Слайд 2«Не удивительно ли, что столь огромное существо, как кашалот, видит мир таким

крошечным глазом и слышит громы ухом, которое куда меньше заячьего?»
(Герман Мелвилл, "Моби Дик")
Слайд 3Обитание - открытая вода и около берега.
Статус вида - распространенный.
Численность групп -

1-50 (1-150) большими группами могут путешествовать вместе.
Расположение спинного плавника - далеко позади центра.
Вес новорожденного - 1 тонна.
Вес взрослого - 20-50 тонн.
Длина новорожденного - 3,5-4,5 м.
Длина взрослого - 11-18 м.
Питание - ведущее место занимают головоногие моллюски, среди них почти 20 видов кальмаров, в том числе гигантские архитеутисы длиной более 10 м, и несколько видов осьминогов. Из рыб поедают, преимущественно в северных водах, морских окуней, алепизавров, корифен, скатов, мелких акул, тресковых, глубоководных рыб - макрурид и удильщиков, а в водах Антарктики - клыкачей. Кальмары, защищаясь от кашалота, оставляют на его голове следы своих присосок в виде вдавленных кругов диаметром до 20 см.
Слайд 5Самый крупный представитель подотряда зубатых китов. Резко выражен половой диморфизм.
Размер самцов

в среднем около 15 м, максимальный до 20 м. Размер самок в среднем 11 м, максимальный до 13 м.
У самцов тело толстое с громадной, до трети длины тела, квадратной головой. У самок тело стройнее, голова занимает не более 1/4 тела, спереди более округлая.
Дыхало смещено в левый передний угол головы и по форме напоминает латинскую букву S. За исключением головы, тело выглядит морщинистым.
Окраска от темно-серой до черной, однотонная, с мелкой пятнистостью.
Спинной плавник имеет вид бугра, позади которого располагаются еще несколько небольших бугров.
Фонтан направлен вперед и влево от конца головы под углом 45°.
Слайд 7Кашалот — один из наиболее многочисленных видов китов. Обычен на всем его

громадном ареале.
С целью коммерческого промысла добывался до 1978 г. Самая мощная популяция, вероятно, обитает в Тихом океане.
Предположительная численность кашалотов северной части Тихого океана определяется приблизительно в 150 тыс. голов (Нисиваки, 1966).
У берегов Чили и Перу в некоторые годы добывали до 4 - 5 тыс. кашалотов, что говорит о большой численности этих китов и в южной половине Тихого океана, причем они многочисленны и в других водах Южного полушария. Так, в некоторые годы в Антарктике добывалось до 6—7 тыс. только самцов кашалотов. Что касается Северной Атлантики, то для этой части Мирового океана отсутствуют даже ориентировочные данные о численности.
Вероятно, эта популяция относительно небольшая, о чем можно судить и по данным сравнительно небольшого промысла. Наибольшее количество кашалотов добывается в водах Азорских о-вов, где общая добыча никогда не превышала 1000 голов в год.
Слайд 10Основу пищи кашалотов и в Северном и в Южном полушариях составляют 2

