- Строение клеточных мембран

Содержание
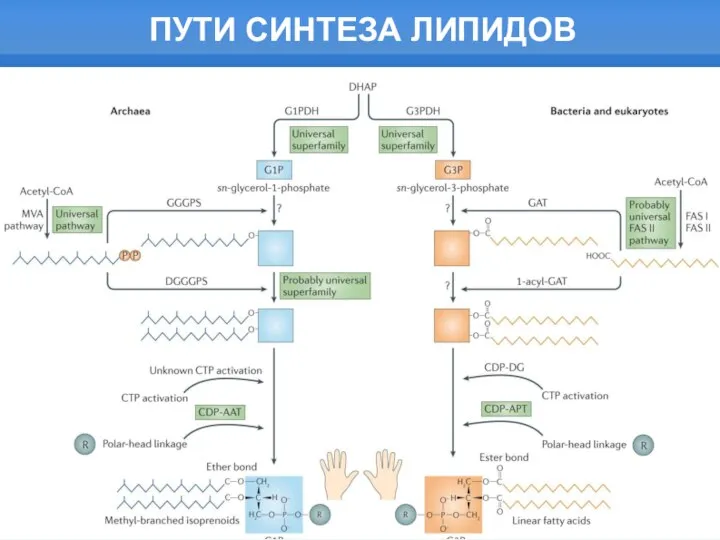
- 2. ПУТИ СИНТЕЗА ЛИПИДОВ
- 3. ПУТИ СИНТЕЗА ТЕРПЕНОВ Филогенетическое распределение мевалонатного пути
- 5. РЕКОНСТРУИРОВАННЫЕ ЛИПИДЫ LUCA Один терпеновый хвост Нет глицерола, терпен прямо соединяется с полярной головой Искусственные мембраны
- 6. РАЗНООБРАЗИЕ И БИОСИНТЕЗ ТЕРПЕНОВ
- 7. МЕМБРАННЫЕ БЕЛКИ Синтез трансмембранных белков обычно требует SRP и транслокон — иначе гидрофобный пептид застревает на
- 8. МЕМБРАННАЯ ЭНЕРГЕТИКА
- 9. СТРУКТУРА МЕМБРАННЫХ АТФаз
- 10. ПРОИСХОЖДЕНИЕ МЕМБРАННЫХ АТФаз
- 11. ПРОИСХОЖДЕНИЕ МЕМБРАННОГО ЭНЕРГЕТИЧЕСКОГО ЦИКЛА
- 12. Изолирующие липиды бактерий и архей
- 13. Больше странных липидов!
- 14. ВОЗМОЖНОСТИ ПРОТОННОЙ И НАТРИЕВОЙ ЭНЕРГЕТИКИ
- 15. Q-ЦИКЛ
- 16. ИСТОРИЯ МЕМБРАН
- 17. ВАРИАНТЫ АНАЭРОБНОГО МЕТАБОЛИЗМА
- 18. СИНТЕЗ ПОРФИРИНОВ Аэробные пути: Металл вставляется в последнюю очередь Анаэробные пути: Металл вставляется на ранних этапах
- 19. СИНТЕЗ ПОРФИРИНОВ: ОБЩАЯ ЧАСТЬ
