- Альтернативный сплайсинг – детская площадка эволюции

Содержание
- 2. Оценки % альтернативно сплайсируемых генов млекопитающих (по году публикации) Человек (выборка из генома) Человек (полные хромосомы)
- 3. Значение альтернативного сплайсинга Функциональное: поддержание белкового разнообразия Человек: ~30.000 генов, >100.000 белков поддержание белкового единообразия Например,
- 4. Эволюция альтернативной экзон-интронной структуры млекопитающие: человек, мышь, собака двукрылые насекомые: Drosophila melanogaster, D. pseudoobscura, Anopheles gambiae
- 5. Элементарные альтернативы Кассетный экзон Альтернативный донорный сайт Альтернативный акцепторный сайт И т.д.: взаимоисключающие экзоны, удержанные интроны,
- 6. EDAS: альтернативный сплайсинг генов человека 20809 генов; 114568 мРНК; 91835 белков; 51713 альтернатив, из них 31746
- 7. Альтернативная экзон-интроная структура генов млекопитающих Тройки ортологичных генов: человек-мышь-собака Следим за судьбой (консервативностью) альтернатив человека в
- 8. Потеря альтернативы в геноме мыши общий предок
- 9. Потеря альтернативы в геноме собаки (хотя теоретически возможно возникновение в общем предке приматов и грызунов) общий
- 10. Появление альтернативы в геноме человека (или ошибка сплайсинга, или экспериментальный шум) Common ancestor
- 11. Неконсервативные альтернативы человека: шум? Консервативные альтернативы Альтернативы в генах человека, отсутствующие в генах мыши – реальны
- 12. Human-specific alternatives: noise? Conserved alternatives Добавим геном собаки Консервативные альтернативы потери у собаки потери у мыши
- 13. Наблюдения Часто вставляемые экзоны консервативны независимо от того, сбивают ли они рамку Редко вставляемые экзоны менее
