- Гормоны поджелудочной железы

Содержание
- 2. Инсулин Инсули́н (от лат. insula — остров) — гормон пептидной природы. Молекула инсулина образована двумя полипептидными
- 3. Синтез инсулина в клетке Синтез и выделение инсулина представляют собой сложный процесс, включающий несколько этапов. Первоначально
- 4. Структурная формула На схеме показана последовательность аминокислот в молекуле инсулина: А-цепь содержит 21 аминокислотный остаток, Б-цепь
- 5. Секреция инсулина Бета-клетки островков Лангерганса поджелудочной железы чувствительны к изменению уровня глюкозы в крови; выделение ими
- 6. Регуляция образования и секреции инсулина Главным стимулятором освобождения инсулина является повышение уровня глюкозы в крови. Дополнительно
- 7. Механизм действия инсулина Так или иначе, инсулин затрагивает все виды обмена веществ во всём организме. Однако
- 8. Эффекты вызываемые инсулином Физиологические эффекты инсулина Инсулин оказывает на обмен веществ и энергии сложное и многогранное
- 9. Регуляция уровня глюкозы в крови Поддержание оптимальной концентрации глюкозы в крови - результат действия множества факторов,
- 10. Заболевания, связанные с действием инсулина Гипергликемия — увеличение уровня сахара в крови. В состоянии гипергликемии увеличивается
- 11. Глюкагон Глюкагон (син. гипергликемическо-гликогенолитический фактор) - белково-пептидный гормон поджелудочной железы, участвующий в регуляции углеводного обмена. Он
- 12. Секреция и синтез глюкагона Гормон синтезируется в альфа-клетках островков Лангерганса. В организме продуцируется также кишечный глюкагон
- 13. Механизм действия и эффекты глюкагона Глюкагон воздействует главным образом на печень, где немедленно стимулирует гликогенолиз, а
- 14. Действие глюкагона Глюкагон оказывает сильное инотропное и хронотропное действие на миокард вследствие увеличения образования цАМФ (то
- 16. Скачать презентацию

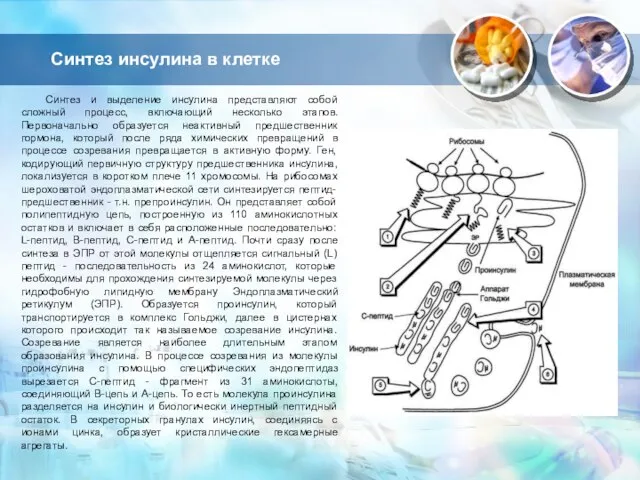
Слайд 3Синтез инсулина в клетке
Синтез и выделение инсулина представляют собой сложный процесс, включающий
Синтез инсулина в клетке
Синтез и выделение инсулина представляют собой сложный процесс, включающий
Слайд 4Структурная формула
На схеме показана последовательность аминокислот в молекуле инсулина: А-цепь содержит 21
Структурная формула
На схеме показана последовательность аминокислот в молекуле инсулина: А-цепь содержит 21

Слайд 5Секреция инсулина
Бета-клетки островков Лангерганса поджелудочной железы чувствительны к изменению уровня глюкозы в
Секреция инсулина
Бета-клетки островков Лангерганса поджелудочной железы чувствительны к изменению уровня глюкозы в

