Индивидуальная изменчивость видов рода Calligonum L. и её свойства (на основе монографического изучения рода Жузгун – Calligonum L.)
- Индивидуальная изменчивость видов рода Calligonum L. и её свойства (на основе монографического изучения рода Жузгун – Calligonum L.)

Содержание
- 2. Род Жузгун – Calligonum L. из семейства Polygonaceae, голарктический, афро-азиатский, крайне полиморфный, представлен 4 секциями и
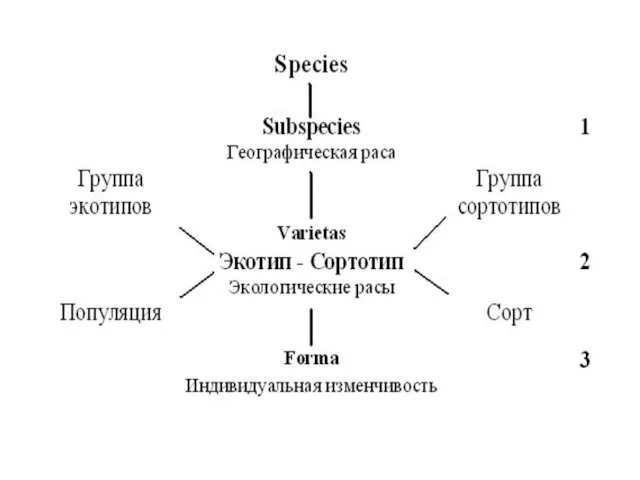
- 3. Тема нашего сообщения касается только третьей подсистемы вида, с которой начинается изучение любого вида, а именно
- 5. Жузгун бакинский – Calligonum bakuense Litv. На берегу Каспийского моря у пос. Дуванный на п-ве Апшерон
- 6. Calligonum bakuense Litv. Эндем Апшеронского полуострова на Кавказе
- 7. Индивидуальная изменчивость плодов дикорастущей популяции Calligonum bakuense Litv. Азербайджан, 59 км юго-западнее г. Баку, по берегу
- 8. Частота встречаемости разностей признаков и их комбинаций (фенов, морфодемов) в популяции Calligonum bakuense Litv. на Каспийском
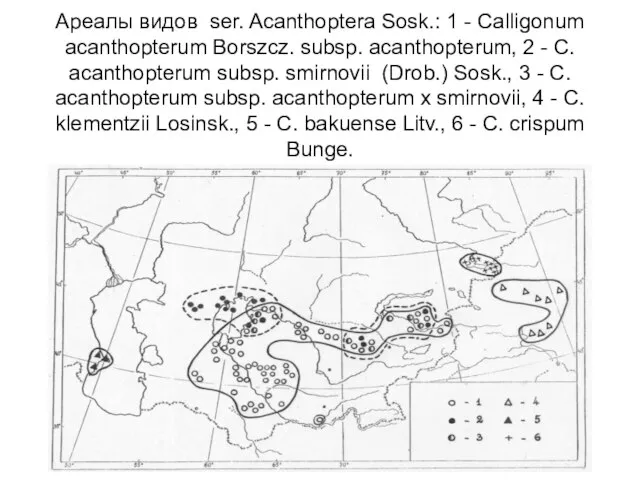
- 9. Ареалы видов ser. Acanthoptera Sosk.: 1 - Calligonum acanthopterum Borszcz. subsp. acanthopterum, 2 - C. acanthopterum
- 10. Зайсанский C. rubicundum Bunge В ареале этого вида, в Зайсанской котловине, где произрастают только 2 вида
- 11. Туранский Calligonum acanthopterum Borszcz. Ярко выраженный фен (морфодем)
- 12. Южнотуранский C. setosum (Litv.) Litv. Два куста с различной окраской и величиной плода
- 13. Кодовая схема изменчивости видов рода жузгун - Calligonum L. по индивидуальным морфологи- ческим признакам репродуктивной сферы
- 14. Кодовая схема … Продолжение 6. Ширина гребня крыла, мм: 6.1 - узкий (1-3 мм); 6.2 -
- 15. Кодовая схема…Продолжение 12. Плотность стояния щетинок: 12.1 - редкое; 12.2 - среднее; 12.3 - густое. 13.
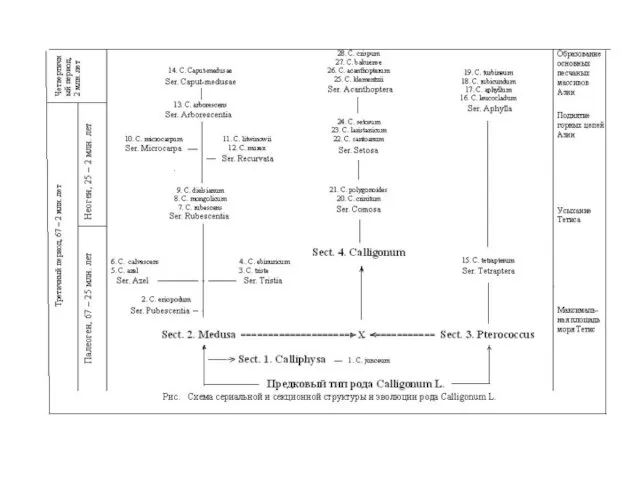
- 16. Имеют изменчивость по 1-2 признакам виды неогенового возраста: Calligonum junceum, C. eriopodum, C. triste, C. ebinuricum,
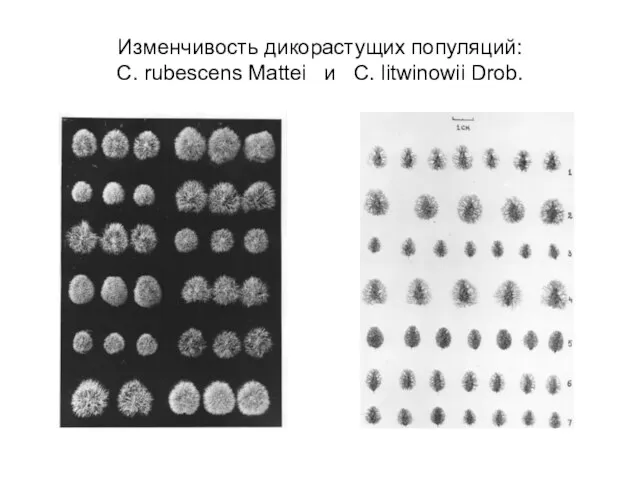
- 18. Изменчивость дикорастущих популяций: C. rubescens Mattei и C. litwinowii Drob.
- 19. Ареалы видов ser. Rubescentia Sosk.: 1 - Calligonum rubescens Mattei, 2 - C. mongolicum Turcz., 3
- 20. Ареалы видов ser. Recurvata Sosk.: 1 - Calligonum litwinowii, 2 - C. murex Bunge.
- 21. Изменчивость дикорастущих популяций C. leucocladum (Schrenk.) Bunge и C. aphyllum (Pall.) Guerke
- 22. Сверотуранский Calligonum aphyllum (Pall.) Guerke