группы животных — головоногие моллюски и рыбы, причем первые занимают явно доминирующее положение. Наиболее подробно питание кашалотов исследовано в северной половине Тихого океана. Данные из остальных районов Мирового океана отрывочны и неполны.
В желудках кашалотов из вод Курильских о-вов (360 желудков) определено 28 видов головоногих моллюсков (21 кальмар и 7 осьминогов).
Головоногие составляют около 95% (по весу) потребляемой кашалотами пищи, рыбы — менее 5%, хотя они и были обнаружены почти у 1/3 исследованных кашалотов.
Из головоногих основное значение имеют кальмары; осьминоги составляют не более 4% съеденной пищи. В то же время кормовое значение практически имеют только 7 видов кальмаров: Gomttis migister, G.fabricii.
Слайд 11G. simile, Taonius pava, Galliteufhis armata, Chiroteuthis veranyi и Male-agroteuthis separata, составляющие
до 80% поедаемых головоногих ,причем 60% приходится на долю 3 видов семейства Gonatidae. Многочисленные в дальневосточных морях каракатицы совершенно не встречаются в желудках кашалотов (Бетешева и Акимушкин, 1955; Акимушкин, 1957; Бетешева, 1960, 1961).
Примечательно, что наиболее многочисленные в дальневосточных водах кальмары Ommatostrephes sloanei-pacificus, обитающие в верхних горизонтах моря, не имеют почти никакого значения в питании кашалотов, а живущие близ поверхности очень многочисленные каракатицы совершенно не поедаются этими китами.
Слайд 12Основная пища состоит из батипелагических видов головоногих моллюсков и рыб, большинство из
которых не поднимается на глубину менее 500 м, а некоторые из них обитают на глубине от 1000 м и ниже (Акимушкин, 1954).
Следовательно, кашалот ловит свою добычу на глубине не менее 300—400 м, где у него почти нет пищевых конкурентов, за исключением быть может китов семейства клюворыловых (Ziphiidae). В поисках пищи кашалот может опускаться до самого дна, так как в желудках иногда находили донных глубоководных животных.
Относительно часто в желудках кашалотов находят совершенно посторонние предметы: резиновые сапоги, мотки проволоки, стеклянные рыболовные поплавки, резиновые перчатки, детские пластмассовые игрушки (автомашина, пистолет, кукла, ведерко и др.), кувшины, пластмассовые сумки, кокосовые орехи, хлорвиниловые мешочки, рыболовные снасти с крючками, пустые бутылки, яблоки и многое другое (Берзин, 1971).
Слайд 14Суточная активность, поведение.
Кашалоты — полигамы, половой диморфизм в размерах у них

ярко выражен. В теплых водах они образуют группы, состоящие обычно из 10—15 самок и одного крупного самца. Нередко формируются группы в сотни голов. Взрослые самцы, не входящие в состав групп (гаремов), держатся отдельно, чаще одиночно, лишь иногда в местах скопления пищи они собираются в относительно большие группы.
Новейшие наблюдения показывают, что в гаремах держатся не самые крупные старые самцы, а более молодые длиной 13—14 м, с большей половой активностью, которые и являются вожаками гаремов.
Они изгоняют старых самцов, и те держатся отдельно от гаремов. Все это заставляет детально пересмотреть современные представления о гаремах, их составе, поведении, взаимоотношении отдельных возрастных и половых групп в мелких и крупных скоплениях и другие стороны семейного и стадного поведения кашалотов (Берзин, 1971; В. А. Земский, Д. Д. Тормосов, Ю. А. Михалев).
Слайд 15В районах летнего обитания самцы кашалотов, в зависимости от возраста и размера

животных, в большинстве случаев образуют группировки определенного состава. Группы состоят чаще из сравнительно одноразмерных животных, чем из разноразмерных. Из 23 исследованных в заливе Аляска группировок 18 (78,3%) состояли из близких по размерам китов и только 5 (21,7%) — из разноразмерных.
Полагают, что причина объединения в группы животных, близких по размерам, заключается в одинаковой возможности добывания пищи, находящейся на разных глубинах. Крупные киты могут нырять глубже и кормиться в таких местах, в которых более мелкие не могут добывать себе корм.
Одноразмерные группы более мелких китов обычно состоят из почти одновозрастных животных (разница в возрасте 1—2 года). В группах, состоящих из более крупных китов, разница в возрасте больше (иногда 10 лет). Вероятно, это определяется большей индивидуальной вариацией размеров взрослых китов, у которых прекратился рост (Тарасович, 1967).
Слайд 16Характер ныряния кашалотов своеобразен; они могут пробыть под водой до 1 ч,

а иногда и больше (предполагают, что максимально 2 ч).
Перед длительным погружением кашалот ныряет резко, тело его круто изгибается, на поверхности моря появляется горб, затем изогнутый хвостовой стебель, зверь ныряет почти вертикально и в большинстве случаев показывает над водой лопасти хвостового плавника.
Появление над морем такой «бабочки» служит верным признаком, что кит ушел надолго, причем вынырнуть он может очень далеко от места погружения,
Слайд 17Максимальная глубина погружения кашалотов не установлена.
Известно 19 случаев повреждения подводных кабелей