- 14. Консервативность белок-кодирующих областей в генах насекомых Технически сложнее (сложности с выравниванием), но наблюдения те же: альтернативные
- 15. Консервативность элементарных альтернатив D.melanogaster в генах D. pseudoobscura голубой – сохранились точно зеленый – вставка интрона
- 16. Консервативность элементарных альтернатив D.melanogaster в генах Anopheles gambiae голубой – сохранились точно зеленый – вставка интрона
- 17. Скорость эволюции и тип отбора в альтернативных и константных областях Пары ортологичных генов человек и мышь
- 18. Гены человека и мыши: несимметричная гистограмма dn/ds(const)–dn/ds(alt) Черный: «тень» левой части. В большей части генов dn/ds(alt)
- 19. Гены Drosophila: слабее отбор в альтернативных областях? Больше замен в альт. обл. Равный уровень замен Больше
- 20. Альтернативные области изменяются быстрее, чем константные dN dN/dS dS dN/dS dS dN 1 0
- 21. Ослабление стабилизирующего отбора в альтернативных областях dN/dS dN dS dN/dS dS dN 1 0
- 22. Положительный отбор во внутренних альтернативных областях генов Drosophila dN/dS dS dN dN(AI)/dS(AI) = 1,43 1,5 0
- 23. Тест МакДональза-Крейтмана: положительный отбор в минорных альтернативных областях Сравнение различий между генами человека и шимпанзе со
- 24. Попытка синтеза Альтернативный сплайсинг часто видоспецифичен «молодые» альтернативные изоформы часто тканеспецифичны … но все же функционален
- 25. Планы: много дрозофил; млекопитающие (ENCODE)
- 26. Что делать? Оценить не только скорость потерь альтернатив, но и скорость приобретений (отличая молодые изоформы от
