОРЫ МОЗЖЕЧКА
АФФЕРЕНТНЫЕ СВЯЗИ
МОХОВИДНЫЕ ВОЛОКНА: от
1) Вестибулярных ядер - вестибулоцеребеллярные тракты
2) Спинного

Слайд 25Связи ядер мозжечка
АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА
ЗУБЧАТЫЕ ЯДРА: от
Связи ядер мозжечка
АФФЕРЕНТНЫЕ СВЯЗИ ВСЕХ ЯДЕР - ОТ КОРЫ МОЗЖЕЧКА
ЗУБЧАТЫЕ ЯДРА: от

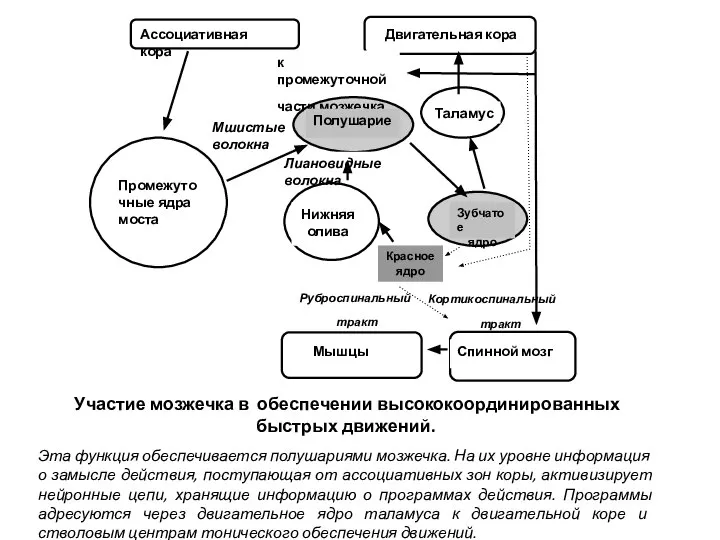
Слайд 30Последствия удаления мозжечка
Фаза раздражения. Длится несколько суток . Причиной является: кровоизлияние, отёк
Последствия удаления мозжечка
Фаза раздражения. Длится несколько суток . Причиной является: кровоизлияние, отёк

Слайд 31ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,
ПРИЗНАКИ ПОРАЖЕНИЯ МОЗЖЕЧКА
ТРИАДА ЛЮЧИАНИ: атония,
астазия,
астения
ТРИАДА ШАРКО: нистагм,

Слайд 32Характер движений после удаления мозжечка
Характер движений после удаления мозжечка

Слайд 33ВЕГЕТАТИВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
Как и симпатическая система, мозжечок выполняет адаптационно-трофическую роль; способствуя активации
ВЕГЕТАТИВНЫЕ ФУНКЦИИ МОЗЖЕЧКА
Как и симпатическая система, мозжечок выполняет адаптационно-трофическую роль; способствуя активации

Слайд 34БАЗАЛЬНЫЕ ЯДРА
Хвостатое
ядро
Скорлупа
Бледный шар
(наружный сегмент)
Бледный шар
(внутренний сегмент)
Субталамическое ядро
Черная субстанция
Миндалина
Скорлупа + Хвостатое
БАЗАЛЬНЫЕ ЯДРА
Хвостатое
ядро
Скорлупа
Бледный шар
(наружный сегмент)
Бледный шар
(внутренний сегмент)
Субталамическое ядро
Черная субстанция
Миндалина
Скорлупа + Хвостатое
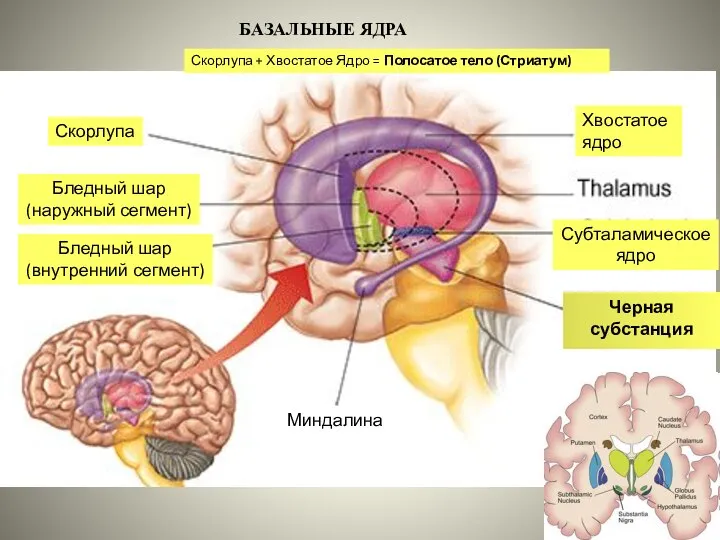
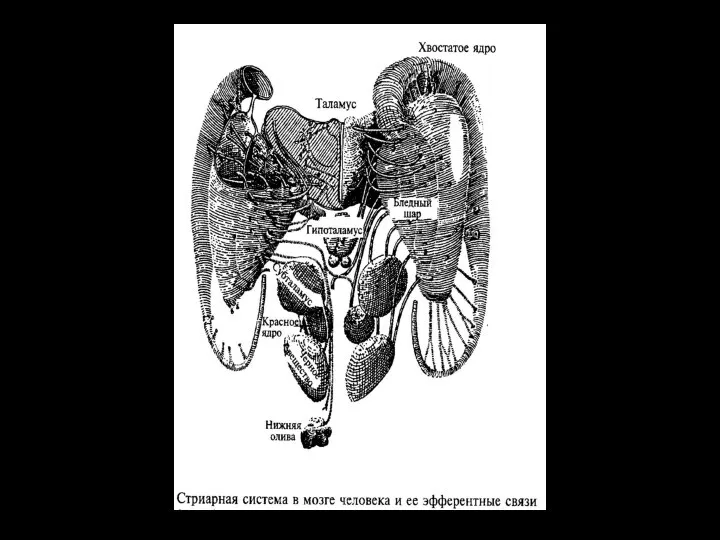
Слайд 36Афференты к стриатуму:
от всех областей коры
от таламуса
от черной субстанции;
Эфференты от стриатума:
к черной
Афференты к стриатуму:
от всех областей коры
от таламуса
от черной субстанции;
Эфференты от стриатума:
к черной

Слайд 37ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
1. Центры координации сочетанных двигательных актов
2. Центры контроля координации тонуса
ФУНКЦИИ БАЗАЛЬНЫХ ГАНГЛИЕВ
1. Центры координации сочетанных двигательных актов
2. Центры контроля координации тонуса