- 23. Изменчивость дикорастущих популяций C. polygonoides L. и C. acanthopterum Borszcz.
- 24. Ареалы видов ser. Comosa Sosk.: 1 - Calligonum crinitum Boiss. subsp. crinitum, 2 - C. crinitum
- 25. Без выяснения ареалов у видов в роде Calligonum L. было напрасно описано только по наиболее часто
- 26. В то же время, отдельные индивидуальные признаки, перечисленные в кодовой таблице могут менять свой ранг и
- 27. Коэффициент вариации по индивидуальным морфологическим (качественным) признакам у наиболее полиморфных видов рода Calligonum находится в пределах
- 29. Согласно закона Гарди-Вайнберга (Лобашев, 1967, с. 613-616; Ли, 1978, с.11; Яблоков, 1980, с. 86 и др.)
- 30. Изменчивость в потомстве одного растения зайсанского C. rubicundum (слева) и северотуранского C. aphyllum c песков Кумторкале
- 31. Коэффициент вариации в условиях естественного произрастания у крайне полиморфных видов C. acanthop- terum, C. aphyllum, C.
- 32. . Для видов с высоким уровнем индивидуальной изменчивости характерен и высокий уровень гомологической изменчивости, которая проявляется
- 33. Гомологическая изменчивость Данная таблица, согласно закона гомологических рядов Н.И. Вавилова, отражает параллельную изменчивость по 8 признакам
- 34. На основании литературных данных можно сделать заключение, что в области индивидуальной изменчивости действуют также три закона
- 35. Северотуранский Calligonum aphyllum
- 36. Северотуранский Calligonum aphyllum
- 37. C. aphyllum (цв.) и C. rubescens (пл.)
- 38. C. turbineum Pavl. - муюнкумский гибридогенный высокорослый вид, в налегающей части ареалов исходных видов – C.
- 39. C. x dubianskii nm. androssowii (C. acanthopterum x C. leucocladum)
- 40. C. x spinulosum (C. acanthopterum x C. aphyllum)
- 41. Южнотуранский C. leucocladum (Schrenk) Bunge, экотип низкорослый извилистый, симпатрический по отношению к другому экотипу – высокорослому
- 42. Южнотуранский C. eriopodum Bunge – многочисленные деревца на заднем плане
- 43. Североприаральский C. murex Bunge – большой песчаный бугор в центре с многочисленными парциальными кустами. Малые Барсуки.
- 44. Североприаральский C. junceum subsp. junceum – два экземпляра стелющегося кустарничка в бедлендовой пустыне Северного Приаралья, чинк
- 45. Коллекция жузгуна на Приаральской опытной станции ВИР, ст. Челкар, 1972 г.
- 47. Скачать презентацию
Слайд 2
Род Жузгун – Calligonum L. из семейства Polygonaceae, голарктический, афро-азиатский, крайне полиморфный,
Род Жузгун – Calligonum L. из семейства Polygonaceae, голарктический, афро-азиатский, крайне полиморфный,