Слайд 6Регуляция образования и секреции инсулина
Главным стимулятором освобождения инсулина является повышение уровня глюкозы
Регуляция образования и секреции инсулина
Главным стимулятором освобождения инсулина является повышение уровня глюкозы

Слайд 7Механизм действия инсулина
Так или иначе, инсулин затрагивает все виды обмена веществ во
Механизм действия инсулина
Так или иначе, инсулин затрагивает все виды обмена веществ во

Слайд 8Эффекты вызываемые инсулином
Физиологические эффекты инсулина Инсулин оказывает на обмен веществ и энергии
Эффекты вызываемые инсулином
Физиологические эффекты инсулина Инсулин оказывает на обмен веществ и энергии

Слайд 9Регуляция уровня глюкозы в крови
Поддержание оптимальной концентрации глюкозы в крови - результат
Регуляция уровня глюкозы в крови
Поддержание оптимальной концентрации глюкозы в крови - результат

Слайд 10Заболевания, связанные с действием инсулина
Гипергликемия — увеличение уровня сахара в крови. В
Заболевания, связанные с действием инсулина
Гипергликемия — увеличение уровня сахара в крови. В

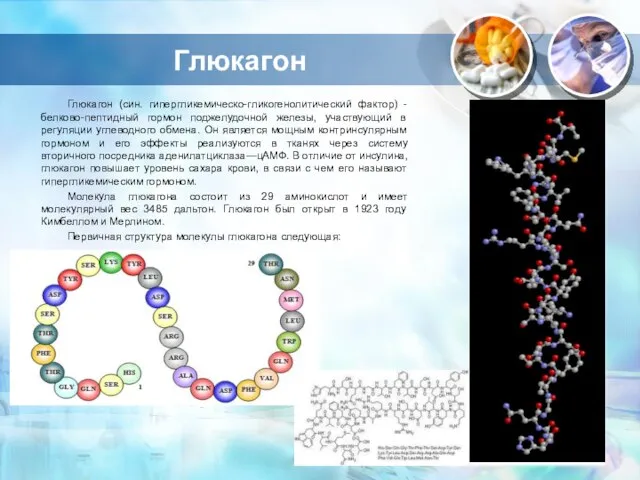
Слайд 11Глюкагон
Глюкагон (син. гипергликемическо-гликогенолитический фактор) - белково-пептидный гормон поджелудочной железы, участвующий в регуляции
Глюкагон
Глюкагон (син. гипергликемическо-гликогенолитический фактор) - белково-пептидный гормон поджелудочной железы, участвующий в регуляции
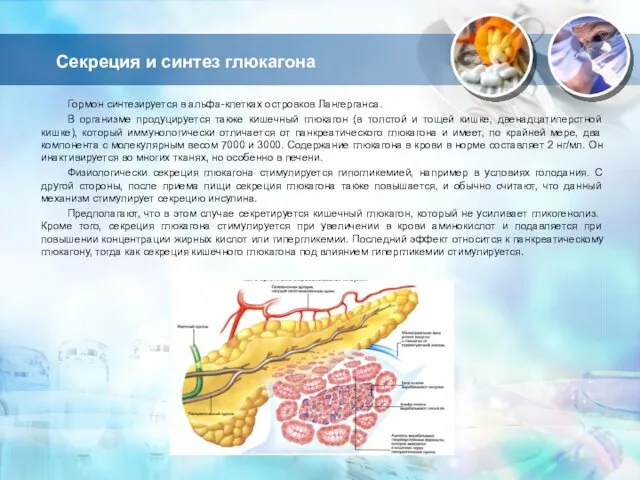
Слайд 12Секреция и синтез глюкагона
Гормон синтезируется в альфа-клетках островков Лангерганса.
В организме продуцируется
Секреция и синтез глюкагона
Гормон синтезируется в альфа-клетках островков Лангерганса.
В организме продуцируется
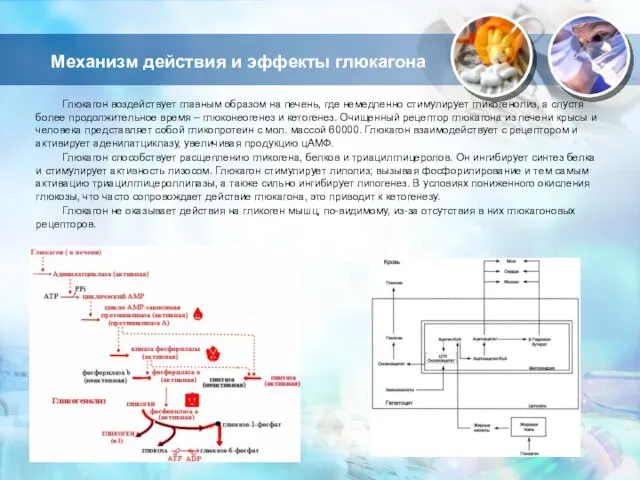
Слайд 13Механизм действия и эффекты глюкагона
Глюкагон воздействует главным образом на печень, где немедленно
Механизм действия и эффекты глюкагона
Глюкагон воздействует главным образом на печень, где немедленно
Слайд 14Действие глюкагона
Глюкагон оказывает сильное инотропное и хронотропное действие на миокард вследствие увеличения
Действие глюкагона
Глюкагон оказывает сильное инотропное и хронотропное действие на миокард вследствие увеличения

