- Вегетативная функция ЦНС

Содержание
- 2. Под вегетативной (от лат. vegetare – расти) деятельностью организма понимают работу внутренних органов, которая обеспечивает энергией
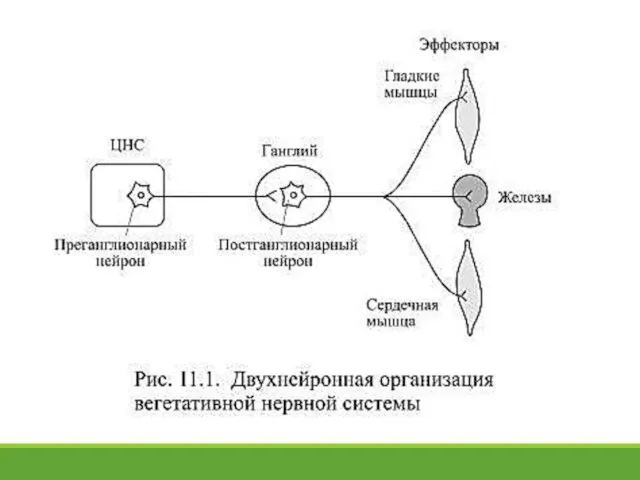
- 3. 11.1. Вегетативная нервная система Вегетативная нервная система иннервирует гладкие мышцы внутренних органов, сердце и внешнесекреторные железы
- 4. В самом конце XIX века британский физиолог Джон Лэнгли (Langley J.) подразделил вегетативную нервную систему на
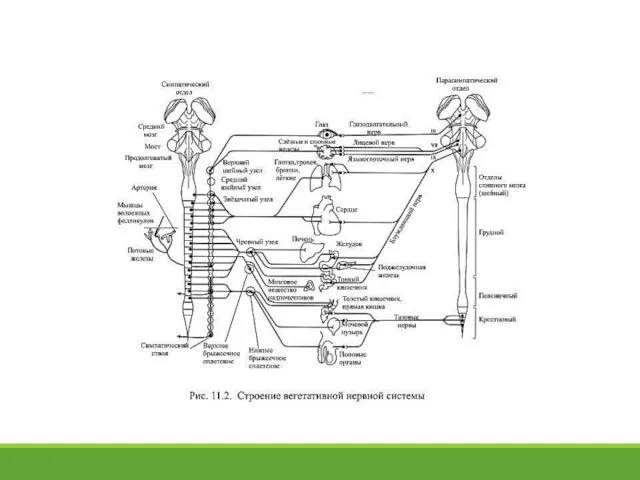
- 5. 11.2. Периферический отдел вегетативной нервной системы Прежде всего необходимо отметить, что периферический отдел вегетативной нервной системы
- 8. Симпатические ганглии принято подразделять на два типа: паравертебральные и превертебральные. Паравертебральные ганглии образуют т.н. симпатические стволы,
- 9. 11.3. Тонус вегетативных нервов Многие вегетативные нейроны обнаруживают фоновую спонтанную активность, т.е. способность самопроизвольно генерировать потенциалы
- 10. Так, например, если по симпатическим нервам, контролирующим гладкие мышцы артерий, проходит 2 импульса в секунду, то
- 11. 11.4. Афферентное звено вегетативных рефлексов Вегетативные реакции можно наблюдать при раздражении почти любой рецептивной области, но
- 12. Интерорецепторы могут принадлежать одному из двух типов чувствительных нейронов. Во-первых, они могут быть чувствительными окончаниями нейронов
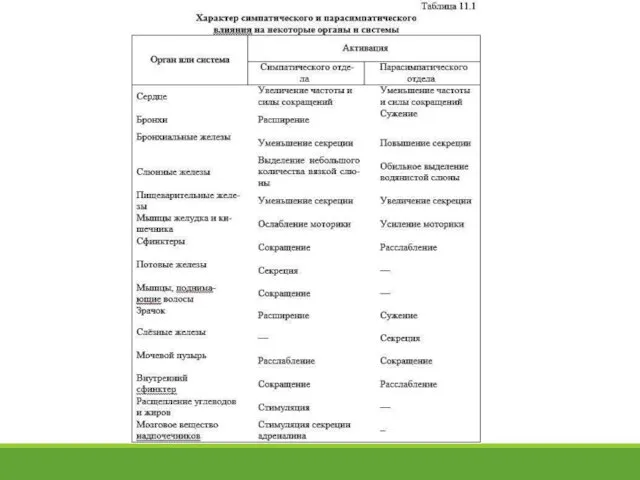
- 13. 11.5. Характер симпатического и парасимпатического влияния на деятельность внутренних органов Большинство органов имеют двойную, т.е. симпатическую
- 14. Влияние двух отделов вегетативной нервной системы на управляемые органы чаще всего противоположно по направленности сдвигов, что
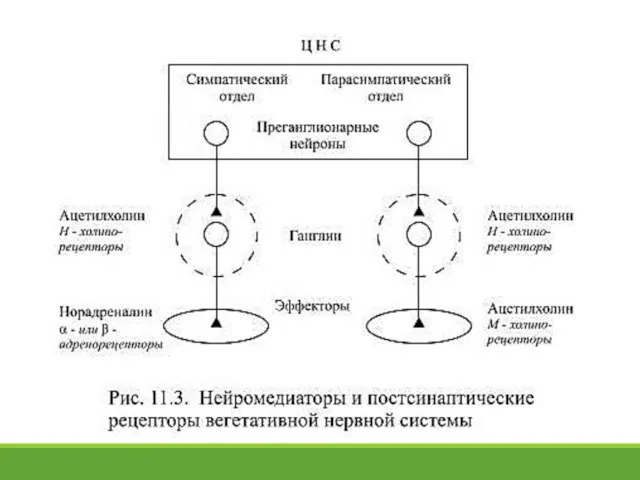
- 16. 11.6. Передача возбуждения в синапсах вегетативной нервной системы В вегетативных ганглиях и симпатического, и парасимпатического отделов
- 18. Согласно другой точке зрения артерии скелетных мышц управляются адренэргическими нейронами, причём норадреналин действует на них через
- 19. Если же концентрация медиатора в синаптической щели становится низкой, с ним взаимодействуют преимущественно b-рецепторы пресинаптической мембраны,
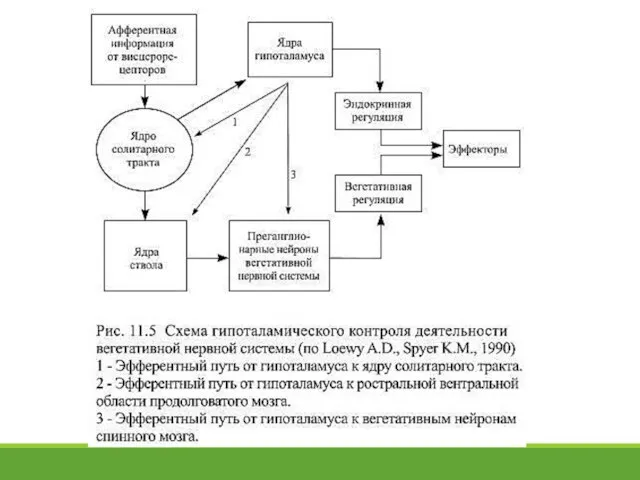
- 20. 11.7. Центры вегетативной регуляции спинного мозга и ствола Многие преганглионарные и постганглионарные нейроны способны активироваться независимо
- 21. Симпатические нейроны, контролирующие потовые железы, сосуды кожи и скелетных мышц располагаются латерально по отношению к нейронам,
- 22. В ретикулярной формации мозгового ствола, где расположены вегетативные центры, используется несколько медиаторных систем, осуществляющих контроль важнейших
- 23. 11. 8. Роль гипоталамуса в регуляции вегетативных функций На гипоталамус приходится менее 1% объёма мозга, однако
- 25. В 40-х годах ХХ века Вальтер Хесс (Hess W.) через введённые с помощью стереотаксической техники электроды