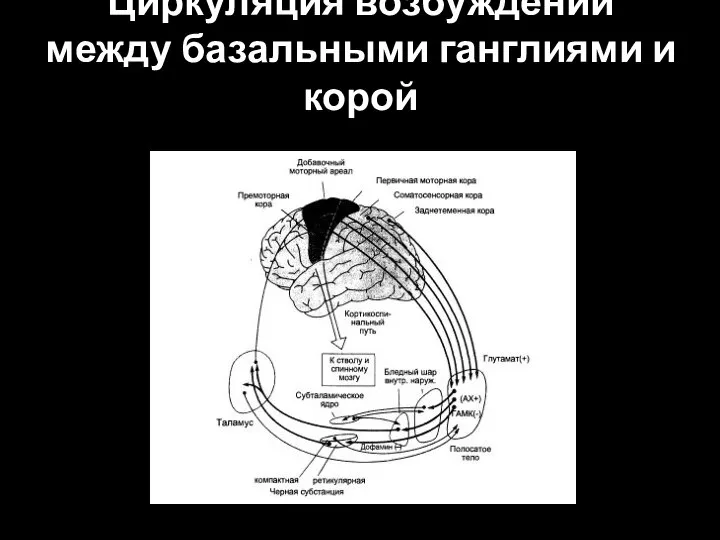
Слайд 38Циркуляция возбуждений между базальными ганглиями и корой
Циркуляция возбуждений между базальными ганглиями и корой
Слайд 39Концепция множественных раздельных, параллельно действующих функциональных петель.
Модульная организация.
Концепция множественных раздельных, параллельно действующих функциональных петель.
Модульная организация.

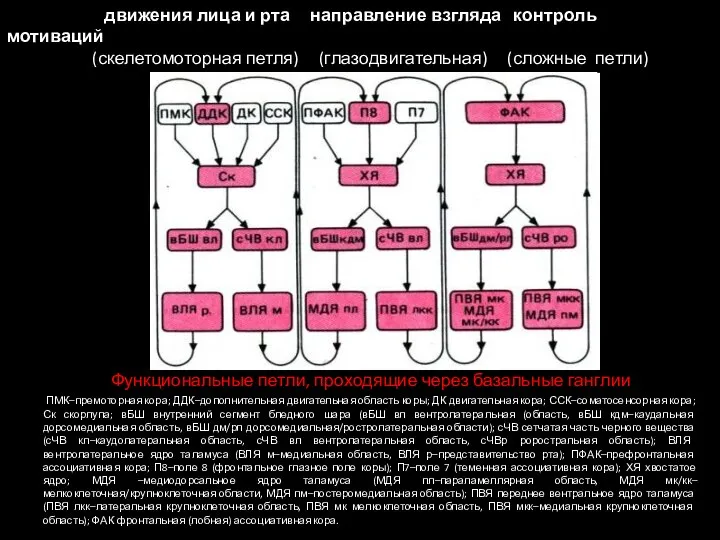
Слайд 40Функциональные петли, проходящие через базальные ганглии
ПМК–премоторная кора; ДДК–дополнительная двигательная область
Функциональные петли, проходящие через базальные ганглии
ПМК–премоторная кора; ДДК–дополнительная двигательная область

Слайд 43Красное ядро – стимуляция сгибателей, торможение разгибателей
В случае перерезки головного мозга ниже
Красное ядро – стимуляция сгибателей, торможение разгибателей
В случае перерезки головного мозга ниже

Слайд 44
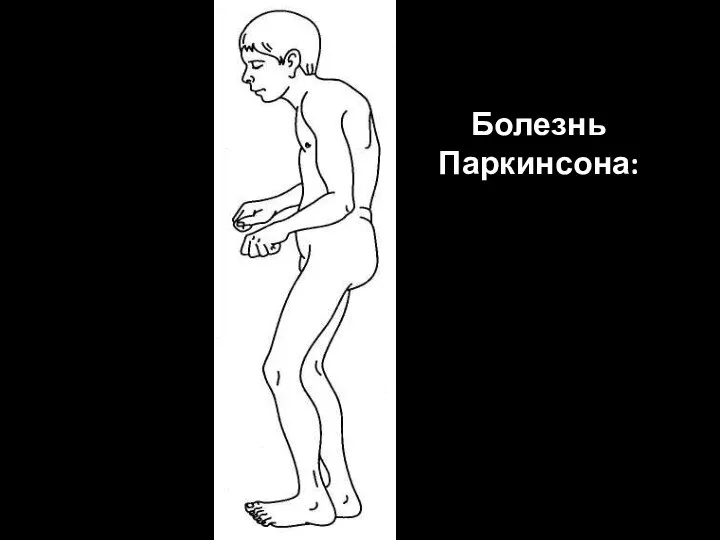
Болезнь Паркинсона:
ригидность,
тремор покоя,
акинезия,
эмоциональная тупость
Болезнь Паркинсона:
ригидность,
тремор покоя,
акинезия,
эмоциональная тупость
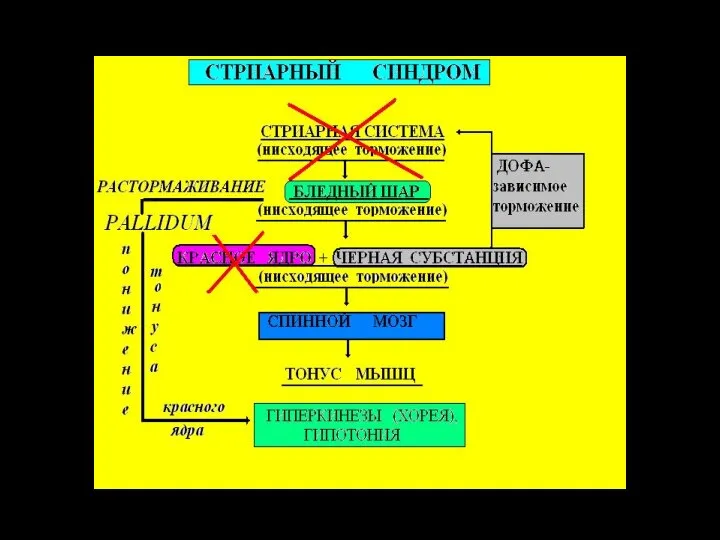
Слайд 45Паркинсонизм
Причина - ↓ меланина (предшественника дофамина) в черной субстанции.
Гипокинетические и гиперкинетические признаки:
Паркинсонизм
Причина - ↓ меланина (предшественника дофамина) в черной субстанции.
Гипокинетические и гиперкинетические признаки:

Слайд 47Патология базальных ядер
Базальные ядра совместно с корой больших полушарий контролируют два
Патология базальных ядер
Базальные ядра совместно с корой больших полушарий контролируют два

Слайд 48Патология ядер
Атетоз – непроизвольные медленные стереотипные, вычурные движения небольшого объёма в дистальных
Патология ядер
Атетоз – непроизвольные медленные стереотипные, вычурные движения небольшого объёма в дистальных

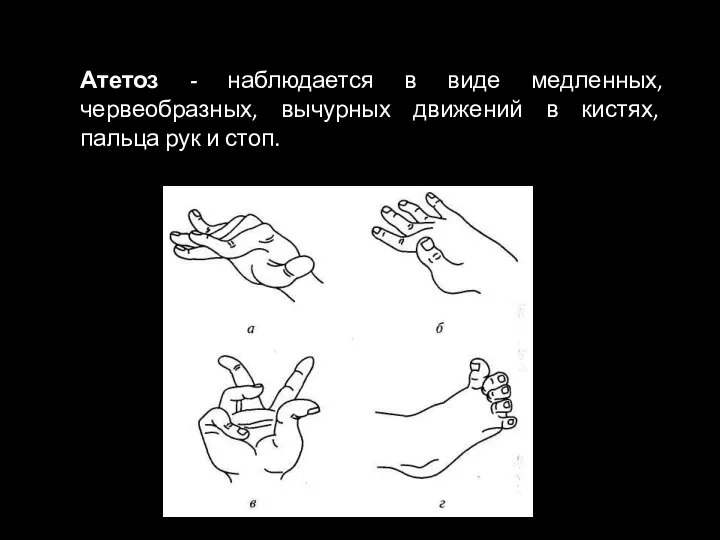
Слайд 49Атетоз - наблюдается в виде медленных, червеобразных, вычурных движений в кистях, пальца
Атетоз - наблюдается в виде медленных, червеобразных, вычурных движений в кистях, пальца
Слайд 50Торсионный спазм
Торсионный спазм

Слайд 51Лицевой гемиспазм
Лицевой гемиспазм

Слайд 52Гемибаллизм — производимые с большой силой крупные, размашистые бросковые движения конечностей
Гемибаллизм — производимые с большой силой крупные, размашистые бросковые движения конечностей

Слайд 53Хорея Хантингтона (Гентингтона) – характеризуется нарастающим гиперкинезом (быстрые беспорядочные нерегулярные движения) пока
Хорея Хантингтона (Гентингтона) – характеризуется нарастающим гиперкинезом (быстрые беспорядочные нерегулярные движения) пока

Слайд 54Хорея Хантингтона
Хорея Хантингтона

Слайд 55ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Поражения хвостатого ядра:
гиперкинезы- атетозы и хорея
ЭФФЕКТЫ ПОРАЖЕНИЯ СТРИОПАЛЛИДАРНОЙ СИСТЕМЫ
Поражения хвостатого ядра:
гиперкинезы- атетозы и хорея

Слайд 56Особенности двигательных программ мозжечка и базальных ганглиев
Мозжечок
Двигательные программы приобретённые
Регуляция быстрых движений
Поражение
Особенности двигательных программ мозжечка и базальных ганглиев
Мозжечок
Двигательные программы приобретённые
Регуляция быстрых движений
Поражение