Слайд 3 Тема нашего сообщения касается только третьей подсистемы вида, с которой начинается
Тема нашего сообщения касается только третьей подсистемы вида, с которой начинается

Слайд 5Жузгун бакинский – Calligonum bakuense Litv. На берегу Каспийского моря у пос.
Жузгун бакинский – Calligonum bakuense Litv. На берегу Каспийского моря у пос.

Слайд 6Calligonum bakuense Litv. Эндем Апшеронского полуострова на Кавказе
Calligonum bakuense Litv. Эндем Апшеронского полуострова на Кавказе

Слайд 7Индивидуальная изменчивость плодов дикорастущей популяции Calligonum bakuense Litv. Азербайджан, 59 км юго-западнее
Индивидуальная изменчивость плодов дикорастущей популяции Calligonum bakuense Litv. Азербайджан, 59 км юго-западнее

Слайд 8
Частота встречаемости разностей признаков и их комбинаций (фенов, морфодемов) в популяции Calligonum
Частота встречаемости разностей признаков и их комбинаций (фенов, морфодемов) в популяции Calligonum

Слайд 9
Ареалы видов ser. Acanthoptera Sosk.: 1 - Calligonum acanthopterum Borszcz. subsp. acanthopterum,
Ареалы видов ser. Acanthoptera Sosk.: 1 - Calligonum acanthopterum Borszcz. subsp. acanthopterum,
Слайд 10Зайсанский C. rubicundum Bunge
В ареале этого вида, в Зайсанской котловине, где произрастают
Зайсанский C. rubicundum Bunge В ареале этого вида, в Зайсанской котловине, где произрастают

Слайд 11Туранский Calligonum acanthopterum Borszcz.
Ярко выраженный фен (морфодем)
Туранский Calligonum acanthopterum Borszcz.
Ярко выраженный фен (морфодем)

Слайд 12Южнотуранский C. setosum (Litv.) Litv.
Два куста с различной окраской и величиной плода
Южнотуранский C. setosum (Litv.) Litv.
Два куста с различной окраской и величиной плода

Слайд 13Кодовая схема изменчивости видов рода жузгун
- Calligonum L. по индивидуальным морфологи-
ческим
- Calligonum L. по индивидуальным морфологи-
ческим

