- Мейоз у грибов

Содержание
- 2. Мейоз - это деление диплоидного ядра, при котором число хромосом уменьшается от диплоидного до гаплоидного. Мейоз
- 3. Наиболее изученные грибы относительно мейоза Neurospora crassa, Sordaria fimicola, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Aspergillus nidulans, Ascobolus
- 4. Преимущества мицелиальных грибов для изучения мейоза Короткий жизненный цикл за время которого может быть проанализировано несколько
- 5. Этапы формирования аскоспор
- 7. Этапы формирования базидий
- 8. Особенности профазы I мейоза Профаза I мейоза – уникальная, протяженная стадия (3-10 часов) с активной транскрипцией
- 10. Два кроссинговера Два кроссинговера
- 11. Число хромосом в ядрах разных видов грибов
- 12. Мейоз у Sordaria macrospora
- 13. Перед кариогамией происходит удвоение ДНК (S-фаза клеточного цикла грибов). При мейозе S-фаза длиннее, чем при митозе.
- 14. Премейотические механизмы “Checking and Cleaning” работают перед S-фазой Repeat-induced-point mutation (RIP) mechanism (механизм повторно индуцированных точковых
- 15. A “Checking” Mechanism that Operates After Karyogamy meiotic silencing by unpaired DNA (MSUD)
- 16. Стадии мейоза Первое деление мейоза профаза I лептотена зиготена пахитена диплотенa диакинез
- 17. Первое деление мейоза метафаза I анафаза I телофаза I
- 18. Второе деление мейоза профаза II метафаза II анафаза II телофаза II
- 19. Профаза I. Лептотена формирование осевых элементов, сближение гомологов
- 20. Профаза I. Зиготена начало сборки синаптонемного комплекса в сайтах инициации синапсиса (букет)
- 21. Профаза I. Пахитена Стадия полного синапсиса. Синаптонемные комплексы – трилинейные нуклеопротеидные структуры состоят из двух боковых
- 22. A–C Synaptonemal complex and recombination nodules of S. macrospora. A Pachytene synaptonemal complex. LE indicates maternal
- 23. Модель синаптонемного комплекса
- 25. Профаза I. Диплотена, диакинез Десинапсис гомологов, разборка синаптонемного комплекса. В конце концов гомологи остаются скрепленными лишь
- 26. Мейоз у Saccharomyces cerevisiae
- 27. Мейоз у Saccharomyces cerevisiae
- 30. МЕЙОЗ II Saccharomyces cerevisiae
- 34. Sherwood, Bennett, 2009 Sherwood, Bennett, 2009 Синхронность мейоза у дрожжей требует согласованной работы более 1000 генов\продуктов
- 35. Регуляция полового размножения у дрожжей Phylogenetic tree of the hemiascomycetes (S. cerevisiae and sequenced Candida species)
- 38. 1 стадия перед слиянием (24 часа) 2-3 стадия лептотены после слияния перед синапсисом 4 зиготена Мейоз
- 39. пахитена профазы 1 ранняя пахитена профазы 1 поздняя пахитена 1 метафаза 1 метафаза 1
- 40. Мейоз у Coprinus cinereus на уровне светового микроскопа 8 диплотена-метафаза 1 9 диакинез 10 телофаза 1
- 41. ТЭМ стадия перед слиянием стадия после слияния перед синапсисом Мейоз у Coprinus cinereus на уровне электронного
- 42. профаза 1 зиготена
- 43. профаза 1 пахитена
- 44. профаза 1 пахитена
- 45. метафаза 1
- 46. метафаза 1 анафаза 1
- 47. ранняя телофаза 1 поздняя телофаза 1
- 48. Мейоз у Agaricus bisporus
- 49. Профаза I мейоза четырехспорового штамма шампиньона Профаза I мейоза четырехспорового штамма шампиньона Лептотена Ранняя зиготена Зиготена
- 50. Запасные включения Одним из способов метаболической регуляции является резервирование большого количества запасных веществ разного состава. Основными
- 51. Были обнаружены впервые у дрожжей в конце ХIХ века и были названы гранулами волютина и метахроматина.
- 53. Полифосфаты
- 54. Распространение полифосфатов
- 55. Окрашивание полиР толуидиновым голубым с последующей дифференциацией кислым спиртом
- 57. В работах И.С.Кулаева было показано, что в каждой органелле дрожжей и других грибов присутствует своя фракция
- 58. Например, в ядре биосинтез высокомолекулярных полифосфатов каким-то образом тесно связан с биосинтезом нуклеиновых кислот, в частности
- 59. Функции полифосатов Резерв фосфора и энергии Хелатор металлов Буфер против защелачивания Участие в формировании и функции
- 60. Наиболее важный биологический эффект полифосфатов – коррекция в процессе роста и развития мицелия, ответ на стресс
- 61. Полифосфаты при исследовании с помощью электронной микроскопии
- 62. Collema, Peltigera Collema, Peltigera
- 63. Полифосфаты у микобионтов Collema (1,2) и Peltigera (5,6)
- 66. Гликоген Основной запасной полисахарид грибной клетки. Синтезируется в цитоплазме. На ультратонких срезах имеет вид мелких сферических
- 67. Запасные липиды (олеосомы) В цитоплазме могут накапливаться триацилглицерины (в молекуле глицерина образуются эфирные связи с жирными
- 68. Формирование липидной капли у Pleurotus ostreatus
- 69. Липидные включения в споре (А) и миелиноподобная структура (Б) в мицелии шампиньона двуспорового и полифосфаты в
- 71. R.Bras.Ci.Solo 2008. v.32:1531-1540 R.Bras.Ci.Solo 2008. v.32:1531-1540
- 73. Липидные капли обнаружены у большинства эукариот и некоторых прокариот LDs (lipid droplets, lipid bodies, lipid particles,
- 74. Функции липидных капель резервный источник углерода, удаление лишних белков из других компартментов для инактивации и/или запасания,
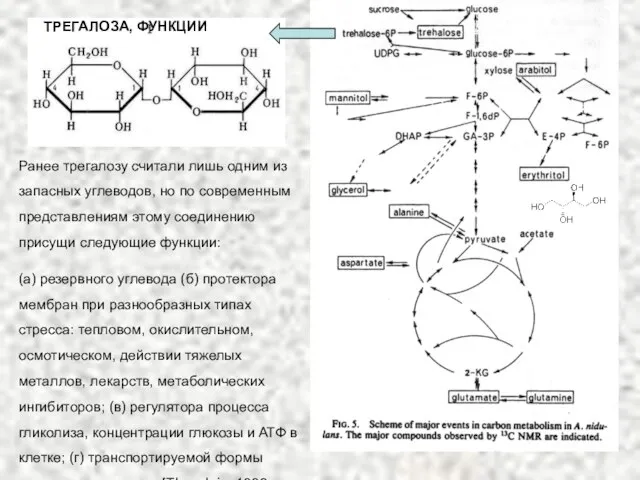
- 75. Трегалоза Важный запасной дисахарид в вегетативных клетках и в спорах грибов. Трегалоза является основным противострессорным веществом,
- 76. Ранее трегалозу считали лишь одним из запасных углеводов, но по современным представлениям этому соединению присущи следующие
- 78. Скачать презентацию