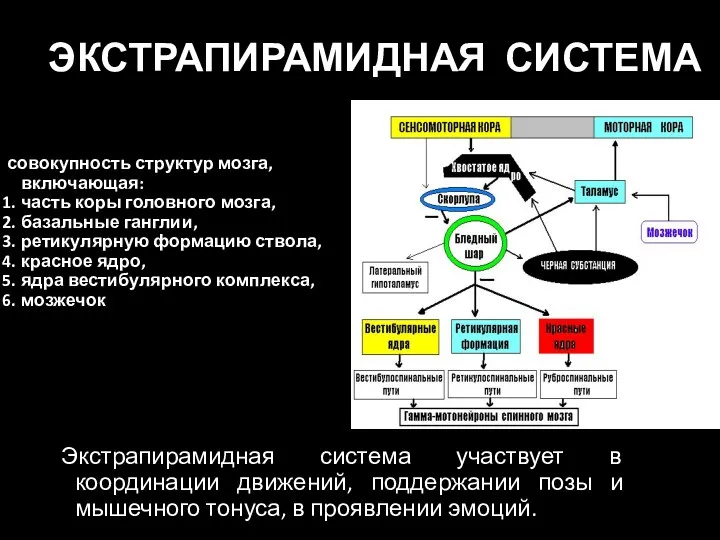
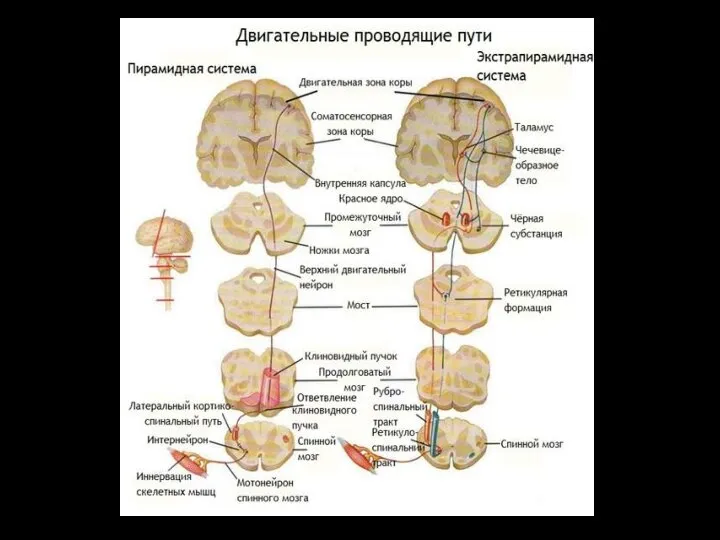
Слайд 57ЭКСТРАПИРАМИДНАЯ СИСТЕМА
совокупность структур мозга, включающая:
часть коры головного мозга,
базальные ганглии,
ретикулярную формацию ствола,
ЭКСТРАПИРАМИДНАЯ СИСТЕМА
совокупность структур мозга, включающая:
часть коры головного мозга,
базальные ганглии,
ретикулярную формацию ствола,
Слайд 59ЛИМБИЧЕСКАЯ СИСТЕМА
ЛИМБИЧЕСКАЯ СИСТЕМА
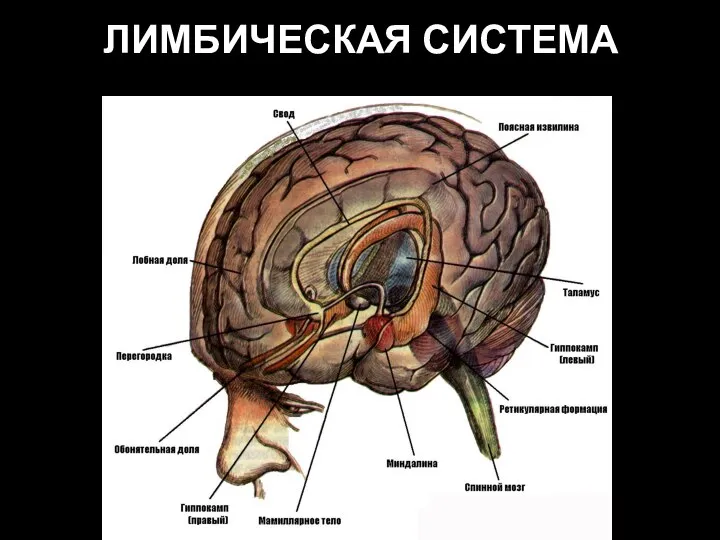
Слайд 60Расположение лимбической системы в виде кольца по краю неокортекса (А); афферентные и
Расположение лимбической системы в виде кольца по краю неокортекса (А); афферентные и
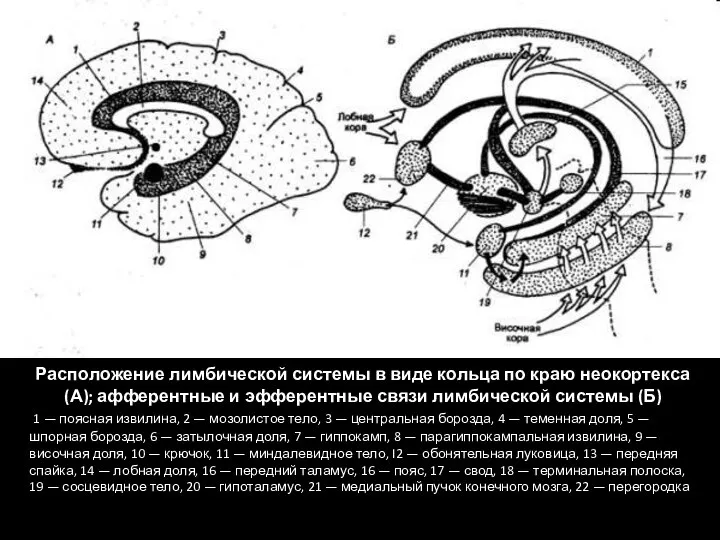
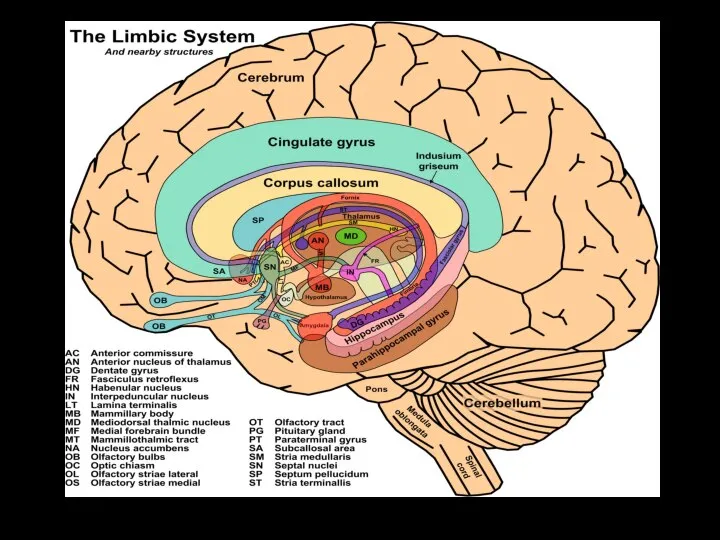
Слайд 622 ЛИМБИЧЕСКИХ КРУГА
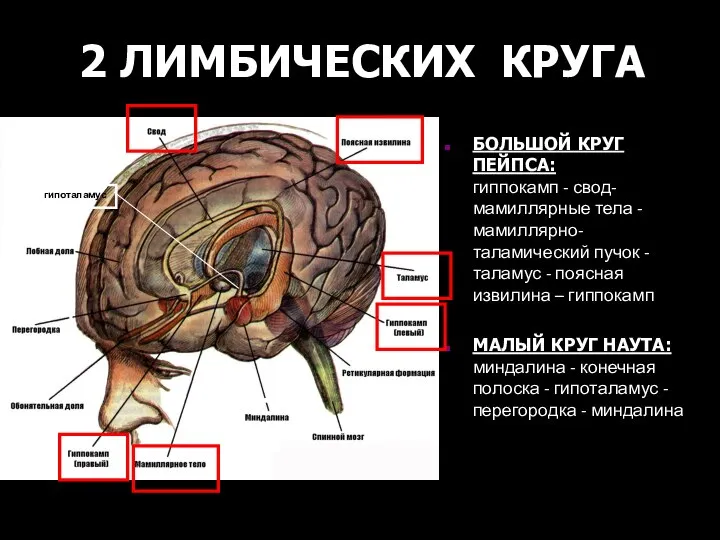
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела - мамиллярно-таламический
2 ЛИМБИЧЕСКИХ КРУГА
БОЛЬШОЙ КРУГ ПЕЙПСА: гиппокамп - свод- мамиллярные тела - мамиллярно-таламический
Слайд 63Основные функции лимбики:
1. Организация вегетативно-соматических компонентов эмоций.
2. Организация кратковременной и долговременной памяти.
3.
Основные функции лимбики:
1. Организация вегетативно-соматических компонентов эмоций.
2. Организация кратковременной и долговременной памяти.
3.