- 20. СИНТЕЗ ПОРФИРИНОВ: РАЗВЕТВЛЕНИЕ ОТ УРОПОРФИРИНОГЕНА III
- 21. ЭВОЛЮЦИЯ ХИНОНОВ
- 22. ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ КИСЛОРОДНОГО ДЫХАНИЯ
- 23. ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ НИТРАТНОГО ДЫХАНИЯ
- 24. ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ СУЛЬФАТНОГО ДЫХАНИЯ
- 25. МОДУЛЬНАЯ СТРУКТУРА ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫХ КОМПЛЕКСОВ
- 26. ПРОИСХОЖДЕНИЕ КОМПЛЕКСА I (NADH-дегидрогеназа) Простейшие похожие комплексы – гидрогеназы Ech, перенос ферредоксин → H+, c переносом
- 27. ПРОИСХОЖДЕНИЕ КОМПЛЕКСА III (цитохром bc1) Обеспечивает разветвление электронного тока для более полного использования редокс-потенциала сильных окислителей
- 28. ПРОИСХОЖДЕНИЕ КОМПЛЕКСА IV (цитохром-с-оксидаза) 3D-структура комплекса IV указывает на слияние 3 похожих субъединиц в его истории
- 30. Скачать презентацию
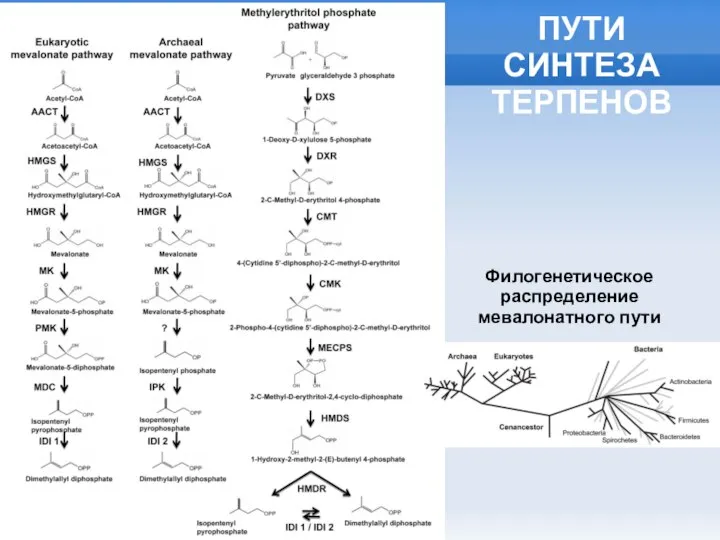
Слайд 3ПУТИ СИНТЕЗА ТЕРПЕНОВ
Филогенетическое распределение
мевалонатного пути