Слайд 3Наиболее изученные грибы относительно мейоза
Neurospora crassa, Sordaria fimicola, Saccharomyces cerevisiae, Schizosaccharomyces
Наиболее изученные грибы относительно мейоза
Neurospora crassa, Sordaria fimicola, Saccharomyces cerevisiae, Schizosaccharomyces

Слайд 4Преимущества мицелиальных грибов для изучения мейоза
Короткий жизненный цикл за время которого может
Преимущества мицелиальных грибов для изучения мейоза
Короткий жизненный цикл за время которого может

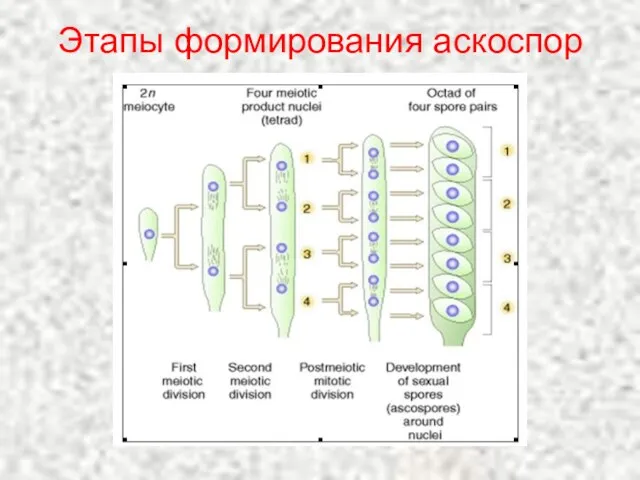
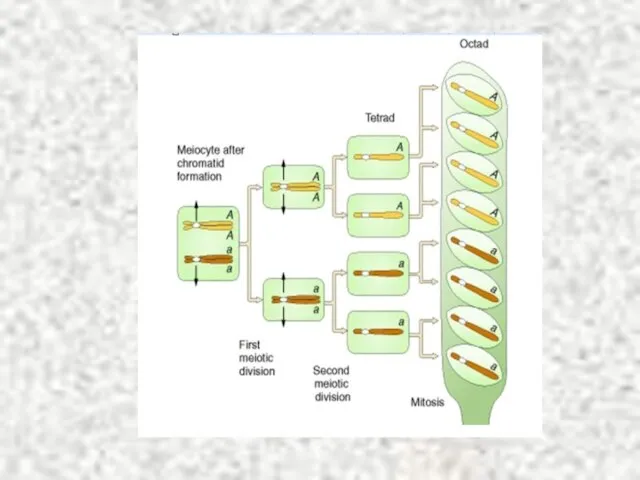
Слайд 5Этапы формирования аскоспор
Этапы формирования аскоспор
Слайд 7Этапы формирования базидий
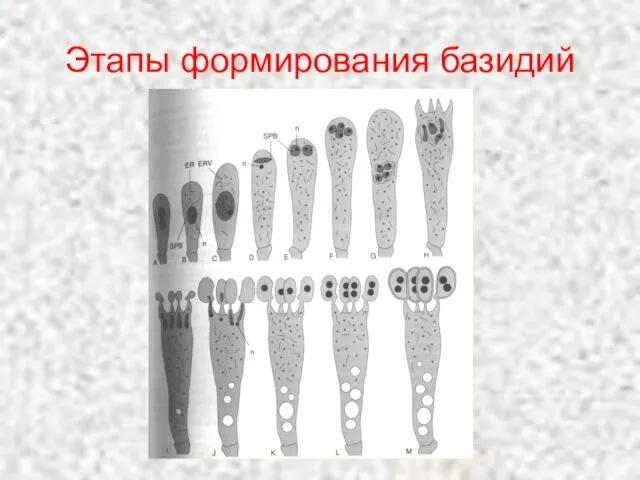
Этапы формирования базидий
Слайд 8Особенности профазы I мейоза
Профаза I мейоза – уникальная, протяженная стадия (3-10 часов)
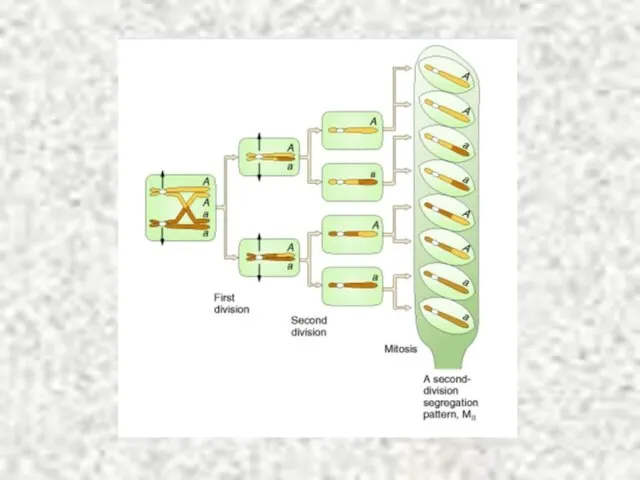
Особенности профазы I мейоза
Профаза I мейоза – уникальная, протяженная стадия (3-10 часов)