Слайд 64Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и гиппокамп
Функциональная структура лимбики по МакЛину (1970)
1. Нижний отдел - миндалина и гиппокамп

Слайд 65ГИППОКАМП
Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными между собой и другими структурами.
Модуль
ГИППОКАМП
Морфологически гиппокамп представлен стереотипно повторяющимися модулями, связанными между собой и другими структурами.
Модуль

Слайд 66 Здесь находятся нейроны, которые позволяют человеку ориентироваться в пространстве, точно определять
Здесь находятся нейроны, которые позволяют человеку ориентироваться в пространстве, точно определять

Слайд 67 Связь модулей создаёт условие циркулирования активности в гиппокампе при обучении. При
Связь модулей создаёт условие циркулирования активности в гиппокампе при обучении. При

Слайд 68Реактивность гиппокампа.
Усиление - θ-ритма у животных коррелирует с высоким уровнем эмоционального напряжения:
Реактивность гиппокампа.
Усиление - θ-ритма у животных коррелирует с высоким уровнем эмоционального напряжения:

Слайд 69Консолидация памяти.
После двустороннего удаления части гиппокампа, пациенты могли удовлетворительно вспоминать полученные прежде
Консолидация памяти.
После двустороннего удаления части гиппокампа, пациенты могли удовлетворительно вспоминать полученные прежде

Слайд 70По своему происхождению гиппокамп является частью обонятельной коры, и на эволюционно ранних
По своему происхождению гиппокамп является частью обонятельной коры, и на эволюционно ранних

Слайд 71Миндалевидное тело
Комплекс ядер, имеющих обширные двусторонние связи с гипоталамусом и всеми частями
Миндалевидное тело
Комплекс ядер, имеющих обширные двусторонние связи с гипоталамусом и всеми частями

Слайд 72 Стимуляция миндалевидного тела вызывает те же эффекты, что и стимуляция гипоталамуса:
Стимуляция миндалевидного тела вызывает те же эффекты, что и стимуляция гипоталамуса:

Слайд 73 Кроме того, стимуляция некоторых ядер может порождать реакции ярости, избегания опасности,
Кроме того, стимуляция некоторых ядер может порождать реакции ярости, избегания опасности,

Слайд 74До операции
После удаления миндалин у Дейва
Влияние миндалин на иерархические отношения в стае
До операции
После удаления миндалин у Дейва
Влияние миндалин на иерархические отношения в стае
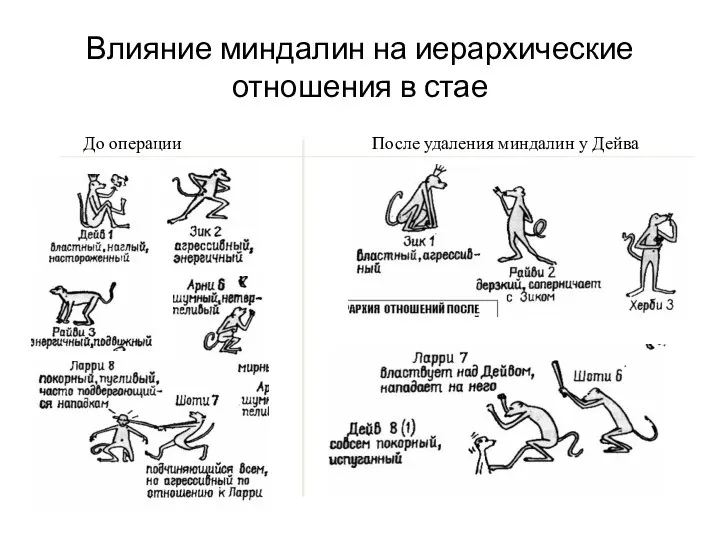
Слайд 75Миндалина образует также обширные связи с обонятельной луковицей. Благодаря этим связям обоняние
Миндалина образует также обширные связи с обонятельной луковицей. Благодаря этим связям обоняние

Слайд 76ЭМОЦИОНАЛЬНЫЙ МОЗГ
ЭМОЦИОНАЛЬНЫЙ МОЗГ
Слайд 77цитаты
Некоторые из эмоциональных выражений, свойственных лицу человека, очень сходны с тем, которые
цитаты
Некоторые из эмоциональных выражений, свойственных лицу человека, очень сходны с тем, которые

Слайд 78Составляют 70% от общей массы мозга.
Площадь коры составляет около 2200 см²,
Площадь коры составляет около 2200 см²,
Слайд 79ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.)
МОТОРНЫЕ ( первичные, вторичные,
ФУНКЦИОНАЛЬНЫЕ ЗОНЫ КОРЫ
СЕНСОРНЫЕ ( зрительные, слуховые, кожные и др.)
МОТОРНЫЕ ( первичные, вторичные,