ПУТИ СИНТЕЗА ТЕРПЕНОВ
Филогенетическое распределение
мевалонатного пути
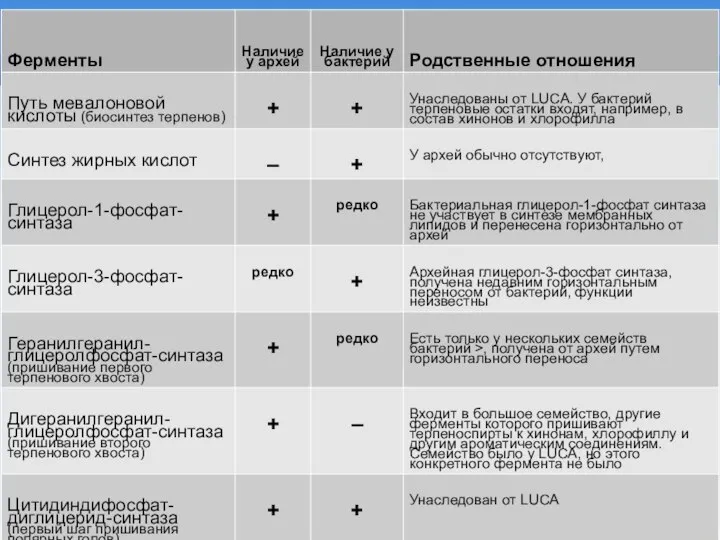
Слайд 5РЕКОНСТРУИРОВАННЫЕ ЛИПИДЫ LUCA
Один терпеновый хвост
Нет глицерола, терпен прямо соединяется с полярной головой
Искусственные
РЕКОНСТРУИРОВАННЫЕ ЛИПИДЫ LUCA
Один терпеновый хвост
Нет глицерола, терпен прямо соединяется с полярной головой
Искусственные

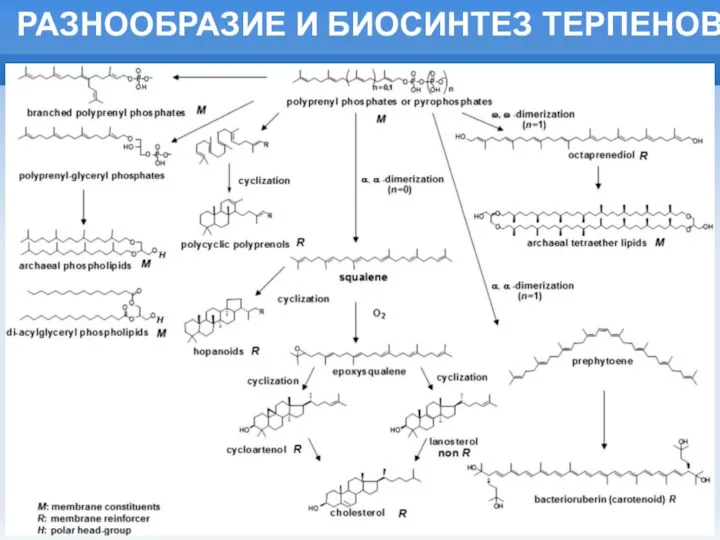
Слайд 6РАЗНООБРАЗИЕ И БИОСИНТЕЗ ТЕРПЕНОВ
РАЗНООБРАЗИЕ И БИОСИНТЕЗ ТЕРПЕНОВ
Слайд 7МЕМБРАННЫЕ БЕЛКИ
Синтез трансмембранных белков обычно требует SRP и транслокон — иначе гидрофобный
МЕМБРАННЫЕ БЕЛКИ
Синтез трансмембранных белков обычно требует SRP и транслокон — иначе гидрофобный
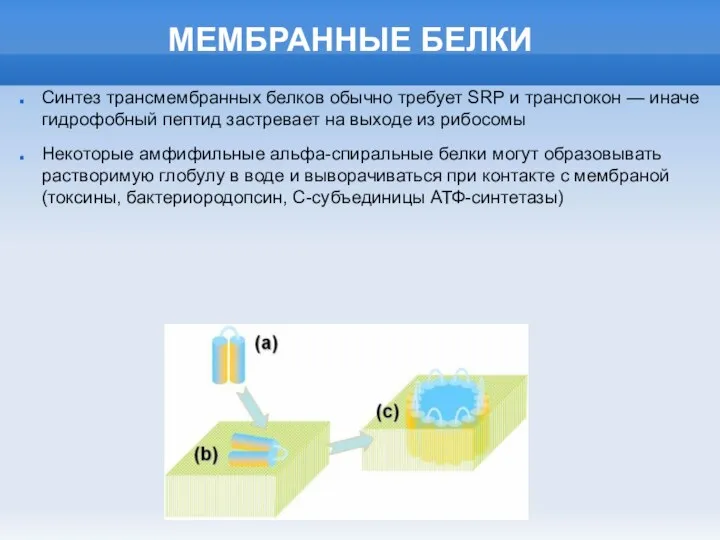
Слайд 8МЕМБРАННАЯ ЭНЕРГЕТИКА
МЕМБРАННАЯ ЭНЕРГЕТИКА
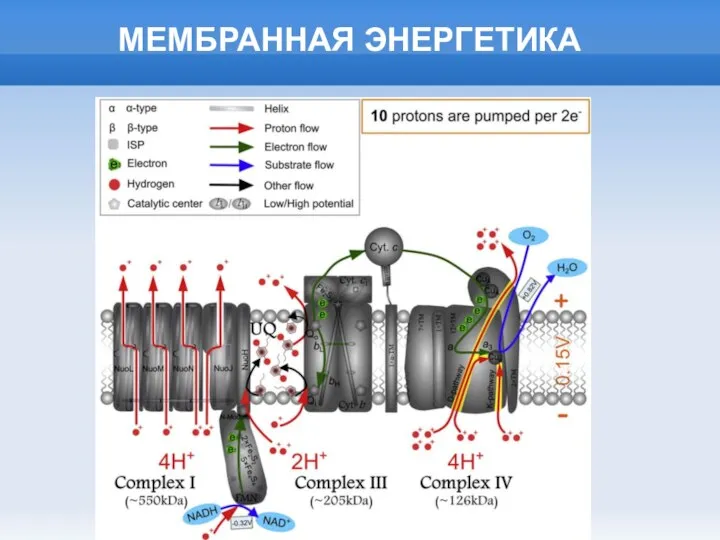
Слайд 9СТРУКТУРА МЕМБРАННЫХ АТФаз
СТРУКТУРА МЕМБРАННЫХ АТФаз
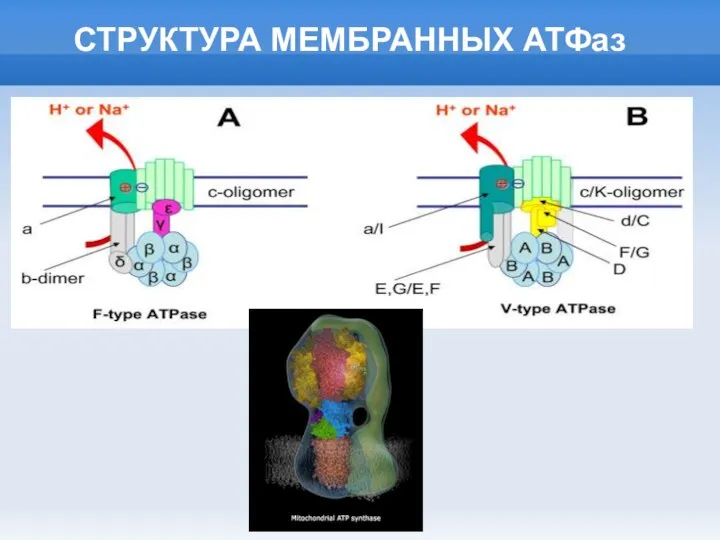
Слайд 10ПРОИСХОЖДЕНИЕ МЕМБРАННЫХ АТФаз
ПРОИСХОЖДЕНИЕ МЕМБРАННЫХ АТФаз