Слайд 14
Кодовая схема … Продолжение
6. Ширина гребня крыла, мм: 6.1 - узкий (1-3
Кодовая схема … Продолжение
6. Ширина гребня крыла, мм: 6.1 - узкий (1-3

Слайд 15Кодовая схема…Продолжение
12. Плотность стояния щетинок: 12.1 - редкое; 12.2 - среднее; 12.3
12. Плотность стояния щетинок: 12.1 - редкое; 12.2 - среднее; 12.3

Слайд 16Имеют изменчивость по 1-2 признакам виды неогенового возраста:
Calligonum junceum, C. eriopodum,
Имеют изменчивость по 1-2 признакам виды неогенового возраста:
Calligonum junceum, C. eriopodum,

Слайд 18Изменчивость дикорастущих популяций:
C. rubescens Mattei и C. litwinowii Drob.
Изменчивость дикорастущих популяций:
C. rubescens Mattei и C. litwinowii Drob.
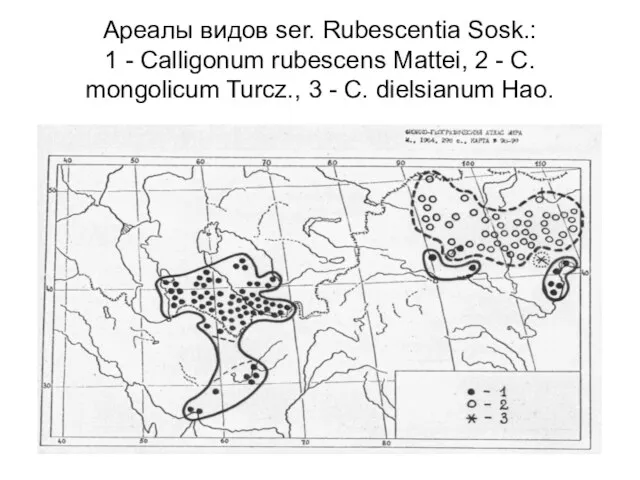
Слайд 19Ареалы видов ser. Rubescentia Sosk.:
1 - Calligonum rubescens Mattei, 2 -
Ареалы видов ser. Rubescentia Sosk.: 1 - Calligonum rubescens Mattei, 2 -
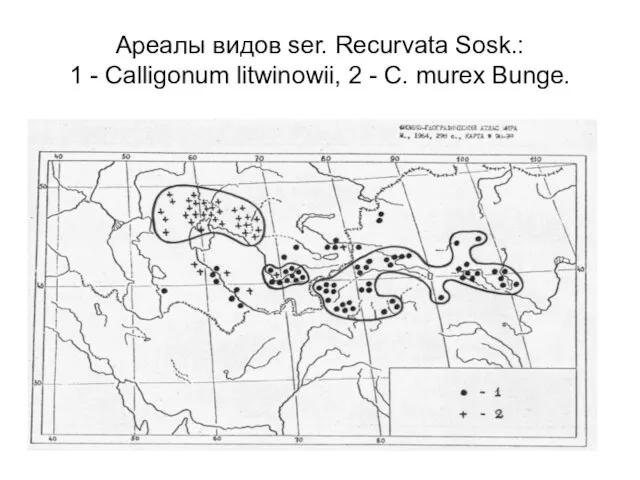
Слайд 20Ареалы видов ser. Recurvata Sosk.:
1 - Calligonum litwinowii, 2 - C.
Ареалы видов ser. Recurvata Sosk.: 1 - Calligonum litwinowii, 2 - C.
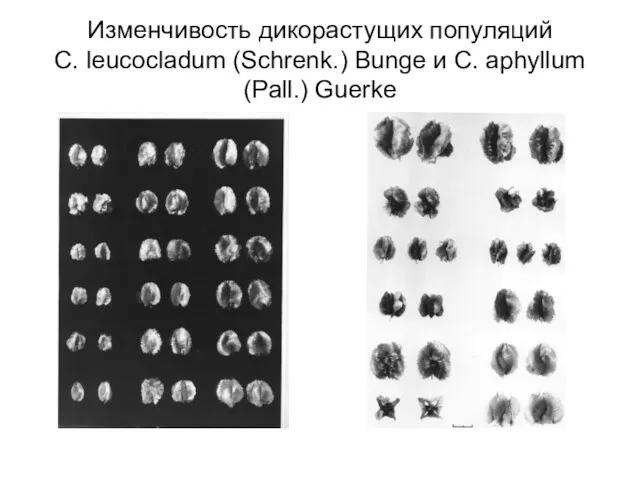
Слайд 21Изменчивость дикорастущих популяций
C. leucocladum (Schrenk.) Bunge и C. aphyllum (Pall.) Guerke
Изменчивость дикорастущих популяций
C. leucocladum (Schrenk.) Bunge и C. aphyllum (Pall.) Guerke
Слайд 22Сверотуранский Calligonum aphyllum (Pall.) Guerke
Сверотуранский Calligonum aphyllum (Pall.) Guerke