- 26. Следует обратить внимание, что не только уже произошедшие отклонения гомеостаза, но и любое потенциально угрожающее гомеостазу
- 28. 11.9. Вегетативные механизмы регуляции кровообращения В замкнутой сети кровеносных сосудов и сердца (Рис. 11.6) постоянно движется
- 30. Таким образом, главными показателями функции кровообращения оказываются частота и сила сердечных сокращений (от неё зависит систолический
- 31. . В результате уменьшается работа сердца (снижается минутный объём) и понижается тонус сосудов, оказывающих сопротивление кровотоку,
- 32. Одновременно с этим увеличенная по частоте импульсация от хеморецепторов сосудов приведёт к учащению и углублению дыхания
- 33. В результате он начинает забирать из общего потока крови больше, чем забирал прежде, а потому в
- 34. 11.10. Основные звенья регуляции дыхания При спокойном дыхании в лёгкие во время вдоха входит около 300-500
- 36. Очень тонкие стенки альвеол и капилляров не препятствуют газообмену: по градиенту парциальных давлений кислород из альвеолярного
- 37. В самих лёгких, в покрывающей их плевре и в воздухоносных путях есть чувствительные нервные окончания, которые
- 38. Сразу после интенсивной физической работы у человека сохраняется одышка и учащённый пульс, которые прекращаются, когда кислородный
- 39. Резюме Постоянство внутренней среды организма является гарантом его свободной деятельности. Быстрое восстановление смещённых гомеостатических констант осуществляет
- 40. 165. В каком отделе спинного мозга находятся тела парасимпатических нейронов? А. Шейный; Б. Грудной; В. Верхние
- 41. 167. Какие ганглии симпатического отдела следует отнести к паравертебральным? А. Симпатический ствол; Б. Шейный; В. Звёздчатый;
- 42. 169. Что из перечисленного отражает повышение тонуса парасимпатического отдела? А. Расширение зрачков; Б. Расширение бронхов; В.
- 43. 171. Деятельность какой эндокринной железы контролируется симпатическими преганглионарными нейронами? А. Кора надпочечников; Б. Мозговое вещество надпочечников;
- 44. 173. С помощью какого медиатора парасимпатические постганглионарные нейроны обычно действуют на эффекторы? А. Ацетилхолин; Б. Адреналин;
- 45. 175. Какие рецепторы должны связаться с медиатором, чтобы в эффекторной клетке началось повышенное расщепление углеводов? А.
- 46. 177. Какой гомеостатический сдвиг окажет непосредственное действие на центральные рецепторы гипоталамуса? А. Повышение артериального давления; Б.
- 47. 179. Где расположены барорецепторы, поставляющие информацию вегетативным центрам продолговатого мозга, осуществляющим регуляцию работы сердца и артериального
- 48. 181. Какое событие скорее всего произойдёт вследствие повышения напряжения углекислого газа в крови? А. Уменьшение частоты
- 50. Скачать презентацию
Слайд 2Под вегетативной (от лат. vegetare – расти) деятельностью организма понимают работу внутренних
Под вегетативной (от лат. vegetare – расти) деятельностью организма понимают работу внутренних