запутавшимися в них кашалотами, причем исключительно в тропических и умеренных водах (между 46° ю. ш. и 46° с. ш.).
Глубина обнаружения мертвых кашалотов была от 118 до 1116 м. Запутавшись в кабеле, кашалот хватает его зубами, нарушая изоляцию, что и вызывает повреждения.
В 1951 г. у кабеля, соединяющего Лиссабон с Малагой, было обнаружено повреждение такого же типа на глубине 2200 м — это пока максимальная известная глубина.
Слайд 18В большинстве случаев глубина повреждений кабеля превышала 500 м, несколько раз —

1000 м. Чаще всего кабель был обмотан вокруг нижней челюсти и хвостового плавника кашалотов.
Полагают, что кит захватывает кабель в пасть во время погони за пищей, затем начинает биться и наматывает на себя слабину кабеля.
Предполагалось, что кашалоты запутываются на малых глубинах и лишь позже постепенно соскальзывают по склону морского дна.
Однако характер опутывания свидетельствует, что это происходит именно на тех местах, где их находят (Хизен, 1957; Яблоков, 1962). Вероятно, кашалоты действительно способны опускаться на глубину 1000 м и более.
Слайд 19После длительного пребывания под водой кашалот долго отдыхает на поверхности. В это

время он лежит почти на одном месте, лишь немного продвигаясь вперед, и, находясь в горизонтальном положении, ритмично погружается в воду, пуская фонтан каждые 10—15 сек.
Количество фонтанов различно и, вероятно, в значительной степени зависит от времени, проведенного под водой. Обычно бывает фонтанов 20—40, в отдельных случаях до 75.
Молодые животные дают меньше фонтанов, чем старые, которые, очевидно, ныряют глубже и на более долгий срок.
Во время этого своеобразного отдыха кашалоты очень спокойны и беспечны, лежат на поверхности по 10 мин и более, так что китобойное судно в большинстве случаев успевает подойти к замеченному киту на верный выстрел.
Во время промысла в Антарктике отмечены случаи, когда китобоец подходил к кашалоту в буквальном смысле вплотную, ударяясь форштевнем (В.А. Арсеньев).
Слайд 20Иногда поднимающийся с глубины кашалот выходит из воды почти вертикально, высовываясь чуть

ли не до половины туловища.
В других случаях зверь высоко выпрыгивает под углом к поверхности моря и затем падает на воду с громадным всплеском.
Слайд 21Скорость движения кашалотов сравнительно мала. Так называемые ходовые киты (переходящие из одного

района в другой), или мигрирующие, идут обычно по прямому курсу со скоростью 5—7 миль в час.
Они плывут близко от поверхности воды, часто и неглубоко ныряют, дают много фонтанов. Пасущиеся животные передвигаются медленного скоростью, вероятно, не более 3 миль. В поисках корма они надолго заныривают и при этом постоянно и очень резко изменяют направление движения под водой. Напуганный или раненый кашалот идет со всей возможной скоростью чаще всего по прямому курсу и часто дает фонтаны. Максимальная скорость движения крупного кашалота вряд ли превышает 10 миль в час.
Во всяком случае корабли со скоростью хода 12 — 12,5 миль всегда настигают стремящегося уйти кашалота через 40—50 мин погони.
Слайд 22Раненый гарпуном кашалот резко идет в глубину, но довольно скоро снова выходит

на поверхность, поднимаясь почти вертикально и выставляя над водой свою громадную тупую голову.
Если своевременно не остановить ход китобойца, то выныривающий кит оказывается под корпусом двигающегося по инерции судна и может сильно ударить головой в днище.
Известны случаи, когда в этой ситуации кит выводил из строя китобоец, погнув лопасти винта или даже гребной вал.
Эти случаи послужили основанием для широко распространенного утверждения, что раненый кашалот часто активно нападает на корабль.
Однако это, безусловно, не умышленное нападение, а чисто случайные удары, объясняющиеся своеобразием выныривания кашалотов (В.А. Арсеньев).
Слайд 23Относительно нередки случаи выбрасывания кашалотов на берег как одиночками, так и группами.