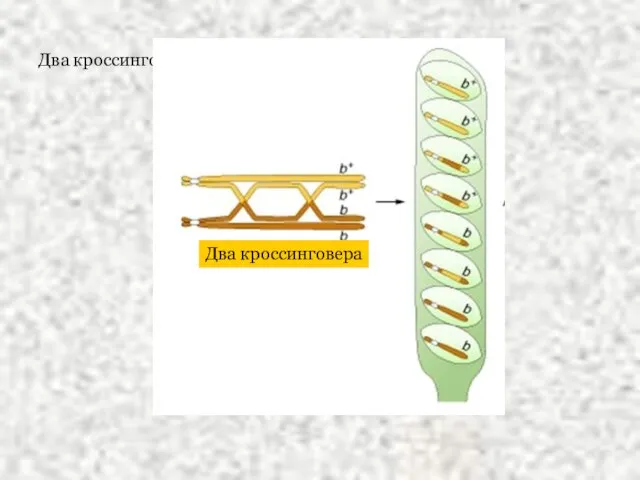
Слайд 10Два кроссинговера
Два кроссинговера
Два кроссинговера
Два кроссинговера
Слайд 11Число хромосом в ядрах разных видов грибов
Число хромосом в ядрах разных видов грибов

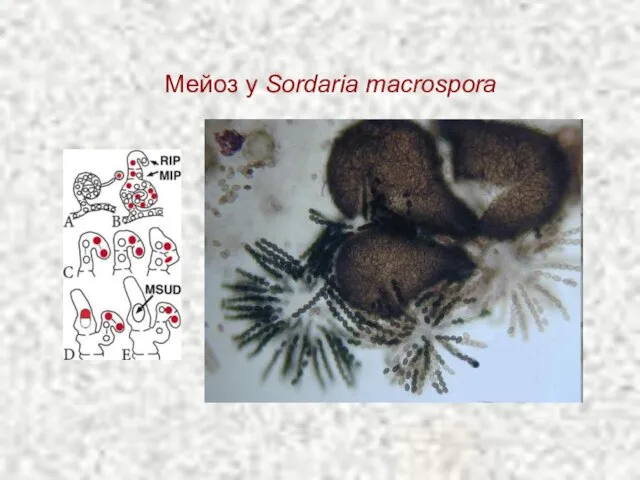
Слайд 12Мейоз у Sordaria macrospora
Мейоз у Sordaria macrospora
Слайд 13Перед кариогамией происходит удвоение ДНК (S-фаза клеточного цикла грибов).
При мейозе S-фаза длиннее,
Перед кариогамией происходит удвоение ДНК (S-фаза клеточного цикла грибов).
При мейозе S-фаза длиннее,

Слайд 14Премейотические механизмы “Checking and Cleaning”
работают перед S-фазой
Repeat-induced-point mutation (RIP) mechanism (механизм повторно
Премейотические механизмы “Checking and Cleaning”
работают перед S-фазой
Repeat-induced-point mutation (RIP) mechanism (механизм повторно

Слайд 15A “Checking” Mechanism that Operates
After Karyogamy
meiotic silencing by unpaired DNA (MSUD)
A “Checking” Mechanism that Operates
After Karyogamy
meiotic silencing by unpaired DNA (MSUD)

Слайд 16Стадии мейоза
Первое деление мейоза
профаза I
лептотена
зиготена
пахитена
диплотенa
диакинез
Стадии мейоза
Первое деление мейоза
профаза I
лептотена
зиготена
пахитена
диплотенa
диакинез

Слайд 17 Первое деление мейоза
метафаза I
анафаза I
телофаза I
Первое деление мейоза
метафаза I
анафаза I
телофаза I

Слайд 18 Второе деление мейоза
профаза II
метафаза II
анафаза II
телофаза II
Второе деление мейоза
профаза II
метафаза II
анафаза II
телофаза II