 Презентация на тему: Овощи и фрукты – самые витаминные продукты!
Презентация на тему: Овощи и фрукты – самые витаминные продукты! Авторское право
Авторское право Лекция 7. Духовно-нравственное воспитание и общие основы православной педагогики
Лекция 7. Духовно-нравственное воспитание и общие основы православной педагогики Информационное обеспечение контрактной системы в сфере закупок
Информационное обеспечение контрактной системы в сфере закупок Деньги
Деньги Система, многообразие и эволюция живой природы
Система, многообразие и эволюция живой природы АППЛИКАЦИЯ ИЗ НИТОК
АППЛИКАЦИЯ ИЗ НИТОК Пожилые
Пожилые Технология обработки пищевых продуктов
Технология обработки пищевых продуктов Рязань
Рязань Fleurs de France
Fleurs de France Факультет з підготовки іноземних громадян
Факультет з підготовки іноземних громадян «Автономные «Зеленые» Поселения» Проект Международной Программы «СОЛНЕЧНЫЙ ПОТОК»
«Автономные «Зеленые» Поселения» Проект Международной Программы «СОЛНЕЧНЫЙ ПОТОК» Naelsya_i_spit (1)
Naelsya_i_spit (1) Прийняття мисцевого бюджету
Прийняття мисцевого бюджету Реклама как один из способов воздействия на сознание подростков, совершающих правонарушения
Реклама как один из способов воздействия на сознание подростков, совершающих правонарушения Фурнитура
Фурнитура Проектирование сухого отсека ракеты-носителя из композиционных материалов с применением численной оптимизации
Проектирование сухого отсека ракеты-носителя из композиционных материалов с применением численной оптимизации Источники трудового права
Источники трудового права The house of my dream izmen
The house of my dream izmen Презентация "Василий Григорьевич Перов" - скачать презентации по МХК
Презентация "Василий Григорьевич Перов" - скачать презентации по МХК Управление себестоимостью и прибылью
Управление себестоимостью и прибылью Игра как эффективное средство обучения английскому
Игра как эффективное средство обучения английскому Плетеные картины
Плетеные картины Каталог факультета психологии
Каталог факультета психологии Заключение договоров в МИЭС
Заключение договоров в МИЭС Презентация на тему НЕРАВЕНСТВО ДОХОДОВ В ОБЩЕСТВЕ
Презентация на тему НЕРАВЕНСТВО ДОХОДОВ В ОБЩЕСТВЕ  Элективный курс«Химик – исследователь»
Элективный курс«Химик – исследователь»