Это отмечено на берегах Дании, Франции, Флориды, Калифорнии, Тасмании и Новой Зеландии.
В 8 случаях зарегистрировано выбрасывание групп китов от 16 до 36 голов.
Иногда такие группы состоят только из самок, но чаще из одних самцов.
Выбросившиеся 16 января 1954 г. на побережье Калифорнии 22 самца были приблизительно одинакового размера (Гильмор, 1957; Томилин, 1962).
Слайд 25Характер и пути миграций кашалотов изучены далеко недостаточно.
Схема миграций такова: в
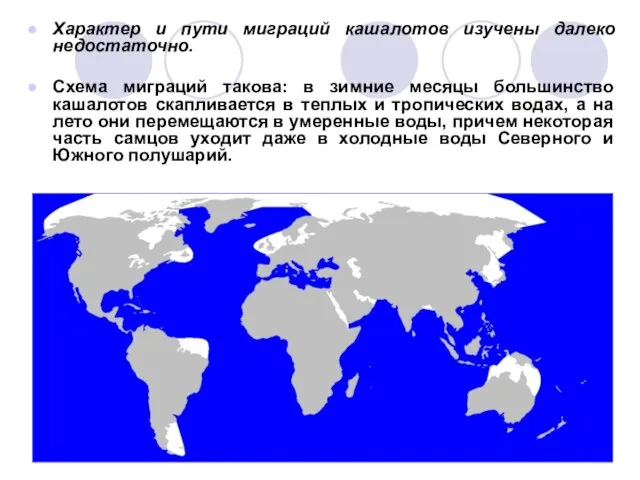
зимние месяцы большинство кашалотов скапливается в теплых и тропических водах, а на лето они перемещаются в умеренные воды, причем некоторая часть самцов уходит даже в холодные воды Северного и Южного полушарий.
Слайд 26Основной фактор, определяющий пути миграций и дальность продвижения кашалотов в холодные воды,
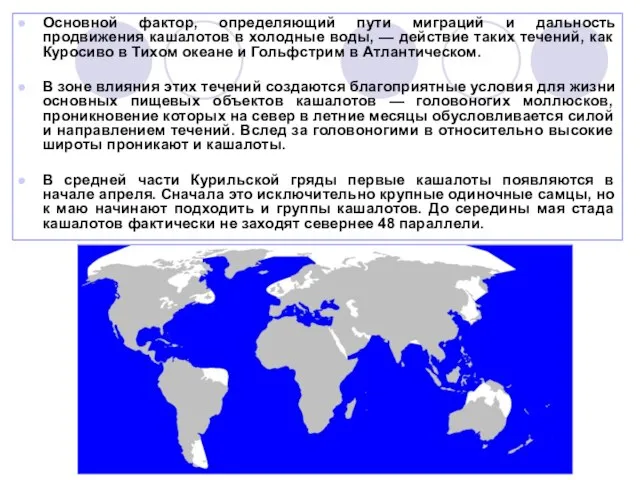
— действие таких течений, как Куросиво в Тихом океане и Гольфстрим в Атлантическом.
В зоне влияния этих течений создаются благоприятные условия для жизни основных пищевых объектов кашалотов — головоногих моллюсков, проникновение которых на север в летние месяцы обусловливается силой и направлением течений. Вслед за головоногими в относительно высокие широты проникают и кашалоты.
В средней части Курильской гряды первые кашалоты появляются в начале апреля. Сначала это исключительно крупные одиночные самцы, но к маю начинают подходить и группы кашалотов. До середины мая стада кашалотов фактически не заходят севернее 48 параллели.
Слайд 27В северной части Курильской гряды в конце апреля — начале мая появляются