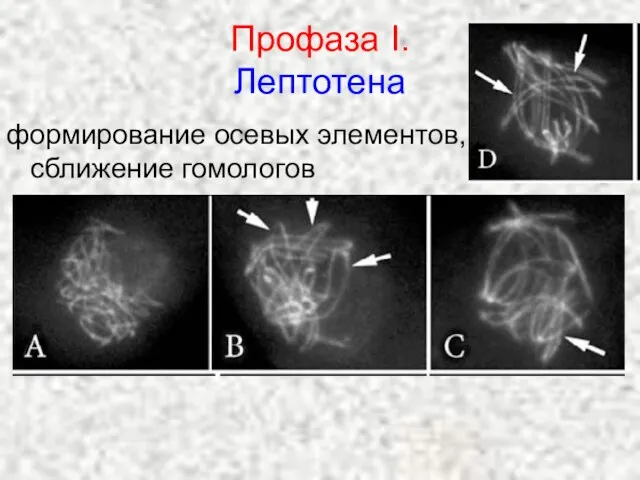
Слайд 19Профаза I.
Лептотена
формирование осевых элементов, сближение гомологов
Профаза I.
Лептотена
формирование осевых элементов, сближение гомологов
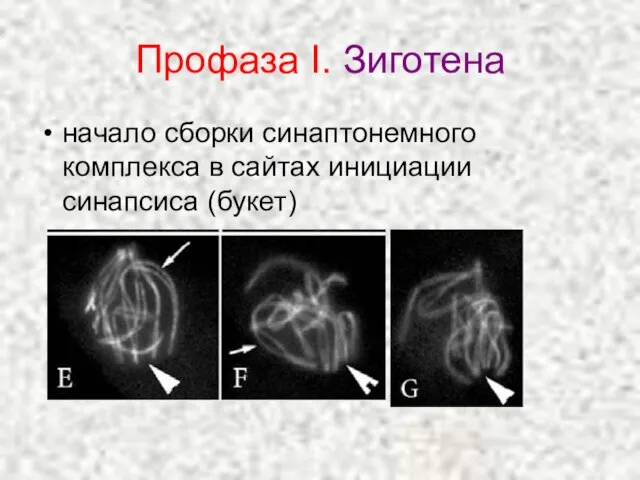
Слайд 20Профаза I. Зиготена
начало сборки синаптонемного комплекса в сайтах инициации синапсиса (букет)
Профаза I. Зиготена
начало сборки синаптонемного комплекса в сайтах инициации синапсиса (букет)
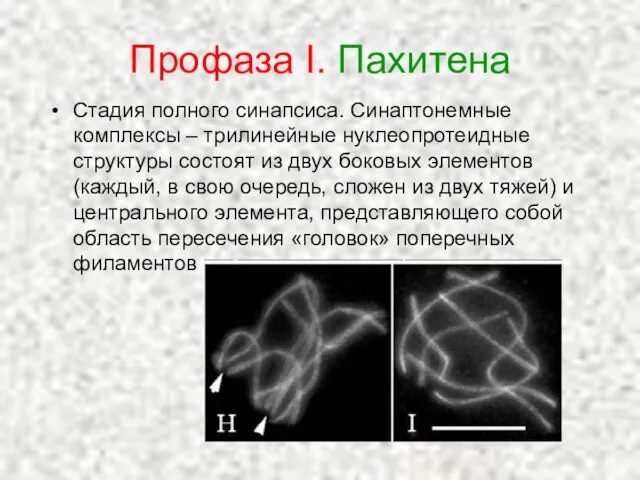
Слайд 21Профаза I. Пахитена
Стадия полного синапсиса. Синаптонемные комплексы – трилинейные нуклеопротеидные структуры состоят
Профаза I. Пахитена
Стадия полного синапсиса. Синаптонемные комплексы – трилинейные нуклеопротеидные структуры состоят
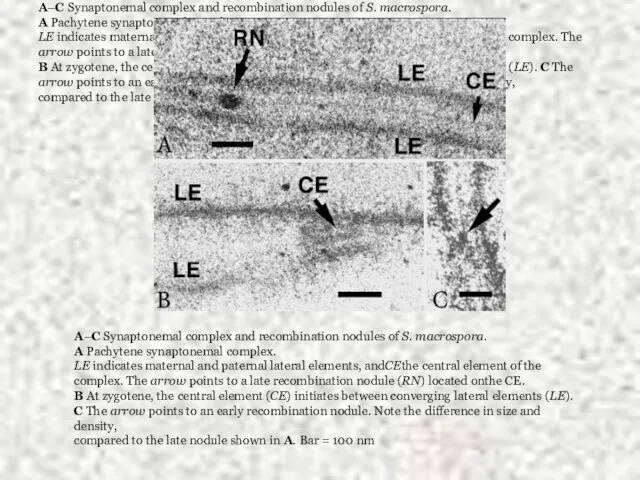
Слайд 22A–C Synaptonemal complex and recombination nodules of S. macrospora.
A Pachytene synaptonemal
A–C Synaptonemal complex and recombination nodules of S. macrospora.
A Pachytene synaptonemal
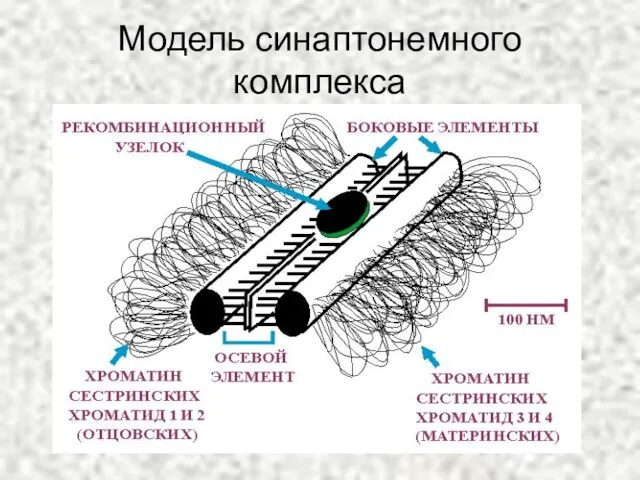
Слайд 23Модель синаптонемного комплекса
Модель синаптонемного комплекса
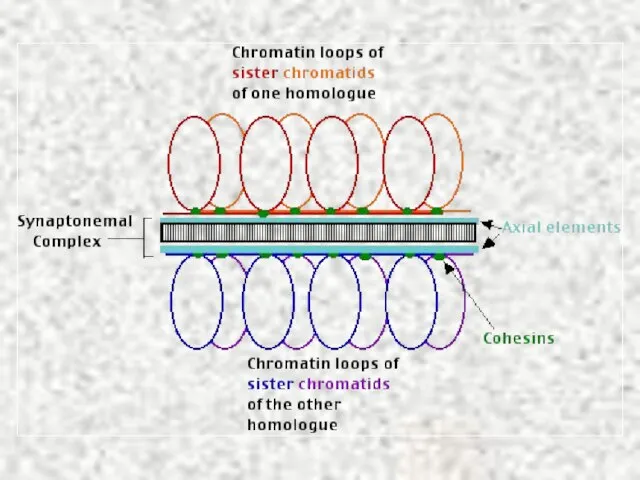
Слайд 25Профаза I. Диплотена, диакинез
Десинапсис гомологов, разборка синаптонемного комплекса. В конце концов гомологи
Профаза I. Диплотена, диакинез
Десинапсис гомологов, разборка синаптонемного комплекса. В конце концов гомологи