Слайд 23Изменчивость дикорастущих популяций
C. polygonoides L. и C. acanthopterum Borszcz.
Изменчивость дикорастущих популяций
C. polygonoides L. и C. acanthopterum Borszcz.

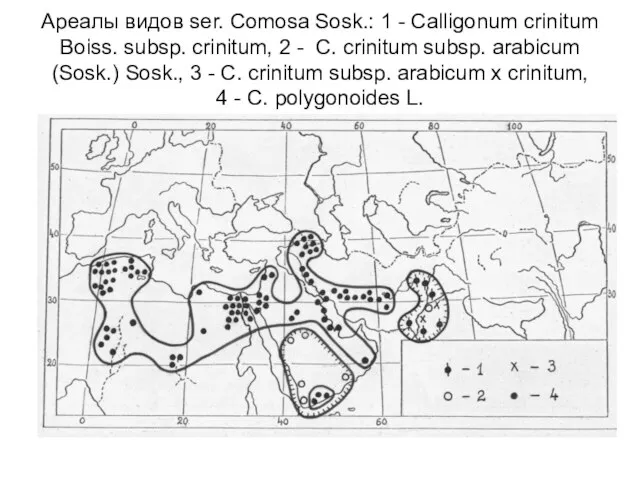
Слайд 24Ареалы видов ser. Comosa Sosk.: 1 - Calligonum crinitum Boiss. subsp. crinitum,
Ареалы видов ser. Comosa Sosk.: 1 - Calligonum crinitum Boiss. subsp. crinitum,
Слайд 25 Без выяснения ареалов у видов в роде Calligonum L. было напрасно
Без выяснения ареалов у видов в роде Calligonum L. было напрасно

Слайд 26В то же время, отдельные индивидуальные признаки, перечисленные в кодовой таблице могут
В то же время, отдельные индивидуальные признаки, перечисленные в кодовой таблице могут

Слайд 27 Коэффициент вариации по индивидуальным морфологическим
(качественным) признакам у наиболее полиморфных видов
Коэффициент вариации по индивидуальным морфологическим
(качественным) признакам у наиболее полиморфных видов

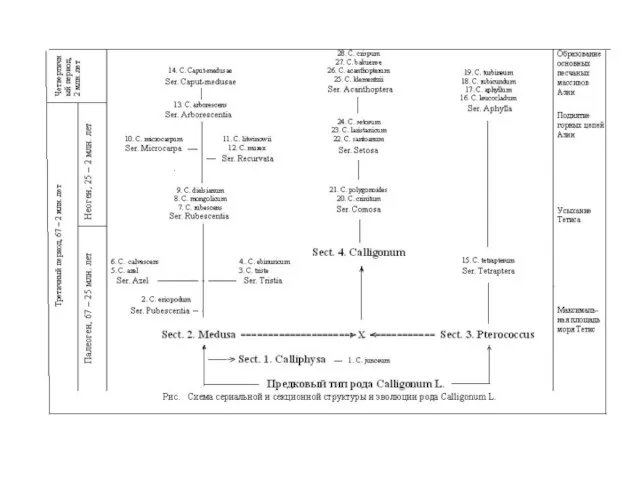
Слайд 29 Согласно закона Гарди-Вайнберга (Лобашев, 1967, с. 613-616;
Ли, 1978, с.11; Яблоков,
Согласно закона Гарди-Вайнберга (Лобашев, 1967, с. 613-616;
Ли, 1978, с.11; Яблоков,