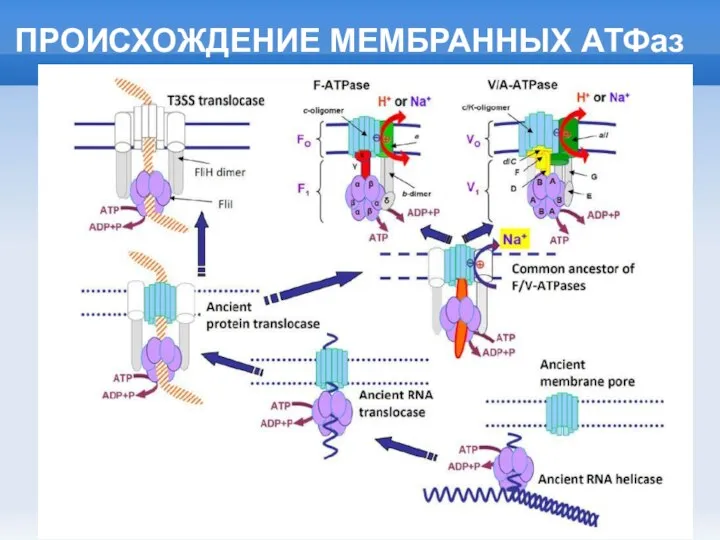
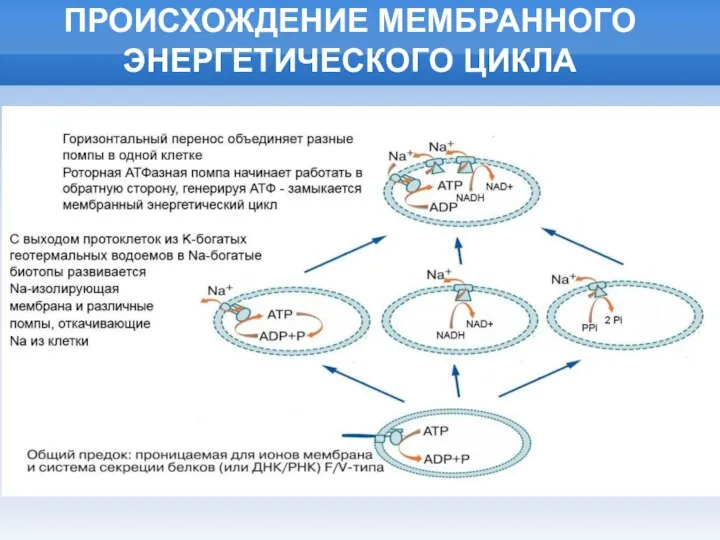
Слайд 11ПРОИСХОЖДЕНИЕ МЕМБРАННОГО ЭНЕРГЕТИЧЕСКОГО ЦИКЛА
ПРОИСХОЖДЕНИЕ МЕМБРАННОГО ЭНЕРГЕТИЧЕСКОГО ЦИКЛА
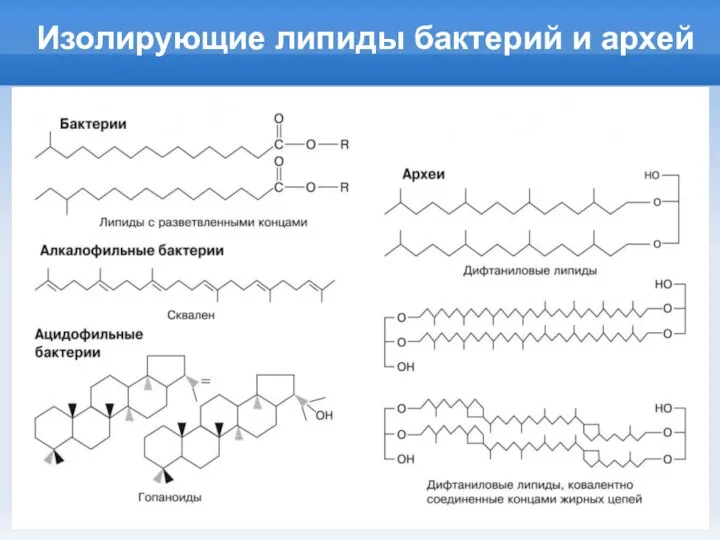
Слайд 12Изолирующие липиды бактерий и архей
Изолирующие липиды бактерий и архей
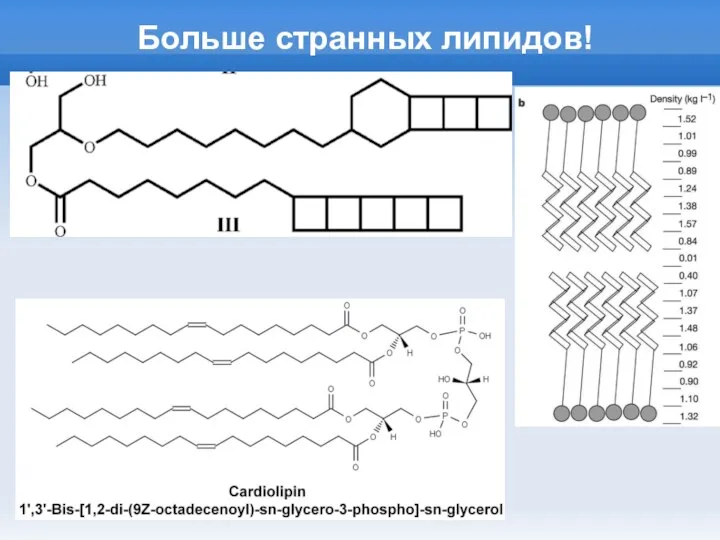
Слайд 13Больше странных липидов!
Больше странных липидов!
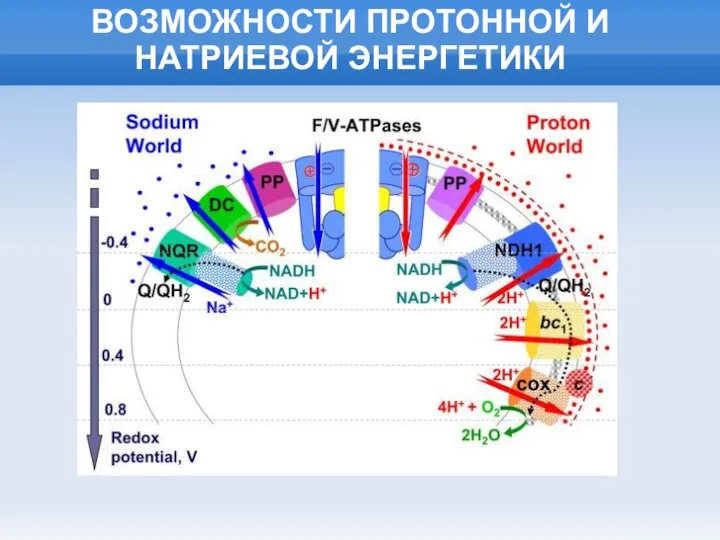
Слайд 14ВОЗМОЖНОСТИ ПРОТОННОЙ И НАТРИЕВОЙ ЭНЕРГЕТИКИ
ВОЗМОЖНОСТИ ПРОТОННОЙ И НАТРИЕВОЙ ЭНЕРГЕТИКИ
Слайд 15Q-ЦИКЛ
Q-ЦИКЛ
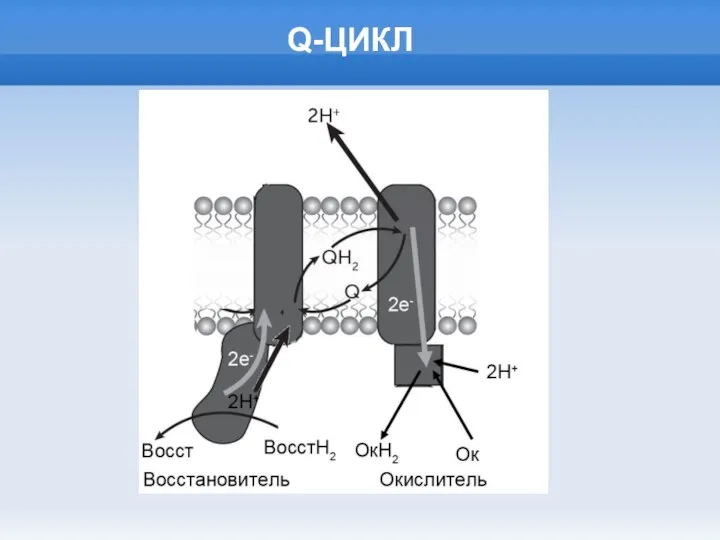
Слайд 16ИСТОРИЯ МЕМБРАН
ИСТОРИЯ МЕМБРАН
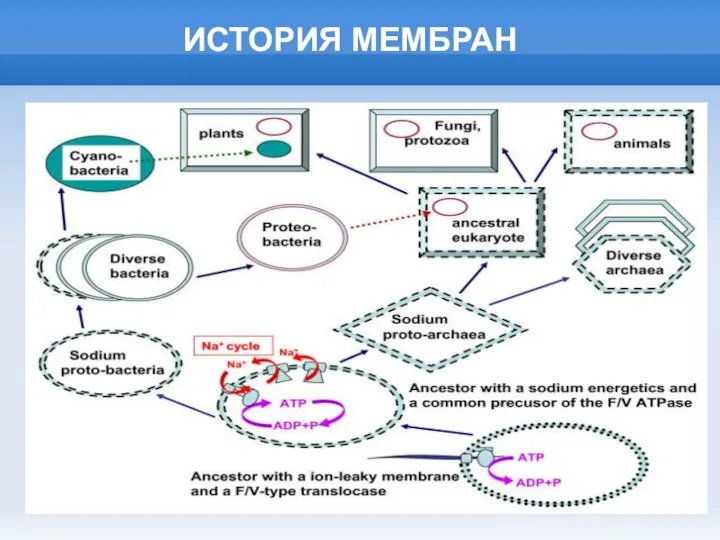
Слайд 17ВАРИАНТЫ АНАЭРОБНОГО МЕТАБОЛИЗМА
ВАРИАНТЫ АНАЭРОБНОГО МЕТАБОЛИЗМА
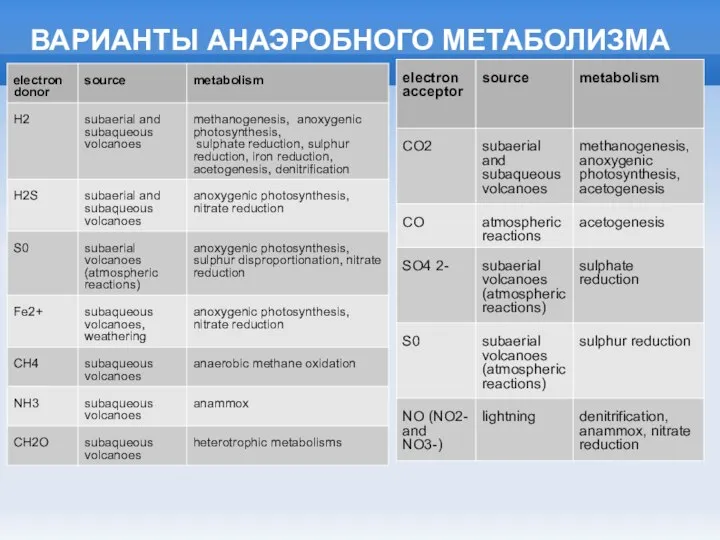
Слайд 18СИНТЕЗ ПОРФИРИНОВ
Аэробные пути:
Металл вставляется в последнюю очередь
Анаэробные пути:
Металл вставляется на ранних этапах
СИНТЕЗ ПОРФИРИНОВ
Аэробные пути:
Металл вставляется в последнюю очередь
Анаэробные пути:
Металл вставляется на ранних этапах
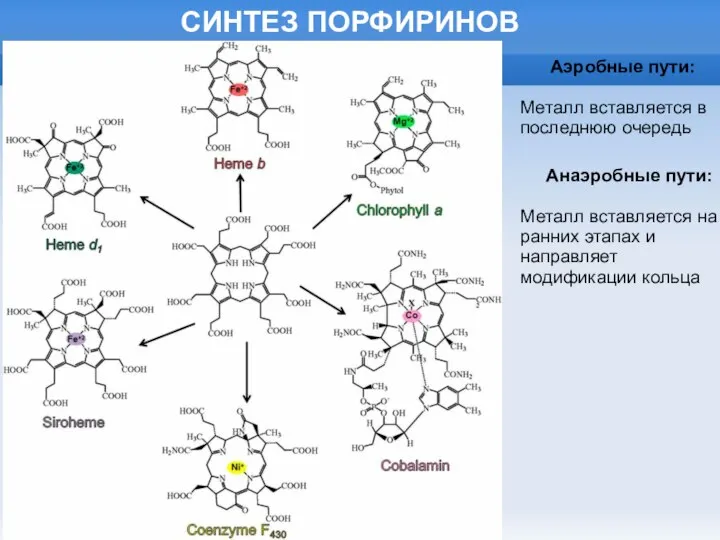
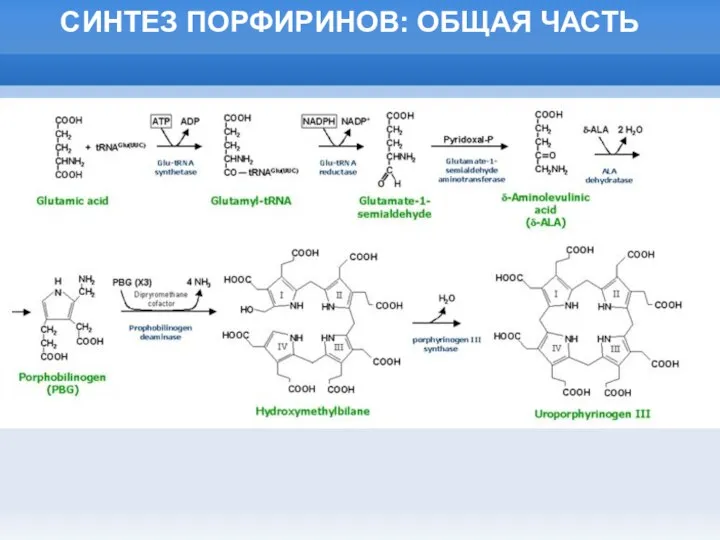
Слайд 19СИНТЕЗ ПОРФИРИНОВ: ОБЩАЯ ЧАСТЬ
СИНТЕЗ ПОРФИРИНОВ: ОБЩАЯ ЧАСТЬ
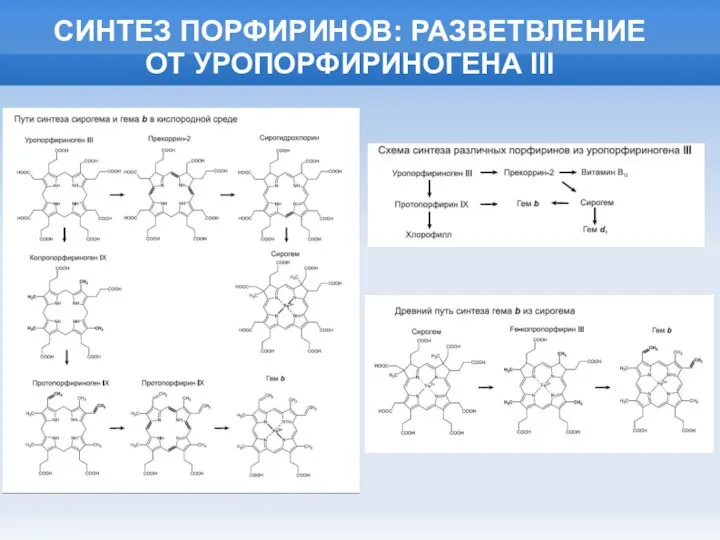
Слайд 20СИНТЕЗ ПОРФИРИНОВ: РАЗВЕТВЛЕНИЕ
ОТ УРОПОРФИРИНОГЕНА III
СИНТЕЗ ПОРФИРИНОВ: РАЗВЕТВЛЕНИЕ
ОТ УРОПОРФИРИНОГЕНА III
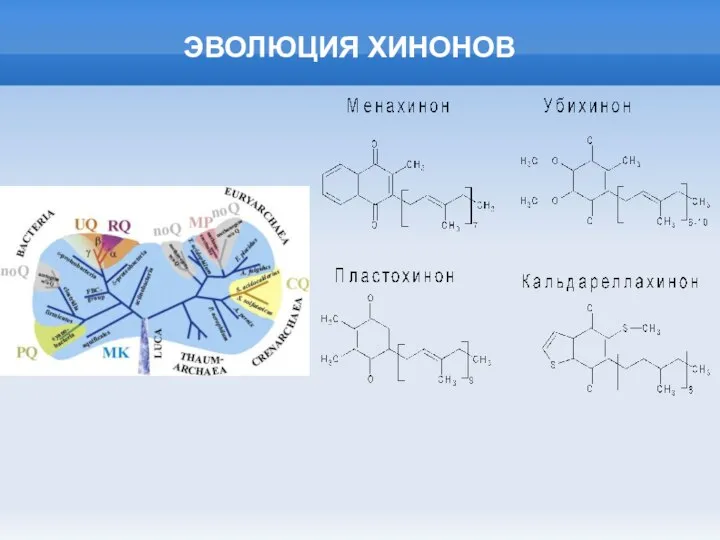
Слайд 21ЭВОЛЮЦИЯ ХИНОНОВ
ЭВОЛЮЦИЯ ХИНОНОВ
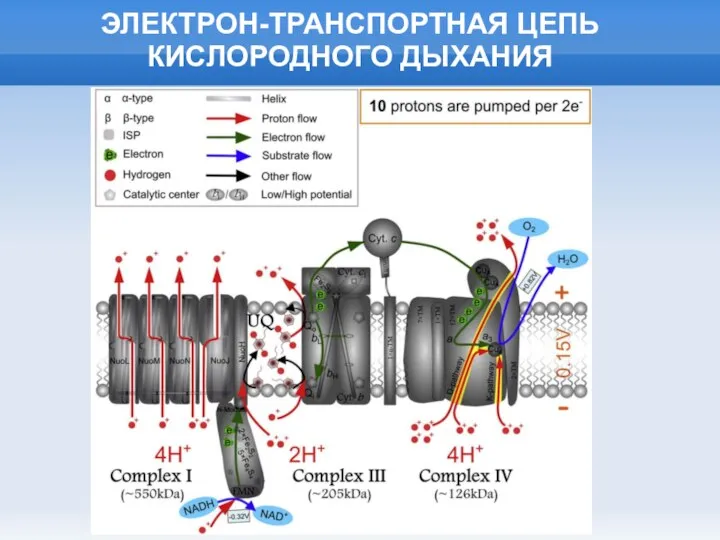
Слайд 22ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ КИСЛОРОДНОГО ДЫХАНИЯ
ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ КИСЛОРОДНОГО ДЫХАНИЯ
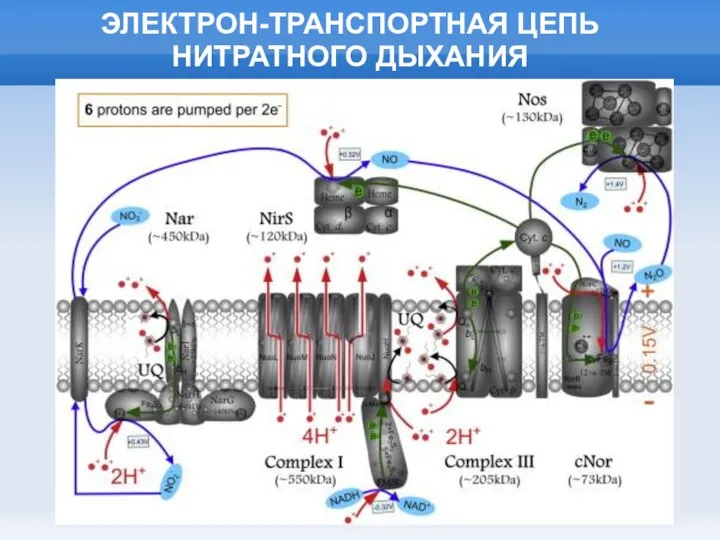
Слайд 23ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ НИТРАТНОГО ДЫХАНИЯ
ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ НИТРАТНОГО ДЫХАНИЯ
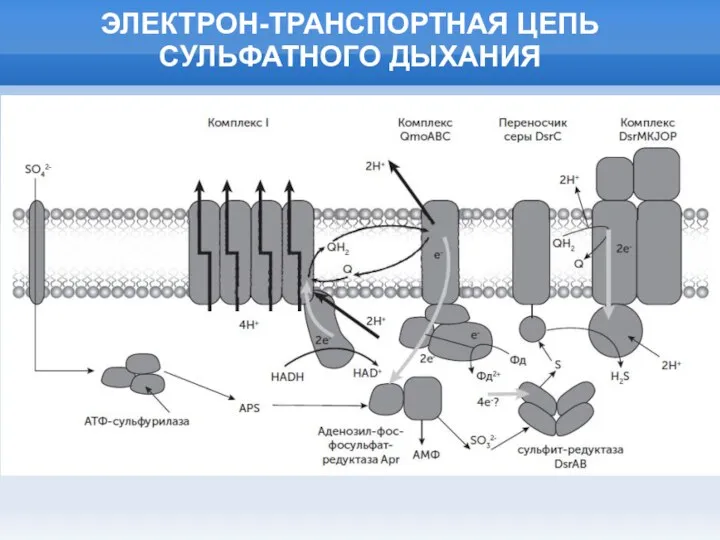
Слайд 24ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ СУЛЬФАТНОГО ДЫХАНИЯ
ЭЛЕКТРОН-ТРАНСПОРТНАЯ ЦЕПЬ СУЛЬФАТНОГО ДЫХАНИЯ