одиночные кашалоты-самцы, в конце мая начинается подход так называемых гаремных китов, а к середине июня здесь образуются устойчивые скопления кашалотов различного пола и возраста.
Такие скопления называют косячными. Крупные самцы в это время в большинстве уходят дальше к северу (Тарасевич, 1965).
В первой половине июня смешанные группы кашалотов довольно равномерно распределяются с тихоокеанской стороны средней и северной части Курильской гряды, причем скопления их отодвигаются дальше от берегов.
Взрослые одиночные самцы в это время наблюдаются в Охотском море, куда совершенно не заходят самки и молодые животные. В июле количество кашалотов у Курильских о-вов значительно уменьшается, и по всему району наблюдаются лишь небольшие группы.
Можно предположить, что в это время стада кашалотов частью проходят к юго-восточной Камчатке, а частично рассеиваются на большой акватории.
Слайд 28В конце июля в средней части Курильской гряды (пролив Буссоль — о-в

Симушир) наблюдается подход с юга новых стад кашалотов среднего размера, состоящих в основном из молодых самцов.
В самом конце июля в районе Уруп — Симушир опять появляются новые косяки, в которых уже много самок; среди них встречаются крупные самцы. Это явно вновь подошедшие гаремы.
В течение двух первых декад августа кашалоты держатся в том же районе и даже несколько распространяются к югу (до пролива Фриза), совершенно не продвигаясь к северу.
В это время косяки состоят из очень мелких животных, однако в конце августа здесь снова начинают попадаться крупные киты.
Вероятно, в это время они уже начинают двигаться из Охотского моря к югу.
Слайд 29Такой характер подходов подтверждает предположение о том, что в летнее время у

Курильских о-вов обитает 2 узколокальных стада кашалотов — южнокурильское и северокурильское. Киты южнокурильского стада летом обитают у северной части Японских о-вов и южной части Курильской гряды, достигая на севере ее средней части.
Северокурильское стадо населяет воды северных Курильских о-вов, держится у побережья юго-восточной Камчатки и может достигать вод Командорских о-вов (С. К. Клумов).
В сентябре в курильских водах наблюдается наибольшее количество кашалотов, так как в это время начинается подход китов, возвращающихся из северных вод. Октябрь — месяц массового ухода кашалотов на юг, и стада их у Курильских о-вов постепенно редеют.
Отдельные животные или даже небольшие группы в благоприятные годы могут оставаться на зиму вблизи островов Курильской гряды (Слепцов, 1955).
Слайд 30Северной границей распространения взрослых самок и молодых кашалотов обоего пола принимается 51°

с.ш. (Омура, 1950), хотя их встречали и севернее — до 52—54° с.ш. — в Кроноцком и Камчатском заливах, у Командорских о-вов (Кирпичников, 19506; Слепцов, 1950; Томилин, 1957).
Следовательно, в благоприятные годы самки могут доходить и до 53—54° с. ш.
Обычной границей распространения взрослых самцов считают 61—62° с. ш. (мыс Наварин), куда могут подходить киты как от западного, так и от восточного побережий Тихого океана. Более вероятно, что северной границей распространения самцов азиатского стада служит район Олюторского залива (60° с.ш.), хотя при благоприятных условиях отдельные киты могут достигать 65°30' с.ш. (Омура, 1950).
Слайд 32Самки кашалотов обычно не выходят за пределы теплых и умеренных вод, поэтому

сезон спаривания и деторождения у них ограничен не так резко, как у тех видов, самки которых совершают регулярные миграции в холодные воды обоих полушарий.
Роды у кашалотов бывают в течение всего года, однако наибольшее количество самок приносит потомстве все же в относительно сжатые сроки.
Для Северного полушария это происходит, по-видимому, в раннеосенние месяцы. Так, в Северной Атлантике наибольшее количество деторождении отмечается с мая по ноябрь с максимумом с июля по сентябрь (Кларк, 1956).
Слайд 33Перед наступлением родов самки концентрируются в затишных зонах, где условия наиболее благоприятны

для новорожденных.
К таким районам в Тихом океане относятся воды Маршальских и Бонинских о-вов, восточного побережья Японии, в меньшей степени — воды Южных Курильских о-вов и Галапагосских о-вов, в Атлантическом океане — воды Азорских, Бермудских о-вов, побережья африканской провинции Наталь и Мадагаскара.
Кашалоты выбирают районы с чистой глубокой водой, расположенные с подветренной стороны островов или рифов (Слепцов, 1955; Томилин, 1957).
Слайд 34В Южном полушарии щенка протекает в период с декабря по апрель с