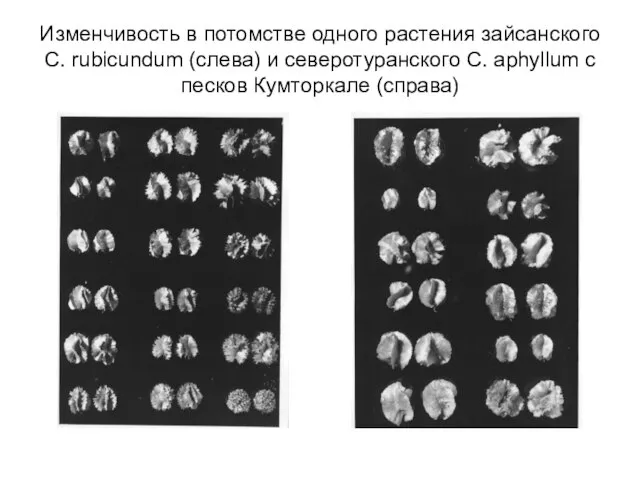
Слайд 30Изменчивость в потомстве одного растения зайсанского C. rubicundum (слева) и северотуранского C.
Изменчивость в потомстве одного растения зайсанского C. rubicundum (слева) и северотуранского C.
Слайд 31 Коэффициент вариации в условиях естественного
произрастания у крайне полиморфных видов C.
Коэффициент вариации в условиях естественного
произрастания у крайне полиморфных видов C.

Слайд 32. Для видов с высоким уровнем индивидуальной изменчивости
характерен и высокий уровень
. Для видов с высоким уровнем индивидуальной изменчивости
характерен и высокий уровень

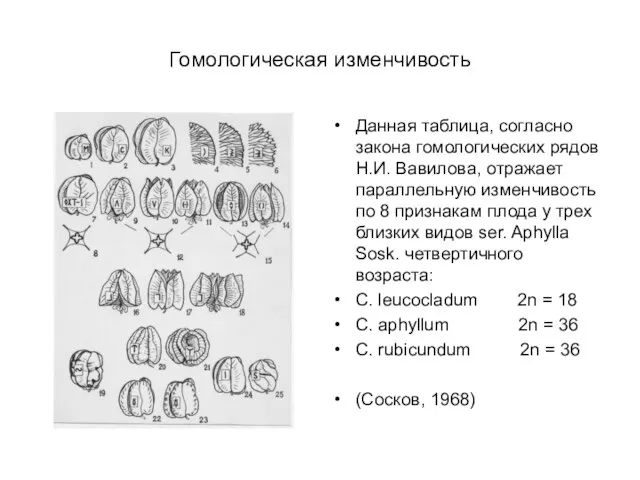
Слайд 33Гомологическая изменчивость
Данная таблица, согласно закона гомологических рядов Н.И. Вавилова, отражает параллельную изменчивость
Гомологическая изменчивость
Данная таблица, согласно закона гомологических рядов Н.И. Вавилова, отражает параллельную изменчивость
Слайд 34 На основании литературных данных можно сделать заключение, что в области индивидуальной
На основании литературных данных можно сделать заключение, что в области индивидуальной

Слайд 35Северотуранский Calligonum aphyllum
Северотуранский Calligonum aphyllum

Слайд 36Северотуранский Calligonum aphyllum
Северотуранский Calligonum aphyllum

Слайд 37C. aphyllum (цв.) и C. rubescens (пл.)
C. aphyllum (цв.) и C. rubescens (пл.)