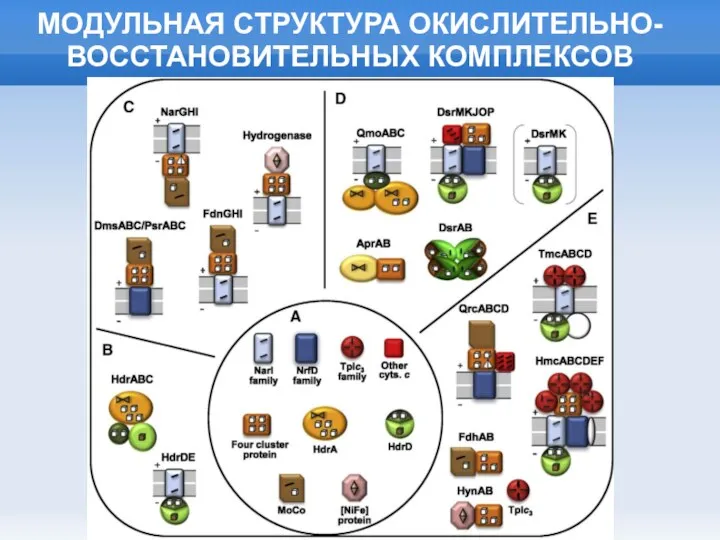
Слайд 25МОДУЛЬНАЯ СТРУКТУРА ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫХ КОМПЛЕКСОВ
МОДУЛЬНАЯ СТРУКТУРА ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫХ КОМПЛЕКСОВ
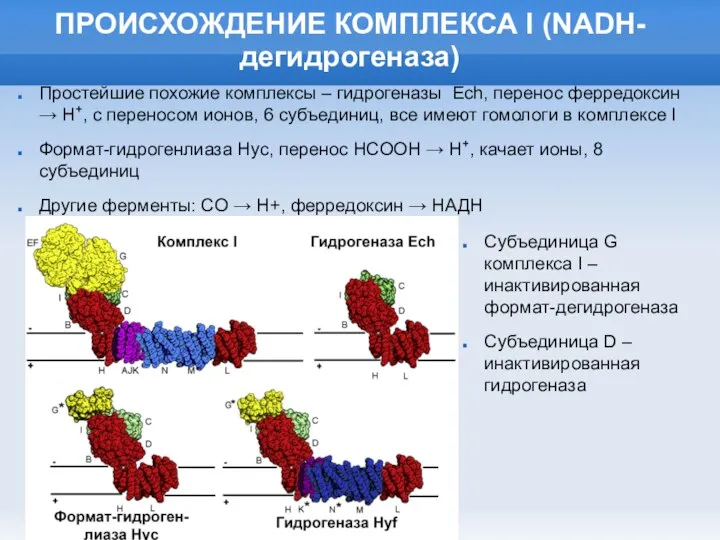
Слайд 26ПРОИСХОЖДЕНИЕ КОМПЛЕКСА I (NADH-дегидрогеназа)
Простейшие похожие комплексы – гидрогеназы Ech, перенос ферредоксин →
ПРОИСХОЖДЕНИЕ КОМПЛЕКСА I (NADH-дегидрогеназа)
Простейшие похожие комплексы – гидрогеназы Ech, перенос ферредоксин →
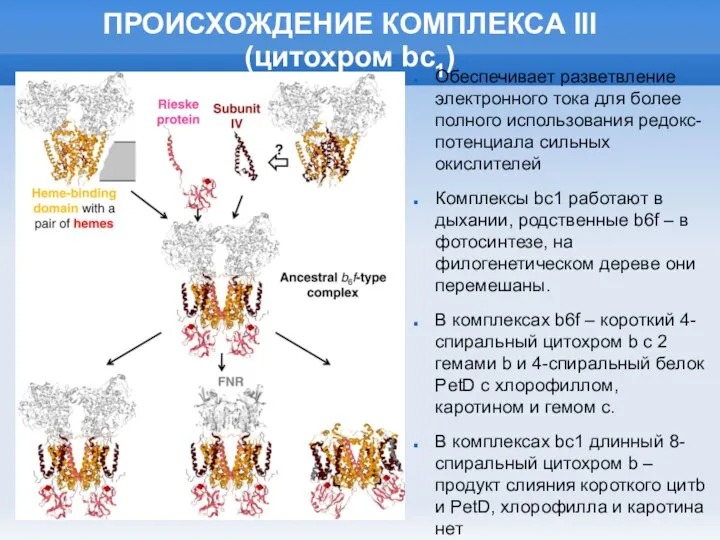
Слайд 27ПРОИСХОЖДЕНИЕ КОМПЛЕКСА III
(цитохром bc1)
Обеспечивает разветвление электронного тока для более полного использования
ПРОИСХОЖДЕНИЕ КОМПЛЕКСА III
(цитохром bc1)
Обеспечивает разветвление электронного тока для более полного использования
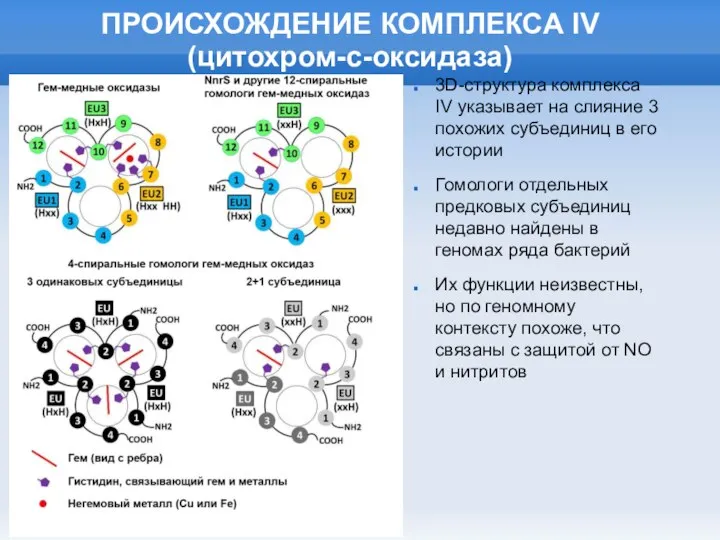
Слайд 28ПРОИСХОЖДЕНИЕ КОМПЛЕКСА IV
(цитохром-с-оксидаза)
3D-структура комплекса IV указывает на слияние 3 похожих субъединиц
ПРОИСХОЖДЕНИЕ КОМПЛЕКСА IV
(цитохром-с-оксидаза)