максимумом в феврале. Роды происходят в районах со спокойными и относительно теплыми водами, там, где мало акул, касаток и других врагов, опасных для новорожденных.
В апреле 1962 г. вблизи о-вов Тристан-да-Кунья с вертолета (Ф. Хомчик) наблюдали процесс рождения детеныша. Среди нескольких групп кашалотов численностью по 25—30 голов каждая выделялась одна, состоящая из 6 кашалотов. Киты непрерывно ныряли рядом друг с другом, поэтому вода казалась мутной.
Скоро вода окрасилась кровью, а среди разошедшихся китов на поверхности воды был виден новорожденный, который сразу же поплыл рядом с матерью. Их сопровождали 4 других кашалота, вероятно, тоже самки.
Слайд 35Наблюдатели свидетельствуют, что во время родов самка держится в вертикальном положении, высунувшись

из воды почти на 1/4 длины тела. У новорожденного лопасти хвостового плавника некоторое время свернуты трубочкой. В это время другие взрослые самки ныряют под новорожденного, помогая ему держаться на поверхности моря (Тормосов, 1970).
Беременность у кашалотов гораздо продолжительнее, чем у большинства других видов китообразных. Полагают, что она длится 16—18 месяцев (Метьюз, 19386; Нисиваки, Хибия и Осуми, 1958; Осуми, 1965). Соотношение самцов и самок у эмбрионов близко 1:1.
Самка обычно приносит одного детеныша, двойни очень редки (0,66%; Кларк, 1956). Периодика размножения определяется в 3 (Кларк, 1956).
Слайд 37Продолжительность эмбрионального развития кашалотов достигает 16 (быть может и 18) месяцев.
Поэтому

рост эмбрионов протекает медленнее, чем у других видов китов, с более коротким периодом эмбрионального развития.
Длина новорожденных детенышей колеблется от 350 до 500 см (Томилин, 1957).
Вес новорожденных может достигать 1000 кг.
Продолжительность периода молочного кормления, по аналогии с другими видами крупных китов, определялась в 6—7 месяцев (Виноградов, 1949), а по новейшим данным — в 24—25 месяцев (Осуми, 1965).
Слайд 38Возраст кашалотов определяют по количеству слоев в дентине, которые образуются в результате

сезонных изменений характера обмена веществ, связанных с изменением условий обитания.
Вплоть до заполнения полости пульпы ежегодно откладывается 2 слоя дентина — более широкий и светлый, сильнее кальцинированный, и более узкий и темный, менее кальцинированный.
Эти 2 слоя образуют одно годовое кольцо, количество которых и показывает возраст кита (Нисиваки, Хибия и Осуми, 1958; Берзин, 1961).
Вопрос о количестве слоев, откладывающихся в течение одного года, все еще не находит единодушия. Возможно, годовых колец может быть два или одно годовое кольцо состоит из иного количества промежуточных колец. Изучение этого вопроса продолжается.
Слайд 39К концу первого года жизни кашалоты северной части Тихого океана достигают длины

6 м или несколько больше, т. е. прирост длины тела за первый год жизни составляет почти 2 м (все цифры средние).
Затем темп роста несколько замедляется, к трем годам животные достигают длины 8 м, вырастая за два года еще на 2 м.
В возрасте 3,5—4 лет самки кашалотов достигают половой зрелости, и темп их роста становится значительно медленнее, чем у самцов, — ежегодный прирост не превышает 50 см.
Наступление физической зрелости и прекращение роста у самок происходит в возрасте 15 лет при длине тела 11 м.
Самка длиной 11,7 м имела возраст более 30 лет, средние пары зубов у нее уже выпали.
Слайд 40Приближенно можно считать, что, к предельному возрасту самки достигают длины 12 м