Слайд 26Мейоз у Saccharomyces cerevisiae
Мейоз у Saccharomyces cerevisiae
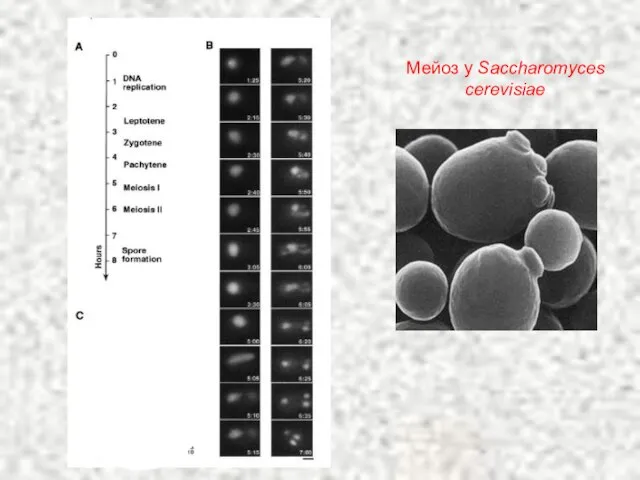
Слайд 27Мейоз у Saccharomyces cerevisiae
Мейоз у Saccharomyces cerevisiae
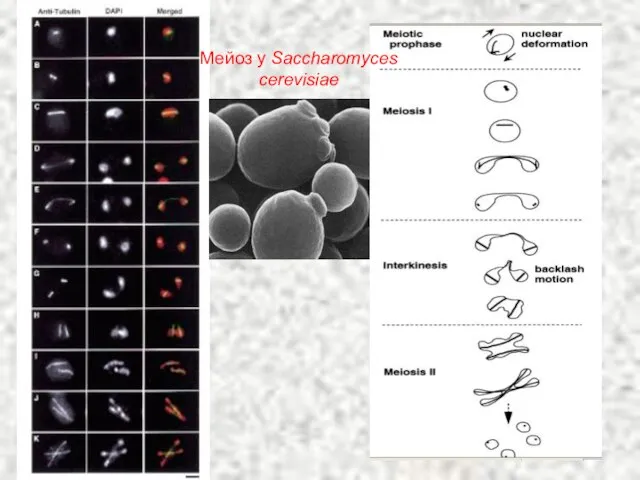
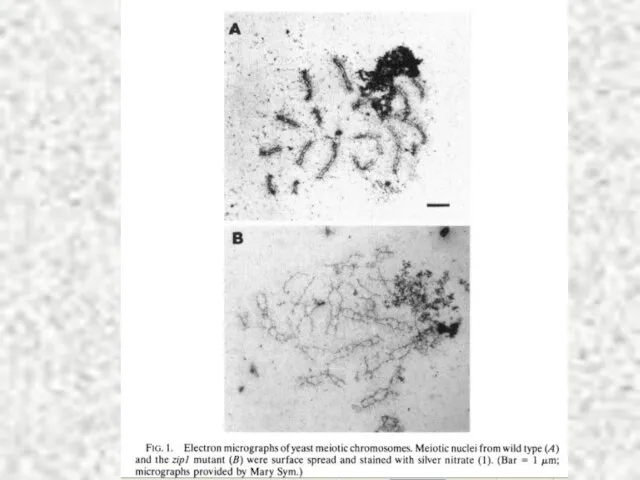
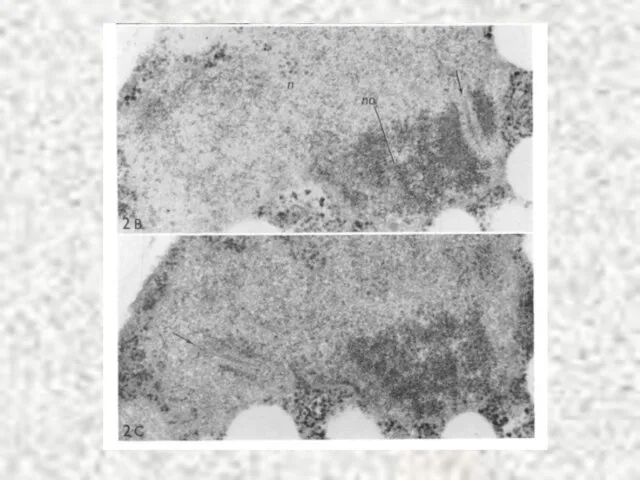
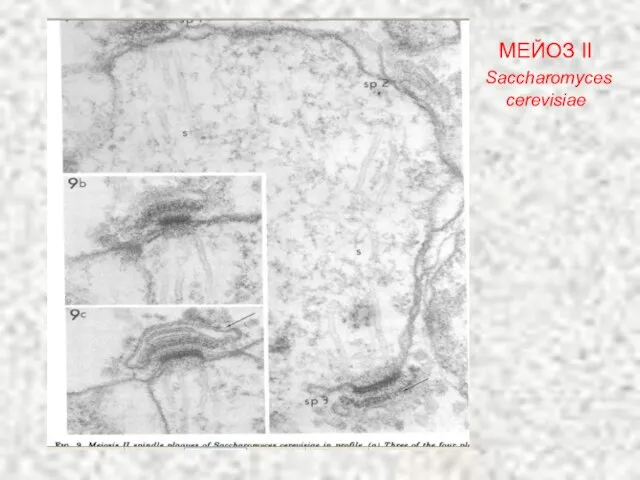
Слайд 30МЕЙОЗ II
Saccharomyces cerevisiae
МЕЙОЗ II
Saccharomyces cerevisiae
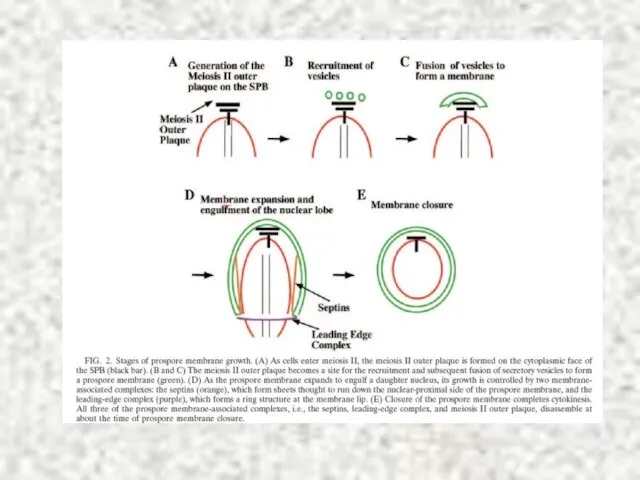
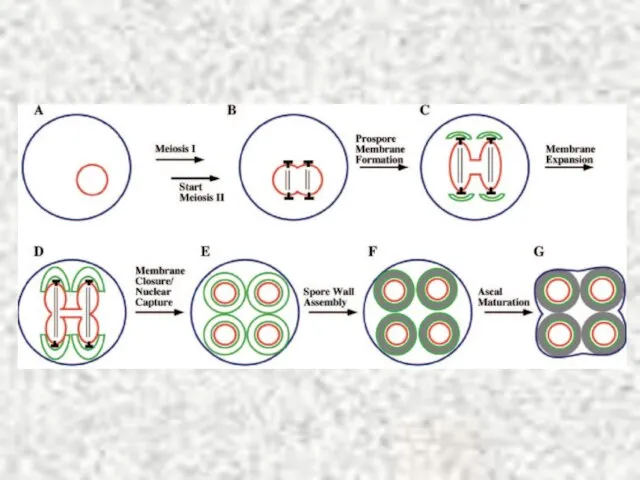
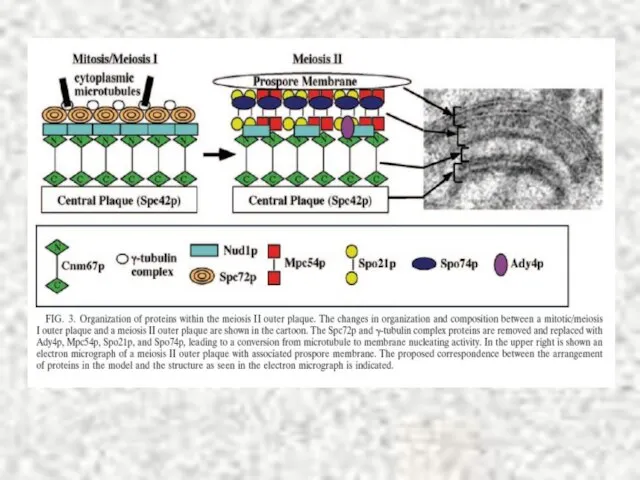
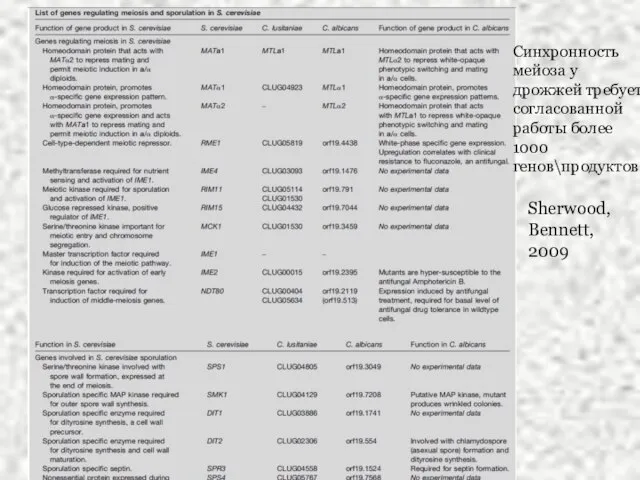
Слайд 34Sherwood, Bennett, 2009
Sherwood, Bennett, 2009
Синхронность мейоза у дрожжей требует согласованной работы более
Sherwood, Bennett, 2009
Sherwood, Bennett, 2009
Синхронность мейоза у дрожжей требует согласованной работы более
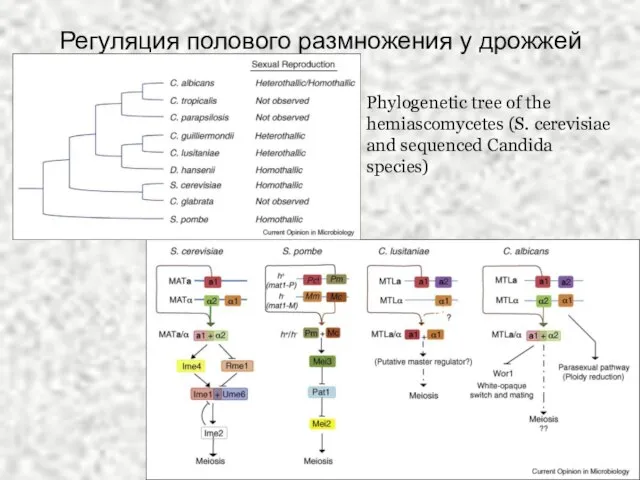
Слайд 35Регуляция полового размножения у дрожжей
Phylogenetic tree of the hemiascomycetes (S. cerevisiae and
Регуляция полового размножения у дрожжей
Phylogenetic tree of the hemiascomycetes (S. cerevisiae and