Слайд 80ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
1. Управление врожденными поведенческими реакциями при помощи накопленного опыта
2. Согласование
ФУНКЦИИ ЛОБНЫХ ДОЛЕЙ
1. Управление врожденными поведенческими реакциями при помощи накопленного опыта
2. Согласование
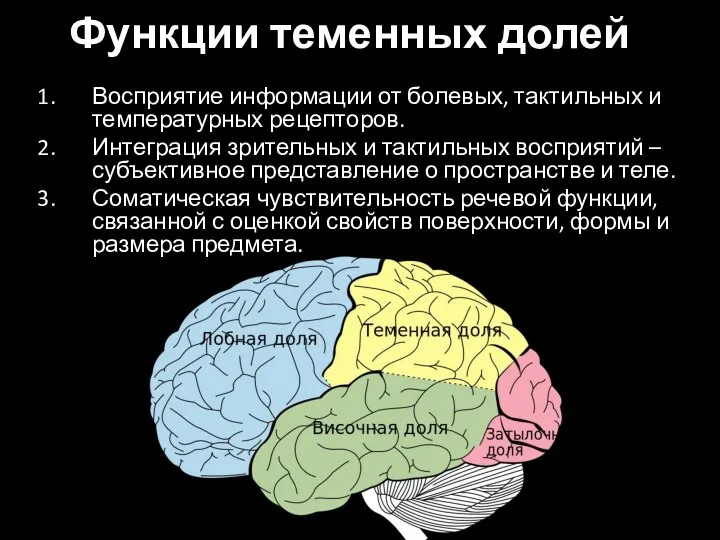
Слайд 81Функции теменных долей
Восприятие информации от болевых, тактильных и температурных рецепторов.
Интеграция зрительных и
Функции теменных долей
Восприятие информации от болевых, тактильных и температурных рецепторов.
Интеграция зрительных и
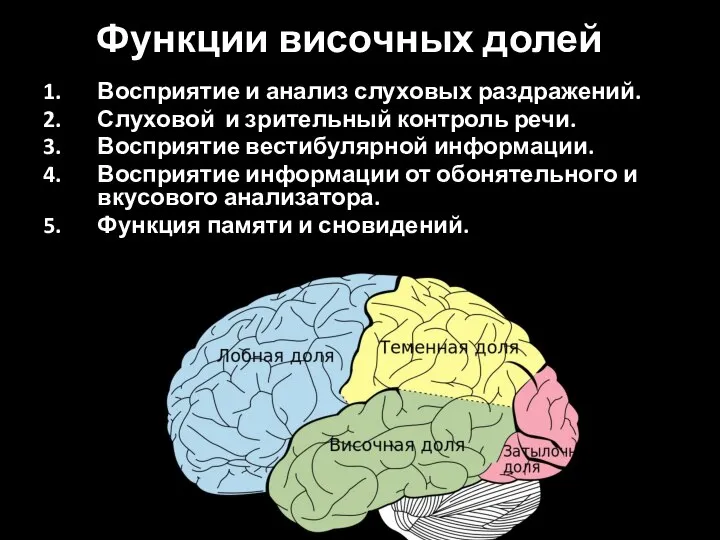
Слайд 82Функции височных долей
Восприятие и анализ слуховых раздражений.
Слуховой и зрительный контроль речи.
Восприятие вестибулярной
Функции височных долей
Восприятие и анализ слуховых раздражений.
Слуховой и зрительный контроль речи.
Восприятие вестибулярной
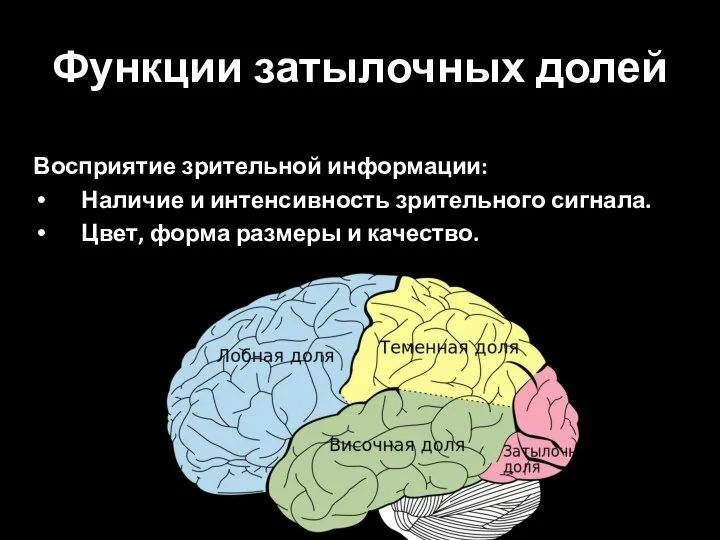
Слайд 83Функции затылочных долей
Восприятие зрительной информации:
Наличие и интенсивность зрительного сигнала.
Цвет, форма размеры и
Функции затылочных долей
Восприятие зрительной информации:
Наличие и интенсивность зрительного сигнала.
Цвет, форма размеры и
Слайд 84Слои коры больших полушарий
1 - молекулярный.
2 - наружный зернистый.
3 -
Слои коры больших полушарий
1 - молекулярный.
2 - наружный зернистый.
3 -

Слайд 85Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500 мкм
Функциональная организация коры
Функциональная единица коры - вертикальная колонка диаметром около 500 мкм

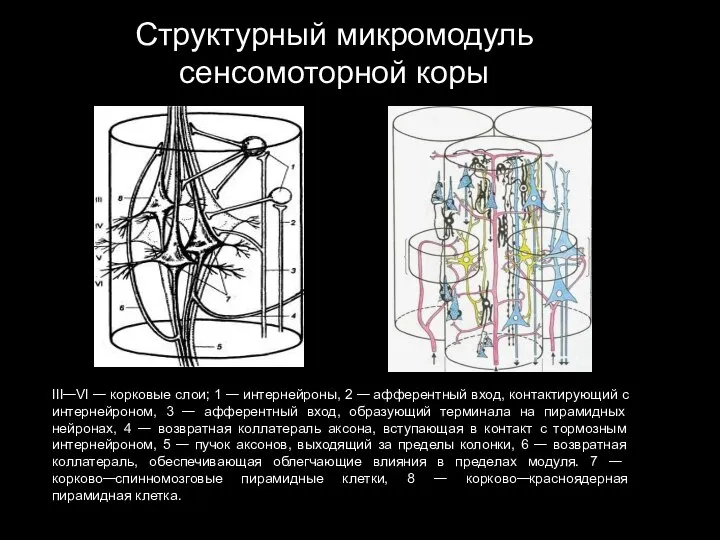
Слайд 86Структурный микромодуль сенсомоторной коры
III—VI — корковые слои; 1 — интернейроны, 2 —
Структурный микромодуль сенсомоторной коры
III—VI — корковые слои; 1 — интернейроны, 2 —
Слайд 87Колонкам корковых нейронов присуща функциональная специализация. Так, в соматосенсорной коре каждая колонка
Колонкам корковых нейронов присуща функциональная специализация. Так, в соматосенсорной коре каждая колонка