Слайд 311.1. Вегетативная нервная система
Вегетативная нервная система иннервирует гладкие мышцы внутренних органов,
11.1. Вегетативная нервная система
Вегетативная нервная система иннервирует гладкие мышцы внутренних органов,

Слайд 4 В самом конце XIX века британский физиолог Джон Лэнгли (Langley J.)
В самом конце XIX века британский физиолог Джон Лэнгли (Langley J.)

Слайд 511.2. Периферический отдел вегетативной нервной системы
Прежде всего необходимо отметить, что периферический
11.2. Периферический отдел вегетативной нервной системы
Прежде всего необходимо отметить, что периферический

Слайд 8Симпатические ганглии принято подразделять на два типа: паравертебральные и превертебральные. Паравертебральные ганглии
Симпатические ганглии принято подразделять на два типа: паравертебральные и превертебральные. Паравертебральные ганглии

Слайд 911.3. Тонус вегетативных нервов
Многие вегетативные нейроны обнаруживают фоновую спонтанную активность, т.е. способность
11.3. Тонус вегетативных нервов
Многие вегетативные нейроны обнаруживают фоновую спонтанную активность, т.е. способность

Слайд 10Так, например, если по симпатическим нервам, контролирующим гладкие мышцы артерий, проходит 2
Так, например, если по симпатическим нервам, контролирующим гладкие мышцы артерий, проходит 2

Слайд 1111.4. Афферентное звено вегетативных рефлексов
Вегетативные реакции можно наблюдать при раздражении почти любой
11.4. Афферентное звено вегетативных рефлексов
Вегетативные реакции можно наблюдать при раздражении почти любой

Слайд 12Интерорецепторы могут принадлежать одному из двух типов чувствительных нейронов.
Во-первых, они могут
Интерорецепторы могут принадлежать одному из двух типов чувствительных нейронов.
Во-первых, они могут

Слайд 1311.5. Характер симпатического и парасимпатического влияния на деятельность внутренних органов
Большинство органов
11.5. Характер симпатического и парасимпатического влияния на деятельность внутренних органов
Большинство органов

Слайд 14Влияние двух отделов вегетативной нервной системы на управляемые органы чаще всего противоположно
Влияние двух отделов вегетативной нервной системы на управляемые органы чаще всего противоположно

Слайд 1611.6. Передача возбуждения в синапсах вегетативной нервной системы
В вегетативных ганглиях и
11.6. Передача возбуждения в синапсах вегетативной нервной системы
В вегетативных ганглиях и