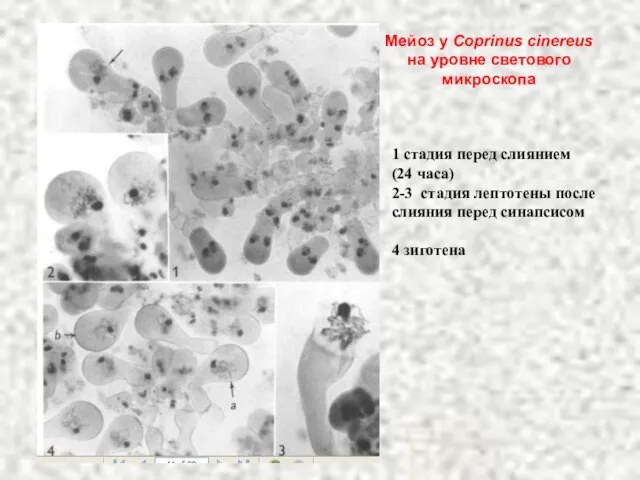
Слайд 381 стадия перед слиянием
(24 часа)
2-3 стадия лептотены после слияния перед синапсисом
4 зиготена
Мейоз
1 стадия перед слиянием
(24 часа)
2-3 стадия лептотены после слияния перед синапсисом
4 зиготена
Мейоз
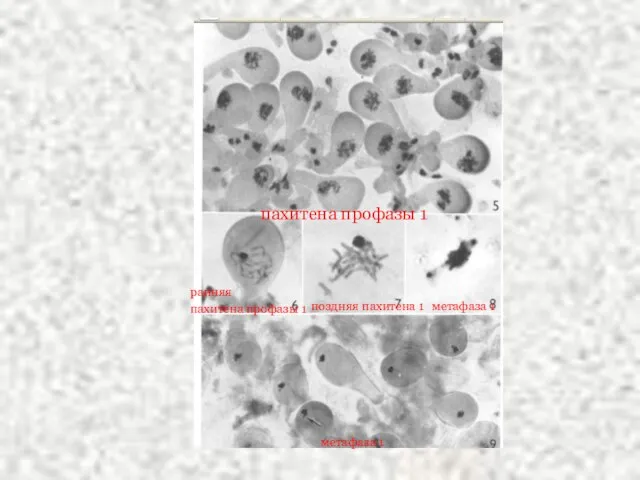
Слайд 39пахитена профазы 1
ранняя
пахитена профазы 1
поздняя пахитена 1
метафаза 1
метафаза 1
пахитена профазы 1
ранняя
пахитена профазы 1
поздняя пахитена 1
метафаза 1
метафаза 1
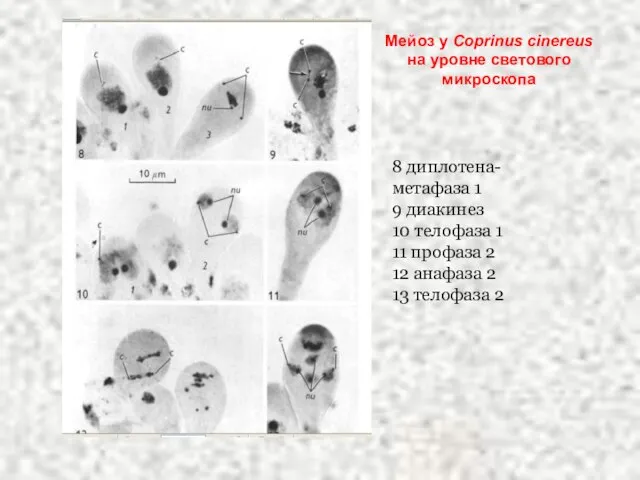
Слайд 40Мейоз у Coprinus cinereus на уровне светового микроскопа
8 диплотена-метафаза 1
9 диакинез
10 телофаза
Мейоз у Coprinus cinereus на уровне светового микроскопа
8 диплотена-метафаза 1
9 диакинез
10 телофаза
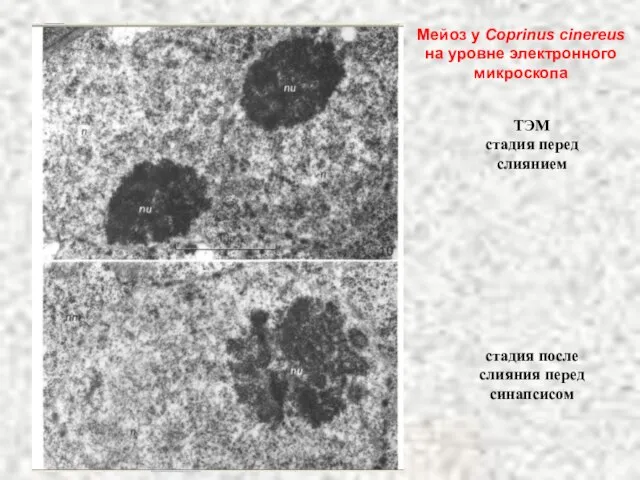
Слайд 41ТЭМ
стадия перед слиянием
стадия после слияния перед синапсисом
Мейоз у Coprinus cinereus на
ТЭМ
стадия перед слиянием
стадия после слияния перед синапсисом
Мейоз у Coprinus cinereus на
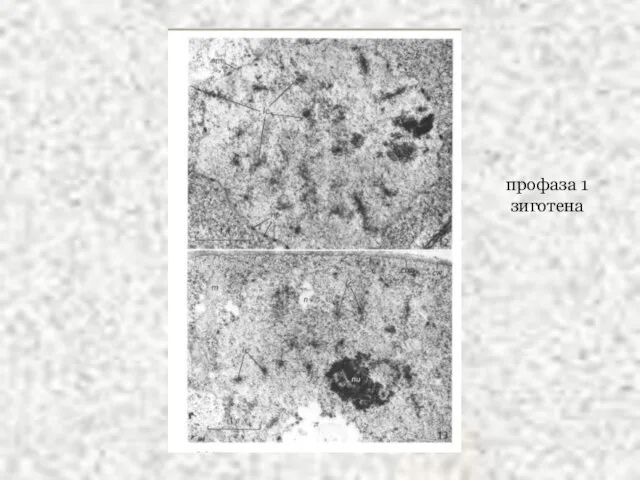
Слайд 42профаза 1
зиготена
профаза 1
зиготена
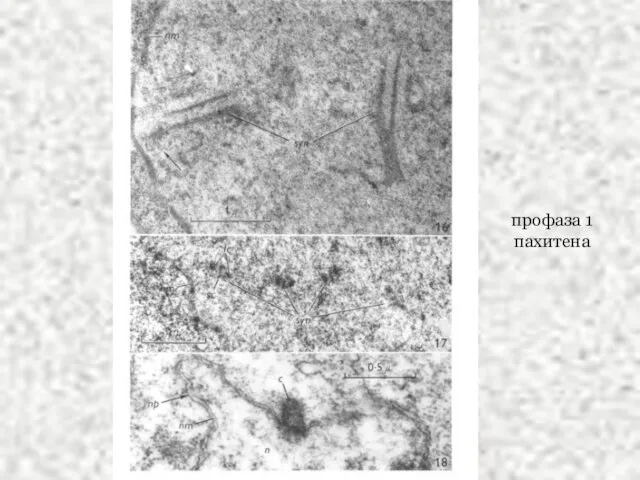
Слайд 43профаза 1
пахитена
профаза 1
пахитена