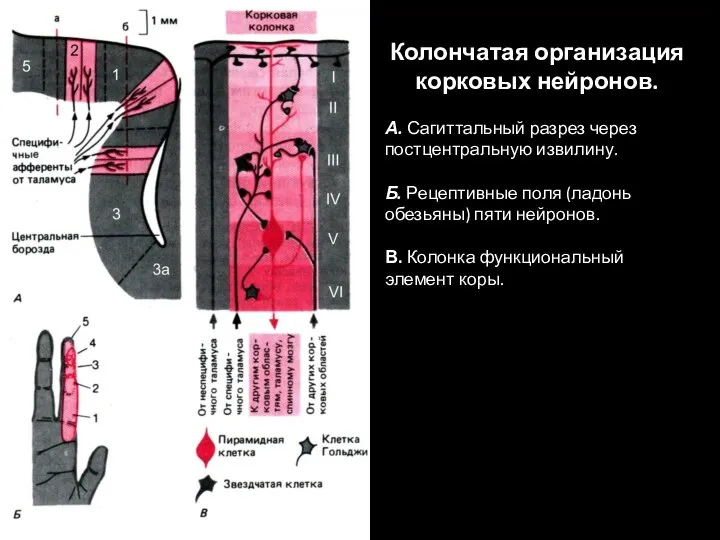
Слайд 88Колончатая организация корковых нейронов.
А. Сагиттальный разрез через постцентральную извилину.
Б. Рецептивные
Колончатая организация корковых нейронов.
А. Сагиттальный разрез через постцентральную извилину.
Б. Рецептивные
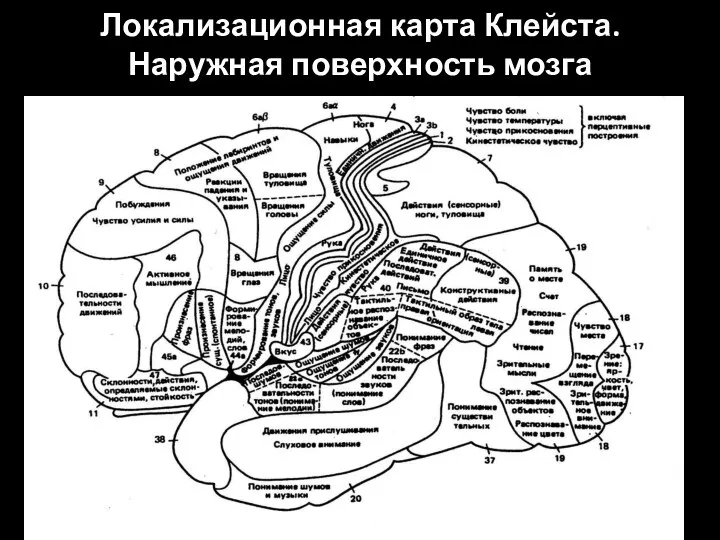
Слайд 89Локализационная карта Клейста. Наружная поверхность мозга
Локализационная карта Клейста. Наружная поверхность мозга
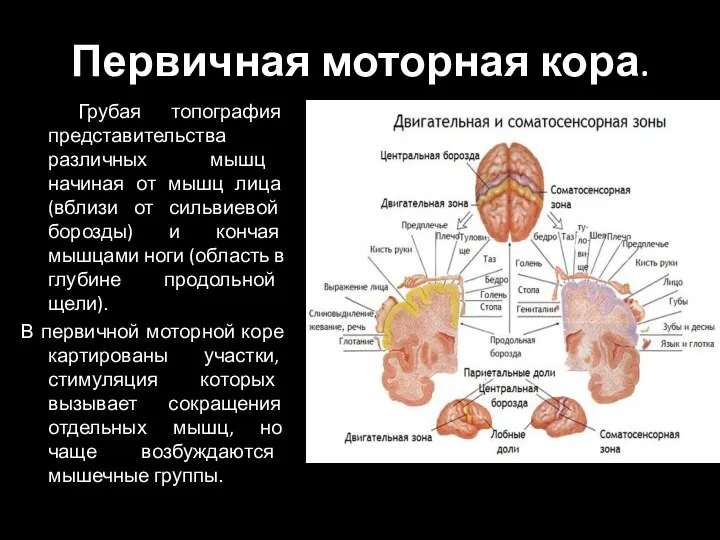
Слайд 90Первичная моторная кора.
Грубая топография представительства различных мышц начиная от мышц лица
Первичная моторная кора.
Грубая топография представительства различных мышц начиная от мышц лица
Слайд 91Премоторная кора
Располагается кпереди от первичной моторной коры, её топографическая организация подобна организации
Премоторная кора
Располагается кпереди от первичной моторной коры, её топографическая организация подобна организации

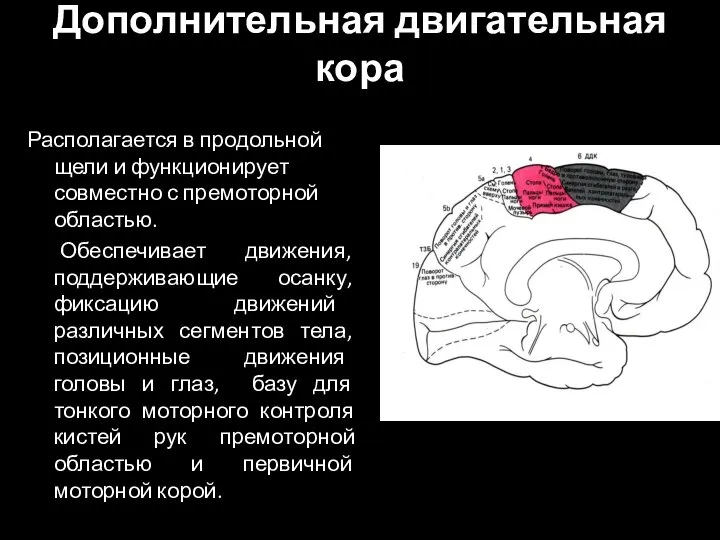
Слайд 92Дополнительная двигательная кора
Располагается в продольной щели и функционирует совместно с премоторной областью.
Дополнительная двигательная кора
Располагается в продольной щели и функционирует совместно с премоторной областью.
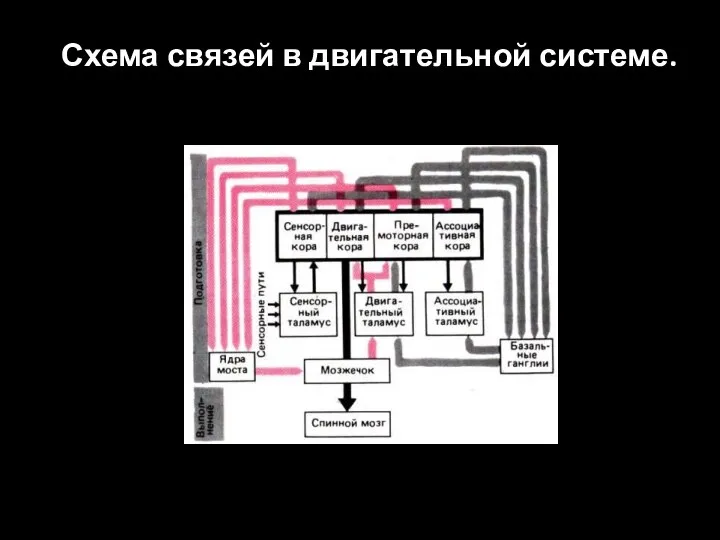
Слайд 93Схема связей в двигательной системе.
Схема связей в двигательной системе.
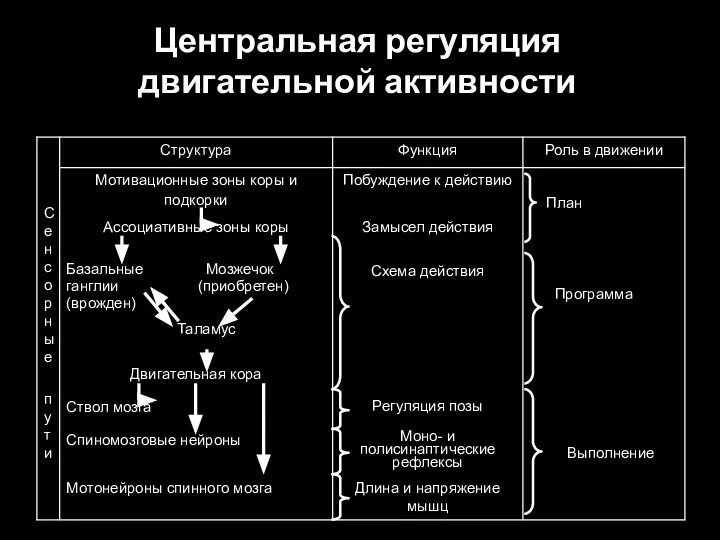
Слайд 94Центральная регуляция двигательной активности
Центральная регуляция двигательной активности
Слайд 95Нарушение функций моторной коры
Повреждение пирамидных путей приводит к гемипарезу — мышечный спазм
Нарушение функций моторной коры
Повреждение пирамидных путей приводит к гемипарезу — мышечный спазм