- 27. Благодарности Обсуждения Евгений Кунин (NCBI) Игорь Рогозин (NCBI) Всеволод Макеев (ГосНИИГенетика) Дмитрий Петров (Stanford) Дмитрий Фришман
- 28. Авторы Андрей Миронов (МГУ, ИППИ) Рамиль Нуртдинов (МГУ) – человек/мышь/собака Дмитрий Малько (ГосНИИГенетика) – дрозофилы/комар Екатерина
- 29. Основные публикации 10,1 D.B.Malko, V.J.Makeev, A.A.Mironov, M.S.Gelfand (2006) Evolution of the exon-intron structure and alternative splicing
- 31. Скачать презентацию
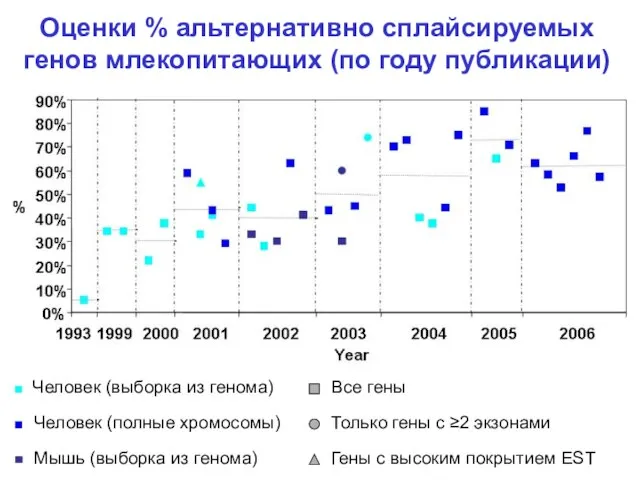
Слайд 2Оценки % альтернативно сплайсируемых генов млекопитающих (по году публикации)
Человек (выборка из генома)
Человек
Оценки % альтернативно сплайсируемых генов млекопитающих (по году публикации)
Человек (выборка из генома)
Человек
Слайд 3Значение альтернативного сплайсинга
Функциональное:
поддержание белкового разнообразия
Человек: ~30.000 генов, >100.000 белков
поддержание белкового единообразия
Например,
Значение альтернативного сплайсинга
Функциональное:
поддержание белкового разнообразия
Человек: ~30.000 генов, >100.000 белков
поддержание белкового единообразия
Например,

Слайд 4Эволюция альтернативной экзон-интронной структуры
млекопитающие: человек, мышь, собака
двукрылые насекомые: Drosophila melanogaster, D.
Эволюция альтернативной экзон-интронной структуры
млекопитающие: человек, мышь, собака
двукрылые насекомые: Drosophila melanogaster, D.

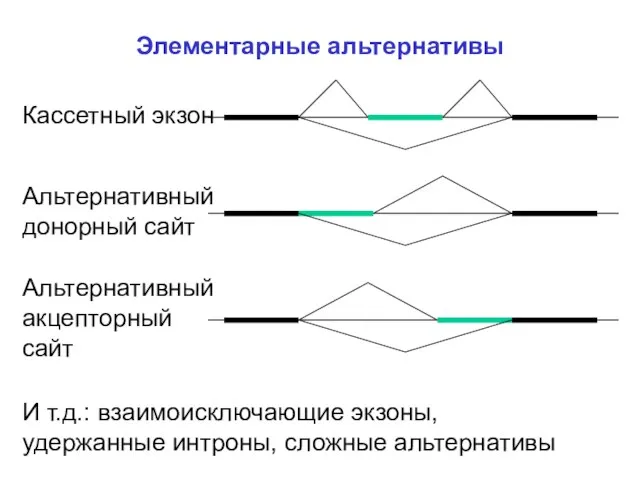
Слайд 5Элементарные альтернативы
Кассетный экзон
Альтернативный донорный сайт
Альтернативный акцепторный сайт
И т.д.: взаимоисключающие экзоны,
удержанные интроны,
Элементарные альтернативы
Кассетный экзон
Альтернативный донорный сайт
Альтернативный акцепторный сайт
И т.д.: взаимоисключающие экзоны, удержанные интроны,