Слайд 18Согласно другой точке зрения артерии скелетных мышц управляются адренэргическими нейронами, причём норадреналин
Согласно другой точке зрения артерии скелетных мышц управляются адренэргическими нейронами, причём норадреналин

Слайд 19 Если же концентрация медиатора в синаптической щели становится низкой, с ним
Если же концентрация медиатора в синаптической щели становится низкой, с ним

Слайд 2011.7. Центры вегетативной регуляции спинного мозга и ствола
Многие преганглионарные и постганглионарные
11.7. Центры вегетативной регуляции спинного мозга и ствола
Многие преганглионарные и постганглионарные

Слайд 21Симпатические нейроны, контролирующие потовые железы, сосуды кожи и скелетных мышц располагаются латерально
Симпатические нейроны, контролирующие потовые железы, сосуды кожи и скелетных мышц располагаются латерально

Слайд 22В ретикулярной формации мозгового ствола, где расположены вегетативные центры, используется несколько медиаторных
В ретикулярной формации мозгового ствола, где расположены вегетативные центры, используется несколько медиаторных

Слайд 2311. 8. Роль гипоталамуса в регуляции вегетативных функций
На гипоталамус приходится менее 1%
11. 8. Роль гипоталамуса в регуляции вегетативных функций
На гипоталамус приходится менее 1%

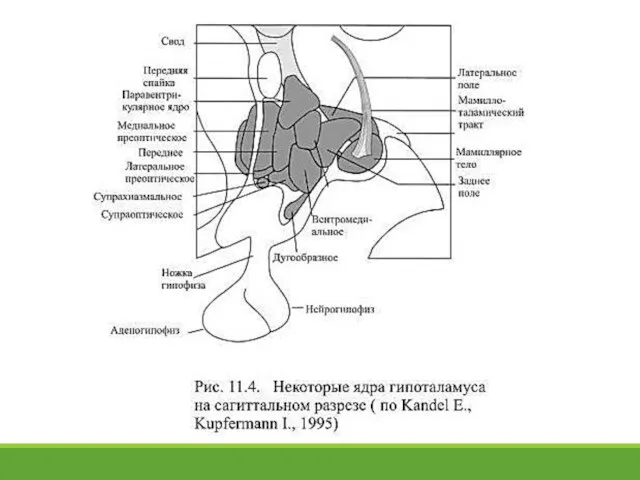
Слайд 25В 40-х годах ХХ века Вальтер Хесс (Hess W.) через введённые с
В 40-х годах ХХ века Вальтер Хесс (Hess W.) через введённые с

Слайд 26Следует обратить внимание, что не только уже произошедшие отклонения гомеостаза, но и
Следует обратить внимание, что не только уже произошедшие отклонения гомеостаза, но и

Слайд 2811.9. Вегетативные механизмы регуляции кровообращения
В замкнутой сети кровеносных сосудов и сердца (Рис.
11.9. Вегетативные механизмы регуляции кровообращения
В замкнутой сети кровеносных сосудов и сердца (Рис.


Слайд 30Таким образом, главными показателями функции кровообращения оказываются частота и сила сердечных сокращений
Таким образом, главными показателями функции кровообращения оказываются частота и сила сердечных сокращений

Слайд 31. В результате уменьшается работа сердца (снижается минутный объём) и понижается тонус
. В результате уменьшается работа сердца (снижается минутный объём) и понижается тонус

Слайд 32Одновременно с этим увеличенная по частоте импульсация от хеморецепторов сосудов приведёт к
Одновременно с этим увеличенная по частоте импульсация от хеморецепторов сосудов приведёт к

Слайд 33В результате он начинает забирать из общего потока крови больше, чем забирал
В результате он начинает забирать из общего потока крови больше, чем забирал

Слайд 3411.10. Основные звенья регуляции дыхания
При спокойном дыхании в лёгкие во время вдоха
11.10. Основные звенья регуляции дыхания
При спокойном дыхании в лёгкие во время вдоха


Слайд 36Очень тонкие стенки альвеол и капилляров не препятствуют газообмену: по градиенту парциальных
Очень тонкие стенки альвеол и капилляров не препятствуют газообмену: по градиенту парциальных

Слайд 37В самих лёгких, в покрывающей их плевре и в воздухоносных путях есть
В самих лёгких, в покрывающей их плевре и в воздухоносных путях есть

Слайд 38 Сразу после интенсивной физической работы у человека сохраняется одышка и учащённый
Сразу после интенсивной физической работы у человека сохраняется одышка и учащённый