Слайд 96Асимметрия полушарий мозга
Асимметрия полушарий мозга

Слайд 97Анатомические различия между двумя полушариями
Правая лобная доля в норме толще, чем левая,
Анатомические различия между двумя полушариями
Правая лобная доля в норме толще, чем левая,

Слайд 98Химические различия между двумя полушариями
В путях между полосатым телом и чёрным веществом
Химические различия между двумя полушариями
В путях между полосатым телом и чёрным веществом

Слайд 99СПЕЦИАЛИЗАЦИЯ И ДОМИНИРОВАНИЕ ПОЛУШАРИЙ.
Считается, что ассиметрия мозга формируется при созревании мозолистого тела.
Функции
СПЕЦИАЛИЗАЦИЯ И ДОМИНИРОВАНИЕ ПОЛУШАРИЙ.
Считается, что ассиметрия мозга формируется при созревании мозолистого тела.
Функции

Слайд 100Межполушарные различия
Межполушарные различия

Слайд 101Левое полушарие
Играет преимущественную роль в экспрессивной и импрессивной речи, в чтении, письме,
Левое полушарие
Играет преимущественную роль в экспрессивной и импрессивной речи, в чтении, письме,

Слайд 102Правое полушарие
Выступает ведущим для неречевого, например, музыкального слуха, зрительно-пространственной ориентации, невербальной памяти,
Правое полушарие
Выступает ведущим для неречевого, например, музыкального слуха, зрительно-пространственной ориентации, невербальной памяти,

 Водоемы и рыбы Омской области
Водоемы и рыбы Омской области Рыбы - обитатели водных пространств
Рыбы - обитатели водных пространств Разнообразие комнатных растений - семейство, родина
Разнообразие комнатных растений - семейство, родина Растения - псаммофиты. Экологические условия произрастания растений на песках
Растения - псаммофиты. Экологические условия произрастания растений на песках Эволюция пищеварит. системы у животных
Эволюция пищеварит. системы у животных Пищеварительная система человека
Пищеварительная система человека Презентация на тему Важнейшие отряды птиц
Презентация на тему Важнейшие отряды птиц  Птицы. Виталий Валентинович Бианки
Птицы. Виталий Валентинович Бианки Мой Цветок - Талисман
Мой Цветок - Талисман Как работают гены и можно ли ими управлять
Как работают гены и можно ли ими управлять Организм человека как единая биологическая система
Организм человека как единая биологическая система Наследование признаков, сцепленных с полом
Наследование признаков, сцепленных с полом Пищеварительная система человека. Питание и пищеварение
Пищеварительная система человека. Питание и пищеварение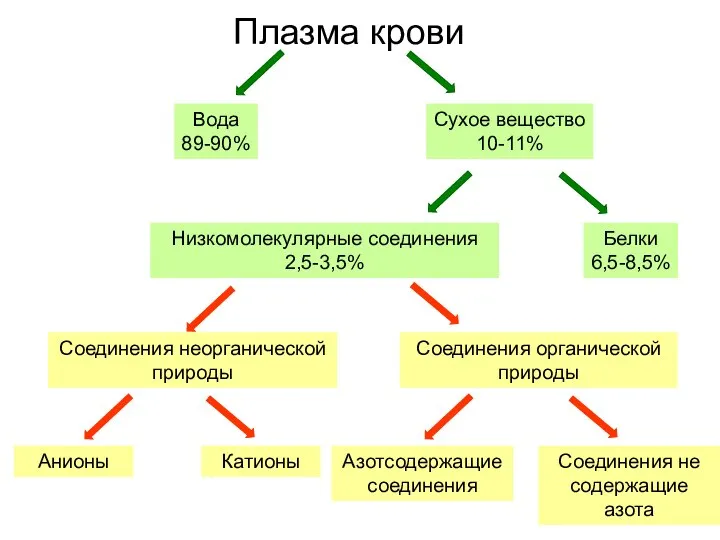 Плазма крови
Плазма крови Корень, его строение и значение
Корень, его строение и значение Отдел Папоротниковидные
Отдел Папоротниковидные Презентация на тему КУДА РАСТЁТ ЧЕЛОВЕЧЕСТВО
Презентация на тему КУДА РАСТЁТ ЧЕЛОВЕЧЕСТВО  Збережемо першоцвіти!
Збережемо першоцвіти! Методическая разработка по выполнению исследовательской работы Фитоиндикация
Методическая разработка по выполнению исследовательской работы Фитоиндикация Многообразие и значение водорослей
Многообразие и значение водорослей Системы органов кровообращения. Строение сердца. Занятие № 25
Системы органов кровообращения. Строение сердца. Занятие № 25 Кровеносная система
Кровеносная система Покриви тіла тварин
Покриви тіла тварин Семейство Бульдоговые летучие мыши (Molossidae)
Семейство Бульдоговые летучие мыши (Molossidae) Какие бывают растения
Какие бывают растения Генетические основы селекции
Генетические основы селекции Анатомия печени
Анатомия печени Происхождение рыб
Происхождение рыб