Слайд 44профаза 1
пахитена
профаза 1
пахитена
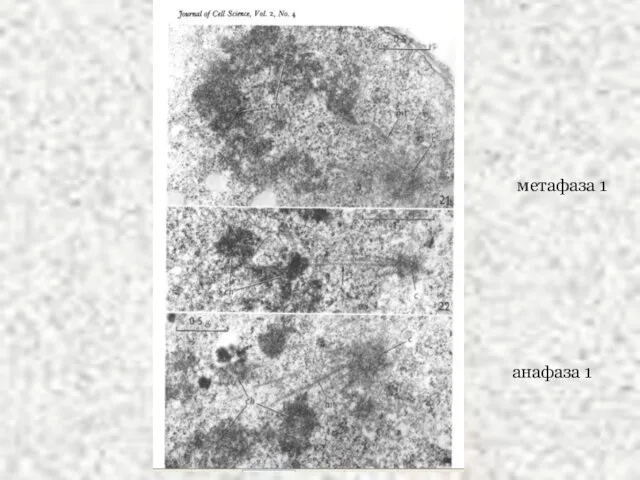
Слайд 45метафаза 1
метафаза 1
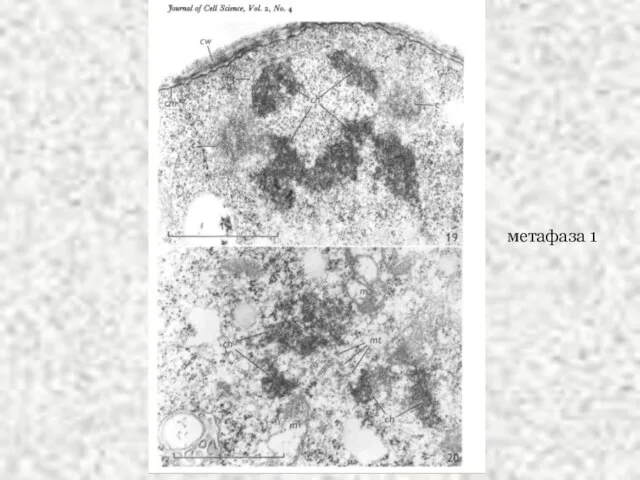
Слайд 46метафаза 1
анафаза 1
метафаза 1
анафаза 1
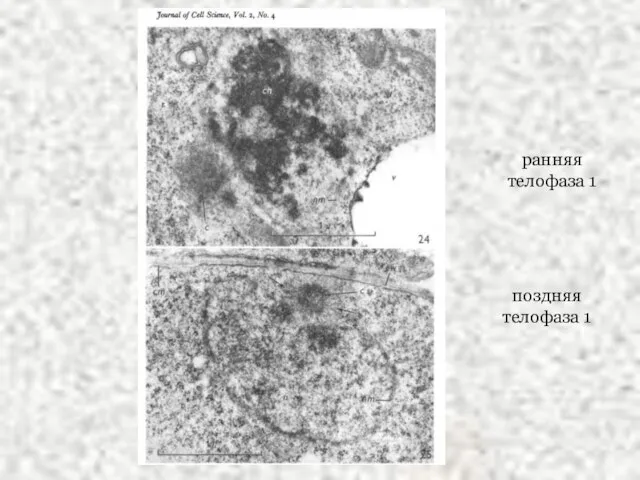
Слайд 47ранняя телофаза 1
поздняя телофаза 1
ранняя телофаза 1
поздняя телофаза 1
Слайд 48Мейоз у Agaricus bisporus
Мейоз у Agaricus bisporus