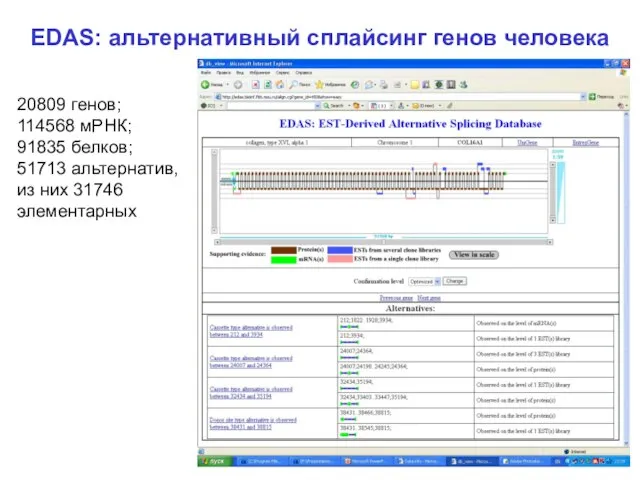
Слайд 6EDAS: альтернативный сплайсинг генов человека
20809 генов;
114568 мРНК;
91835 белков;
51713 альтернатив,
из них 31746 элементарных
EDAS: альтернативный сплайсинг генов человека
20809 генов;
114568 мРНК;
91835 белков;
51713 альтернатив,
из них 31746 элементарных
Слайд 7Альтернативная экзон-интроная структура генов млекопитающих
Тройки ортологичных генов: человек-мышь-собака
Следим за судьбой (консервативностью) альтернатив
Альтернативная экзон-интроная структура генов млекопитающих
Тройки ортологичных генов: человек-мышь-собака
Следим за судьбой (консервативностью) альтернатив

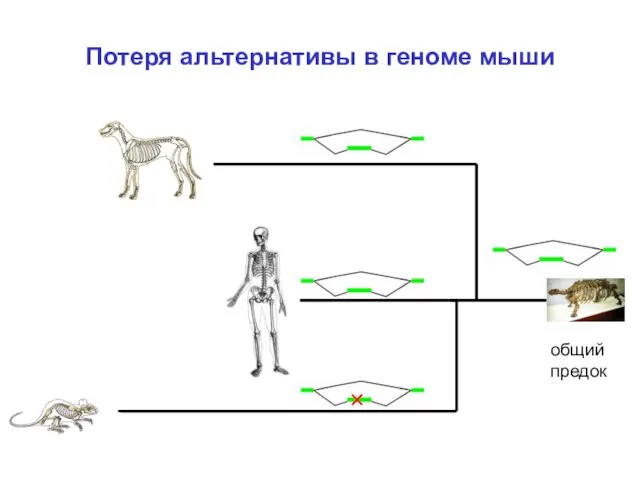
Слайд 8Потеря альтернативы в геноме мыши
общий
предок
Потеря альтернативы в геноме мыши
общий
предок
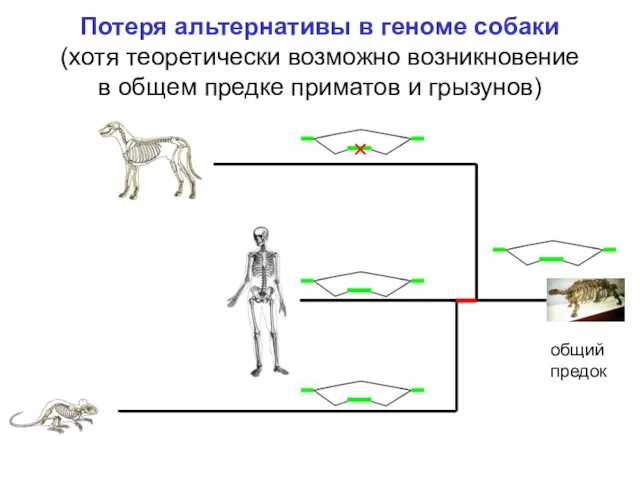
Слайд 9Потеря альтернативы в геноме собаки
(хотя теоретически возможно возникновение в общем предке приматов
Потеря альтернативы в геноме собаки (хотя теоретически возможно возникновение в общем предке приматов
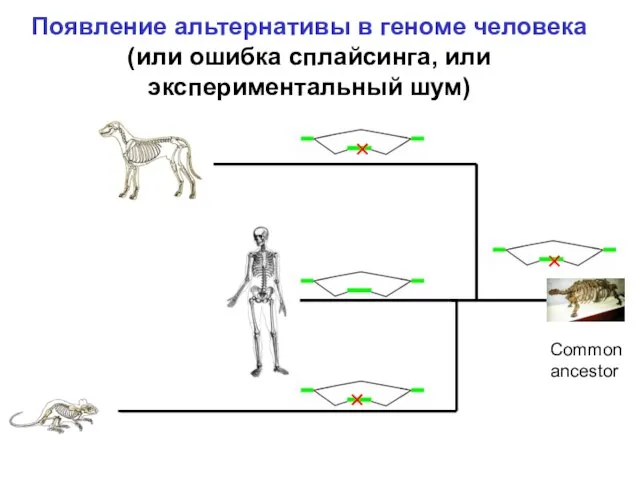
Слайд 10Появление альтернативы в геноме человека (или ошибка сплайсинга, или экспериментальный шум)
Common
ancestor
Появление альтернативы в геноме человека (или ошибка сплайсинга, или экспериментальный шум)
Common
ancestor
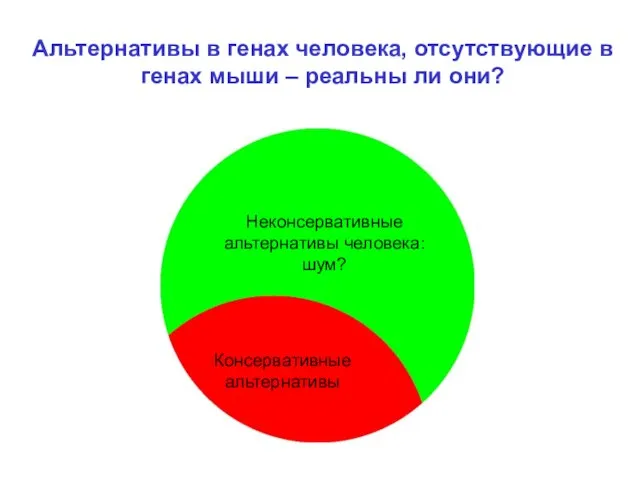
Слайд 11Неконсервативные
альтернативы человека:
шум?
Консервативные
альтернативы
Альтернативы в генах человека, отсутствующие в генах мыши
Неконсервативные
альтернативы человека:
шум?
Консервативные
альтернативы
Альтернативы в генах человека, отсутствующие в генах мыши
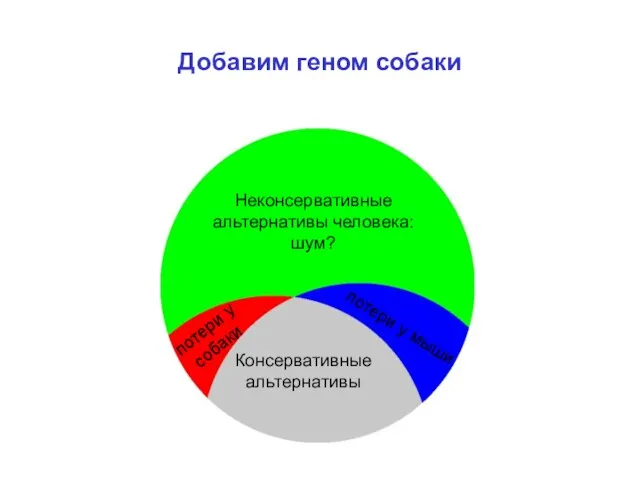
Слайд 12Human-specific alternatives:
noise?