3D-структура комплекса IV указывает на слияние 3 похожих субъединиц
 Хордовые: строение, происхождение и родственные связи
Хордовые: строение, происхождение и родственные связи Растительные объекты,которые надо знать
Растительные объекты,которые надо знать Теории эволюции
Теории эволюции Зрительный анализатор. Урок 13
Зрительный анализатор. Урок 13 Братья наши меньшие
Братья наши меньшие Забота о потомстве
Забота о потомстве Как мыслит собака
Как мыслит собака Первоцветы села Казьминское
Первоцветы села Казьминское Внутреннее строение листа растения
Внутреннее строение листа растения Онтогенез. Эмбриология – наука о развитии организмов
Онтогенез. Эмбриология – наука о развитии организмов Тип членистоногие. Классы Ракообразные, Паукообразные, Насекомые
Тип членистоногие. Классы Ракообразные, Паукообразные, Насекомые Метаболизм углеводов. Энергетический обмен
Метаболизм углеводов. Энергетический обмен Строение скелета человека. Значение скелета для жизнедеятельности организма
Строение скелета человека. Значение скелета для жизнедеятельности организма Презентация на тему Органические вещества живых организмов
Презентация на тему Органические вещества живых организмов  Митоз бен мейоздың ұқсастықтары мен айырмашылықтары
Митоз бен мейоздың ұқсастықтары мен айырмашылықтары Патофизиология клетки
Патофизиология клетки Половое размножение цветковых растений
Половое размножение цветковых растений Круглые черви. (7)
Круглые черви. (7) Цитоплазма растительной клетки
Цитоплазма растительной клетки Реализация наследственной информации в клетке
Реализация наследственной информации в клетке Жизненный цикл бабочки
Жизненный цикл бабочки Эндокринные железы
Эндокринные железы חיות בעולם
חיות בעולם Анализаторы, строение и функции
Анализаторы, строение и функции Огуречные секреты
Огуречные секреты Виды стебля
Виды стебля Введение в предмет. Уровни организации живого организма
Введение в предмет. Уровни организации живого организма Генетика. Эпигенетические изменения
Генетика. Эпигенетические изменения