или немногим больше.
Самцы достигают половой зрелости в 5 лет при длине тела 9,5 м, но темп роста у них в первое время после достижения половозрелости не снижается.
В течение нескольких последующих лет годовой прирост составляет в среднем около 80 см и лишь позже постепенно уменьшается.
Прекращение роста отмечается в возрасте 23—25 лет при длине тела около 16 м (Берзин, 1961, 1964а). Длина тела самцов антарктических кашалотов в период достижения половой зрелости равна 11,5—12,5м, что определяется общими более крупными размерами китов Южного полушария (Метьюз, 19386; Нисиваки, 1955; см. «Географическая изменчивость»).
Полагают, что у животных старше 35—40 лет возраст определить невозможно, так как полость пульпы полностью закрывается. Однако такой возраст не предельный для кашалотов.
Слайд 42Громадная угловатая голова занимает более 1/3 длины тела.
Дыхательное отверстие у кашалотов

расположено на конце морды с левой стороны, а не на верхней точке головы, как у всех других китов, поэтому фонтан его не вертикальный, а направлен вперед и влево под углом примерно 45° к линии горизонта. Фонтан широкий, «пушистый», высотой не более 3—4м.
По характеру фонтана кашалот может быть опознан издали безошибочно.
Окраска однотонная, обычно темная.
При нырянии на глубину часто показывает лопасти хвостового плавника. После долгого погружения лежит на поверхности воды почти без движения, подпуская к себе корабль почти вплотную.
Слайд 441. Википедия
2. floranimal.ru/pages/animal/k/211.html
3. sakhalin.ru/boomerang/sea/kit%20zub10.htm
4. cetacea.ru/sperm.htm
Список литературы:












 Безопасность IoT устройств
Безопасность IoT устройств Тема 26. Теория. Демеркуризация
Тема 26. Теория. Демеркуризация Театр. Тест
Театр. Тест ВОЛЬФА 8срс
ВОЛЬФА 8срс Внеклассное мероприятие
Внеклассное мероприятие Влияние природных факторов на человека
Влияние природных факторов на человека Житие протопоп Аввакум
Житие протопоп Аввакум Жилищное право
Жилищное право Конституция России - путь к правовому государству
Конституция России - путь к правовому государству Управление Активами и пассивами
Управление Активами и пассивами Подвижные игры зимой
Подвижные игры зимой Студенческий cовет Ижевского промышленно-экономического колледжа
Студенческий cовет Ижевского промышленно-экономического колледжа В стране Наречий
В стране Наречий Порядок допуска к медицинской деятельности. Сертификация специалистов. Трудоустройство.
Порядок допуска к медицинской деятельности. Сертификация специалистов. Трудоустройство. Моя профессия - военный
Моя профессия - военный Приглашаем на-военную службу по контракту
Приглашаем на-военную службу по контракту Рекламный спрос на женскую аудиторию в социальных сетях. - презентация
Рекламный спрос на женскую аудиторию в социальных сетях. - презентация Решение тригонометрических уравнений
Решение тригонометрических уравнений Презентация на тему Появление товарного производства. Складывание торговых путей. Установление торговых связей Руси с другими го
Презентация на тему Появление товарного производства. Складывание торговых путей. Установление торговых связей Руси с другими го Влияние коннотаций положительных ценностей в рекламной фотографии на выбор товара потребителями
Влияние коннотаций положительных ценностей в рекламной фотографии на выбор товара потребителями Экологические праздники
Экологические праздники BPS
BPS Нормативно-правовое обеспечение инклюзивного образования
Нормативно-правовое обеспечение инклюзивного образования Бесплатное продвижение возможно, или внутренняя оптимизация сайта.
Бесплатное продвижение возможно, или внутренняя оптимизация сайта. Искусство XX века. Изобразительное искусство модерна
Искусство XX века. Изобразительное искусство модерна «Нам дан во владение самый богатый, меткий, могучий и поистине волшебный русский язык» К.Г.Паустовский
«Нам дан во владение самый богатый, меткий, могучий и поистине волшебный русский язык» К.Г.Паустовский Презентация на тему Параллелограмм. Свойства параллелограмма
Презентация на тему Параллелограмм. Свойства параллелограмма