Conserved
alternatives
Добавим геном собаки
Консервативные
альтернативы
потери у собаки
потери у мыши
Неконсервативные
альтернативы
Human-specific alternatives:
noise?
Conserved
alternatives
Добавим геном собаки
Консервативные
альтернативы
потери у собаки
потери у мыши
Неконсервативные альтернативы
Слайд 13Наблюдения
Часто вставляемые экзоны консервативны независимо от того, сбивают ли они рамку
Редко вставляемые
Наблюдения
Часто вставляемые экзоны консервативны независимо от того, сбивают ли они рамку
Редко вставляемые

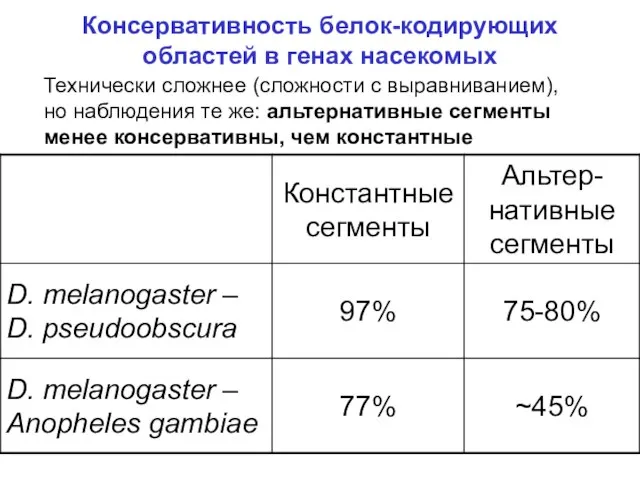
Слайд 14Консервативность белок-кодирующих областей в генах насекомых
Технически сложнее (сложности с выравниванием), но наблюдения
Консервативность белок-кодирующих областей в генах насекомых
Технически сложнее (сложности с выравниванием), но наблюдения
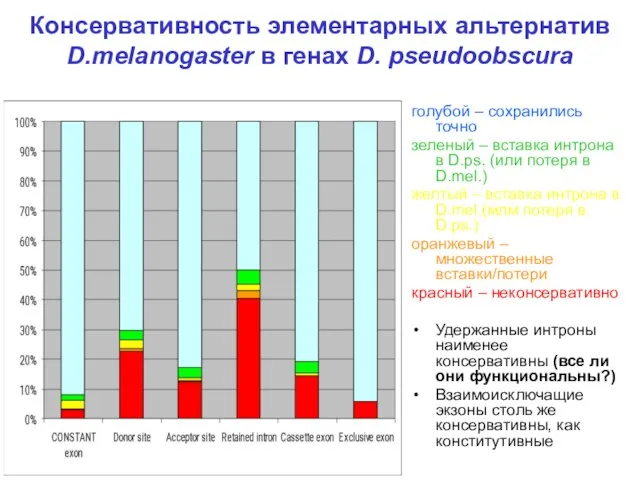
Слайд 15Консервативность элементарных альтернатив D.melanogaster в генах D. pseudoobscura
голубой – сохранились точно
зеленый –
Консервативность элементарных альтернатив D.melanogaster в генах D. pseudoobscura
голубой – сохранились точно
зеленый –
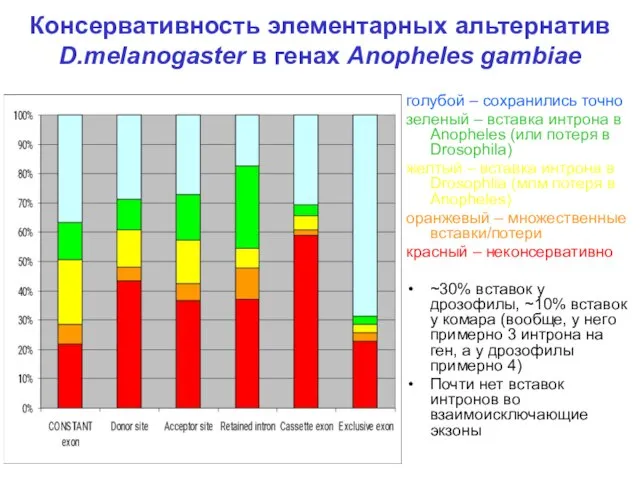