Слайд 39Резюме
Постоянство внутренней среды организма является гарантом его свободной деятельности.
Быстрое восстановление смещённых
Резюме
Постоянство внутренней среды организма является гарантом его свободной деятельности.
Быстрое восстановление смещённых

Слайд 40165. В каком отделе спинного мозга находятся тела парасимпатических нейронов?
А. Шейный;
165. В каком отделе спинного мозга находятся тела парасимпатических нейронов?
А. Шейный;

Слайд 41167. Какие ганглии симпатического отдела следует отнести к паравертебральным?
А. Симпатический ствол;
167. Какие ганглии симпатического отдела следует отнести к паравертебральным?
А. Симпатический ствол;

Слайд 42169. Что из перечисленного отражает повышение тонуса парасимпатического отдела?
А. Расширение зрачков;
169. Что из перечисленного отражает повышение тонуса парасимпатического отдела?
А. Расширение зрачков;

Слайд 43171. Деятельность какой эндокринной железы контролируется симпатическими преганглионарными нейронами?
А. Кора надпочечников;
171. Деятельность какой эндокринной железы контролируется симпатическими преганглионарными нейронами?
А. Кора надпочечников;

Слайд 44173. С помощью какого медиатора парасимпатические постганглионарные нейроны обычно действуют на эффекторы?
173. С помощью какого медиатора парасимпатические постганглионарные нейроны обычно действуют на эффекторы?

Слайд 45175. Какие рецепторы должны связаться с медиатором, чтобы в эффекторной клетке началось
175. Какие рецепторы должны связаться с медиатором, чтобы в эффекторной клетке началось

Слайд 46177. Какой гомеостатический сдвиг окажет непосредственное действие на центральные рецепторы гипоталамуса?
А.
177. Какой гомеостатический сдвиг окажет непосредственное действие на центральные рецепторы гипоталамуса?
А.

Слайд 47179. Где расположены барорецепторы, поставляющие информацию вегетативным центрам продолговатого мозга, осуществляющим регуляцию
179. Где расположены барорецепторы, поставляющие информацию вегетативным центрам продолговатого мозга, осуществляющим регуляцию

Слайд 48181. Какое событие скорее всего произойдёт вследствие повышения напряжения углекислого газа в
181. Какое событие скорее всего произойдёт вследствие повышения напряжения углекислого газа в

 Функциональная анатомия спинного мозга
Функциональная анатомия спинного мозга Нервная система
Нервная система Jesień w lesie
Jesień w lesie Питание, увлажнение кожи. Защита кожи
Питание, увлажнение кожи. Защита кожи Жизненные формы растений
Жизненные формы растений Екі факторлы дисперсиялық талдау
Екі факторлы дисперсиялық талдау Схемы скрещивания. Генетика. 10 класс
Схемы скрещивания. Генетика. 10 класс Животные нашего края
Животные нашего края Undersea world
Undersea world Типы развития насекомых
Типы развития насекомых Презентация на тему ГЕНЕТИКА ПОЛА
Презентация на тему ГЕНЕТИКА ПОЛА  Цветы: колокольчик, бубенчик, букашник, кольник
Цветы: колокольчик, бубенчик, букашник, кольник ЦНС (центральная нервная система)
ЦНС (центральная нервная система) Животные дома
Животные дома Презентация на тему Биогеоценоз
Презентация на тему Биогеоценоз  Практическая работа по биологии. Черенкование растений
Практическая работа по биологии. Черенкование растений Какие бывают жуки
Какие бывают жуки Зоопланктон. Типы зоопланктона
Зоопланктон. Типы зоопланктона Растения как объект технологии
Растения как объект технологии Животные, обитающие на Кавказе
Животные, обитающие на Кавказе Обмен углеводов. Переваривание и всасывание углеводов у детей. Метаболизм гликогена
Обмен углеводов. Переваривание и всасывание углеводов у детей. Метаболизм гликогена Нервная регуляция автономных функций
Нервная регуляция автономных функций Презентация на тему Клеточное строение растений
Презентация на тему Клеточное строение растений  Мышечная ткань
Мышечная ткань Презентация на тему Грегор Иоган Мендель и его исследования
Презентация на тему Грегор Иоган Мендель и его исследования  Путешествие во времени, учебное занятие. Эволюция органического мира
Путешествие во времени, учебное занятие. Эволюция органического мира Размножение и развитие растений. Опыление
Размножение и развитие растений. Опыление Мероприятие для 7-9-х классов Северный Ледовитый океан: экология, история, современность
Мероприятие для 7-9-х классов Северный Ледовитый океан: экология, история, современность