Слайд 38C. turbineum Pavl. - муюнкумский гибридогенный высокорослый вид, в налегающей части ареалов
C. turbineum Pavl. - муюнкумский гибридогенный высокорослый вид, в налегающей части ареалов

Слайд 39C. x dubianskii nm. androssowii
(C. acanthopterum x C. leucocladum)
C. x dubianskii nm. androssowii
(C. acanthopterum x C. leucocladum)

Слайд 40C. x spinulosum (C. acanthopterum x C. aphyllum)
C. x spinulosum (C. acanthopterum x C. aphyllum)

Слайд 41Южнотуранский C. leucocladum (Schrenk) Bunge,
экотип низкорослый извилистый, симпатрический по отношению к другому
Южнотуранский C. leucocladum (Schrenk) Bunge, экотип низкорослый извилистый, симпатрический по отношению к другому

Слайд 42Южнотуранский C. eriopodum Bunge –
многочисленные деревца
на заднем плане
Южнотуранский C. eriopodum Bunge –
многочисленные деревца
на заднем плане

Слайд 43Североприаральский C. murex Bunge – большой песчаный бугор в центре с многочисленными
Североприаральский C. murex Bunge – большой песчаный бугор в центре с многочисленными

Слайд 44Североприаральский C. junceum subsp. junceum – два экземпляра стелющегося кустарничка в бедлендовой
Североприаральский C. junceum subsp. junceum – два экземпляра стелющегося кустарничка в бедлендовой

Слайд 45Коллекция жузгуна на Приаральской опытной станции ВИР, ст. Челкар, 1972 г.
Коллекция жузгуна на Приаральской опытной станции ВИР, ст. Челкар, 1972 г.

 Украина. Донецкая область. Коммунальное предприятие Коммунальник. Функции юридического отдела
Украина. Донецкая область. Коммунальное предприятие Коммунальник. Функции юридического отдела Презентация на тему ПОЛЕВЫЕ ЦВЕТЫ
Презентация на тему ПОЛЕВЫЕ ЦВЕТЫ  여행을 대신해 드립니다_제작지원 제안서_220922_카자흐스탄
여행을 대신해 드립니다_제작지원 제안서_220922_카자흐스탄 Britain
Britain Как превратить клиентов в фанатов
Как превратить клиентов в фанатов Возмещение расходов на осуществление капитального или текущего ремонта жилых помещений
Возмещение расходов на осуществление капитального или текущего ремонта жилых помещений Презентация на тему Вещественные доказательства. Понятие вещественных доказательств. Значение вещественных доказательств
Презентация на тему Вещественные доказательства. Понятие вещественных доказательств. Значение вещественных доказательств  Несмертельна зброя
Несмертельна зброя Трансформация личности
Трансформация личности В. кзал х. зяин м. к. роны п. льто уч. ник ч. ловек о о аа а е е.
В. кзал х. зяин м. к. роны п. льто уч. ник ч. ловек о о аа а е е. Признаки текста. Анализ текста
Признаки текста. Анализ текста Калуга
Калуга Rubbish polution
Rubbish polution  Гидросфера
Гидросфера Первая медицинская помощь при ранениях
Первая медицинская помощь при ранениях Квалификационная работа: Оптимизация производства буровзрывных работ
Квалификационная работа: Оптимизация производства буровзрывных работ Мониторинг учебного процесса в начальной школе.
Мониторинг учебного процесса в начальной школе. Методические рекомендации по планированию работы библиотек МБУК ПР МЦБ на 2022год
Методические рекомендации по планированию работы библиотек МБУК ПР МЦБ на 2022год Сравнительная характеристика критериев NPV и IRR
Сравнительная характеристика критериев NPV и IRR Презентация на тему «Понятие и сущность государства» по предмету ТГП. Подготовила Прыгунова Диана Сергеевна.
Презентация на тему «Понятие и сущность государства» по предмету ТГП. Подготовила Прыгунова Диана Сергеевна.  Дизайн-проект
Дизайн-проект Система работы по профилактике правонарушений и безнадзорности
Система работы по профилактике правонарушений и безнадзорности Технологические предложения по производству и переработке полбы в крупу
Технологические предложения по производству и переработке полбы в крупу Особенности развития хозяйства России
Особенности развития хозяйства России Самоопределение личности школьника
Самоопределение личности школьника Умный Дом 2010 07
Умный Дом 2010 07 Нестандартный урок. Каков он?
Нестандартный урок. Каков он? Транспортная логистика
Транспортная логистика