Слайд 16Консервативность элементарных альтернатив D.melanogaster в генах Anopheles gambiae
голубой – сохранились точно
зеленый –
Консервативность элементарных альтернатив D.melanogaster в генах Anopheles gambiae
голубой – сохранились точно
зеленый –
Слайд 17Скорость эволюции и тип отбора в альтернативных и константных областях
Пары ортологичных генов
человек
Скорость эволюции и тип отбора в альтернативных и константных областях
Пары ортологичных генов
человек

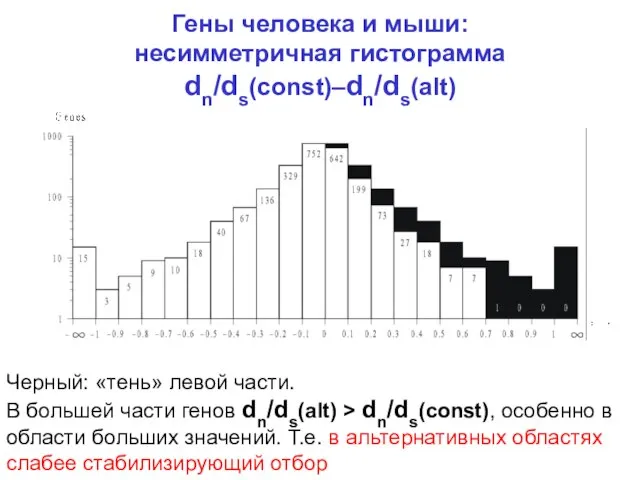
Слайд 18Гены человека и мыши:
несимметричная гистограмма
dn/ds(const)–dn/ds(alt)
Черный: «тень» левой части.
В большей части
Гены человека и мыши:
несимметричная гистограмма
dn/ds(const)–dn/ds(alt)
Черный: «тень» левой части. В большей части
Слайд 19Гены Drosophila:
слабее отбор в альтернативных областях?
Больше замен в альт. обл.
Равный уровень
Гены Drosophila:
слабее отбор в альтернативных областях?
Больше замен в альт. обл.
Равный уровень

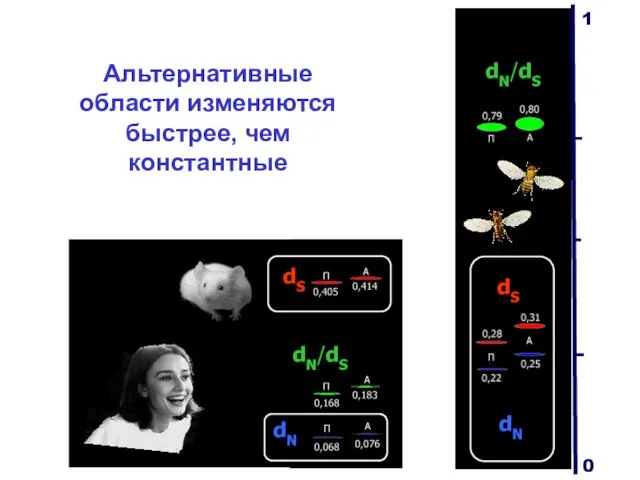
Слайд 20Альтернативные области изменяются быстрее, чем константные
dN
dN/dS
dS
dN/dS
dS
dN
1
0
Альтернативные области изменяются быстрее, чем константные
dN
dN/dS
dS
dN/dS
dS
dN
1
0
Слайд 21Ослабление стабилизирующего отбора в альтернативных областях
dN/dS
dN
dS
dN/dS
dS
dN
1
0
Ослабление стабилизирующего отбора в альтернативных областях
dN/dS
dN
dS
dN/dS
dS
dN
1
0

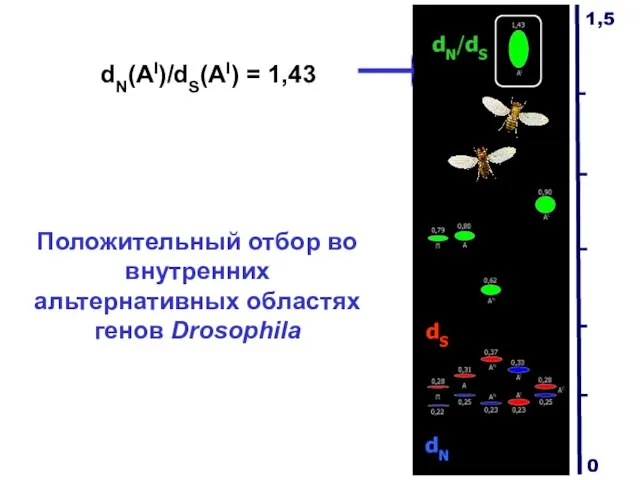
Слайд 22Положительный отбор во внутренних альтернативных областях генов Drosophila
dN/dS
dS
dN
dN(AI)/dS(AI) = 1,43
1,5
0
Положительный отбор во внутренних альтернативных областях генов Drosophila
dN/dS
dS
dN
dN(AI)/dS(AI) = 1,43
1,5
0
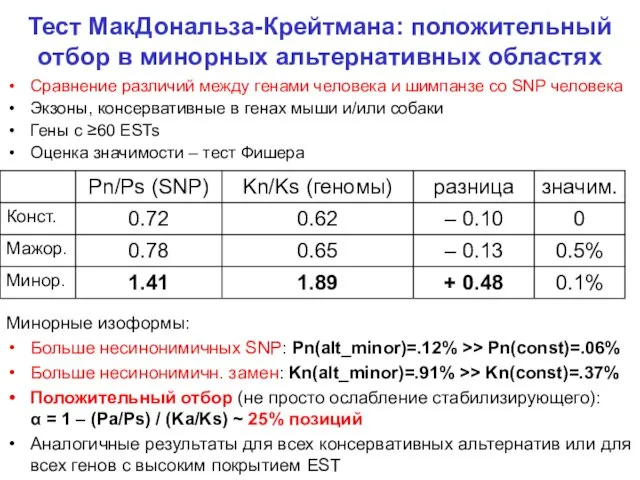
Слайд 23Тест МакДональза-Крейтмана: положительный отбор в минорных альтернативных областях
Сравнение различий между генами человека
Тест МакДональза-Крейтмана: положительный отбор в минорных альтернативных областях
Сравнение различий между генами человека
Слайд 24Попытка синтеза
Альтернативный сплайсинг часто видоспецифичен
«молодые» альтернативные изоформы часто тканеспецифичны
… но все же
Попытка синтеза
Альтернативный сплайсинг часто видоспецифичен
«молодые» альтернативные изоформы часто тканеспецифичны
… но все же

Слайд 25Планы: много дрозофил;
млекопитающие (ENCODE)
Планы: много дрозофил;
млекопитающие (ENCODE)
Слайд 26Что делать?
Оценить не только скорость потерь альтернатив, но и скорость приобретений (отличая
Что делать?
Оценить не только скорость потерь альтернатив, но и скорость приобретений (отличая

Слайд 27Благодарности
Обсуждения
Евгений Кунин (NCBI)
Игорь Рогозин (NCBI)
Всеволод Макеев (ГосНИИГенетика)
Дмитрий Петров (Stanford)
Дмитрий Фришман (GSF,
Благодарности
Обсуждения
Евгений Кунин (NCBI)
Игорь Рогозин (NCBI)
Всеволод Макеев (ГосНИИГенетика)
Дмитрий Петров (Stanford)
Дмитрий Фришман (GSF,

Слайд 28Авторы
Андрей Миронов (МГУ, ИППИ)
Рамиль Нуртдинов (МГУ)
– человек/мышь/собака
Дмитрий Малько (ГосНИИГенетика)
– дрозофилы/комар
Екатерина
Авторы
Андрей Миронов (МГУ, ИППИ)
Рамиль Нуртдинов (МГУ)
– человек/мышь/собака
Дмитрий Малько (ГосНИИГенетика)
– дрозофилы/комар
Екатерина

Слайд 29Основные публикации
10,1 D.B.Malko, V.J.Makeev, A.A.Mironov, M.S.Gelfand (2006) Evolution of the exon-intron structure and
Основные публикации
10,1 D.B.Malko, V.J.Makeev, A.A.Mironov, M.S.Gelfand (2006) Evolution of the exon-intron structure and

 Прыжок в высоту способом «перешагивание»
Прыжок в высоту способом «перешагивание» Избирательная система и система подсчета результатов голосования
Избирательная система и система подсчета результатов голосования Акробатические элементы. Упражнения на гибкость, растяжка, координация
Акробатические элементы. Упражнения на гибкость, растяжка, координация Английская революция
Английская революция Барлық жеңіс өзіңді-өзің жеңуден басталмақ
Барлық жеңіс өзіңді-өзің жеңуден басталмақ підготовка до ділових переговорів
підготовка до ділових переговорів Приклад заповнення ПКО
Приклад заповнення ПКО Презентация
Презентация Подготовила учитель истории Подготовила учитель истории МОУ Талаканской СОШ №6 Черткова Ольга Александровна
Подготовила учитель истории Подготовила учитель истории МОУ Талаканской СОШ №6 Черткова Ольга Александровна Основное энергетическое уравнение турбины. Кавитация в гидромашинах. Режимы работы гидромашин и их регулирование
Основное энергетическое уравнение турбины. Кавитация в гидромашинах. Режимы работы гидромашин и их регулирование Птица года-варакушка
Птица года-варакушка Северо-западный экономический район (9 класс)
Северо-западный экономический район (9 класс) Презентация на тему Основные закономерности возникновения государства
Презентация на тему Основные закономерности возникновения государства  Дробные числительные. 6 класс
Дробные числительные. 6 класс ПРОЕКТ «Волшебный мир семейного музея» Толубаева Е.Б., методист МОУ ДПО «Ме
ПРОЕКТ «Волшебный мир семейного музея» Толубаева Е.Б., методист МОУ ДПО «Ме Носов "Затейники" 2 класс
Носов "Затейники" 2 класс Kahrs Life. Новый тип деревянных полов
Kahrs Life. Новый тип деревянных полов Коламбия пикчерз не представляет…
Коламбия пикчерз не представляет… Трудные случаи пунктуации Запятая перед союзом И (Подготовка к ЕГЭ)
Трудные случаи пунктуации Запятая перед союзом И (Подготовка к ЕГЭ) Гимнастика в 5-11 классах
Гимнастика в 5-11 классах Мультик с шаром
Мультик с шаром Итоги государственной (итоговой) аттестации -2012
Итоги государственной (итоговой) аттестации -2012 Презентация на тему Решение задач 1 класс
Презентация на тему Решение задач 1 класс  МЛУ-ТБ:Приверженность лечению
МЛУ-ТБ:Приверженность лечению L_r_1_Kriminalisticheskaya_fotografia
L_r_1_Kriminalisticheskaya_fotografia Ослепительная улыбка на всю жизнь
Ослепительная улыбка на всю жизнь ПОРТФОЛИО
ПОРТФОЛИО Российский союз выставок и ярмарок как эффективная площадка коммуникаций Выездное заседание ЭКСПОКЛУБА «Актуальные вопросы выст
Российский союз выставок и ярмарок как эффективная площадка коммуникаций Выездное заседание ЭКСПОКЛУБА «Актуальные вопросы выст