Слайд 49Профаза I мейоза четырехспорового штамма шампиньона
Профаза I мейоза четырехспорового штамма шампиньона
Лептотена
Ранняя зиготена
Зиготена
Пахитена
Пахитена
Поздняя
Профаза I мейоза четырехспорового штамма шампиньона
Профаза I мейоза четырехспорового штамма шампиньона
Лептотена
Ранняя зиготена
Зиготена
Пахитена
Пахитена
Поздняя

Слайд 50Запасные включения
Одним из способов метаболической регуляции является резервирование большого количества запасных веществ
Запасные включения
Одним из способов метаболической регуляции является резервирование большого количества запасных веществ

Слайд 51Были обнаружены впервые у дрожжей в конце ХIХ века и были названы
Были обнаружены впервые у дрожжей в конце ХIХ века и были названы


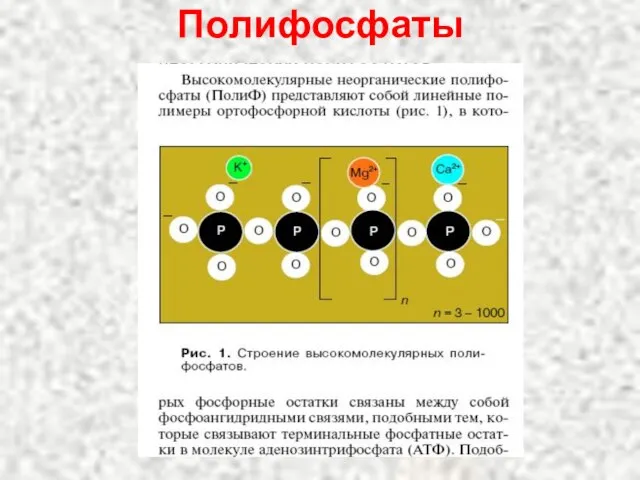
Слайд 53Полифосфаты
Полифосфаты
Слайд 54Распространение полифосфатов
Распространение полифосфатов

Слайд 55Окрашивание полиР толуидиновым голубым с последующей дифференциацией кислым спиртом
Окрашивание полиР толуидиновым голубым с последующей дифференциацией кислым спиртом


Слайд 57В работах И.С.Кулаева было показано, что в каждой органелле дрожжей и других
В работах И.С.Кулаева было показано, что в каждой органелле дрожжей и других

Слайд 58Например, в ядре биосинтез высокомолекулярных полифосфатов каким-то образом тесно связан с биосинтезом
Например, в ядре биосинтез высокомолекулярных полифосфатов каким-то образом тесно связан с биосинтезом

Слайд 59Функции полифосатов
Резерв фосфора и энергии
Хелатор металлов
Буфер против защелачивания
Участие в формировании и функции
Функции полифосатов
Резерв фосфора и энергии
Хелатор металлов
Буфер против защелачивания
Участие в формировании и функции

Слайд 60Наиболее важный биологический эффект полифосфатов – коррекция в процессе роста и развития
Наиболее важный биологический эффект полифосфатов – коррекция в процессе роста и развития

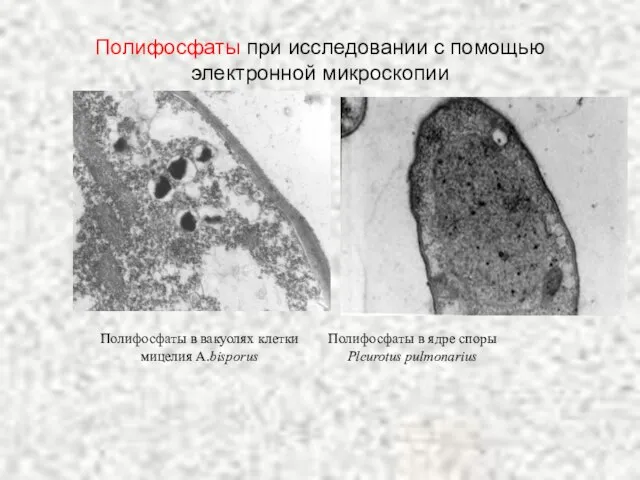
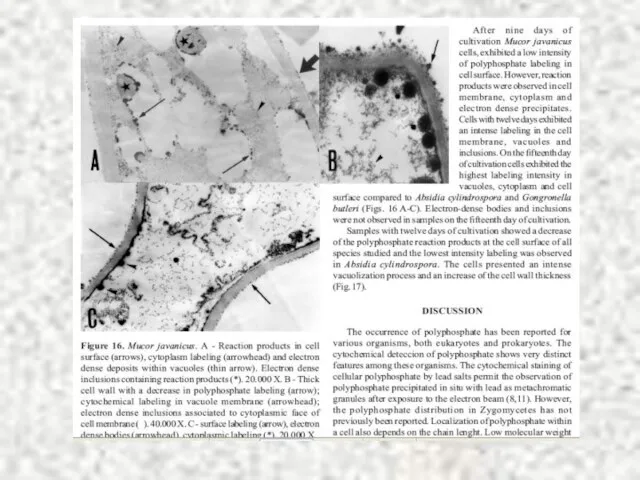
Слайд 61Полифосфаты при исследовании с помощью электронной микроскопии
Полифосфаты при исследовании с помощью электронной микроскопии
Слайд 62Collema, Peltigera
Collema, Peltigera
Collema, Peltigera
Collema, Peltigera

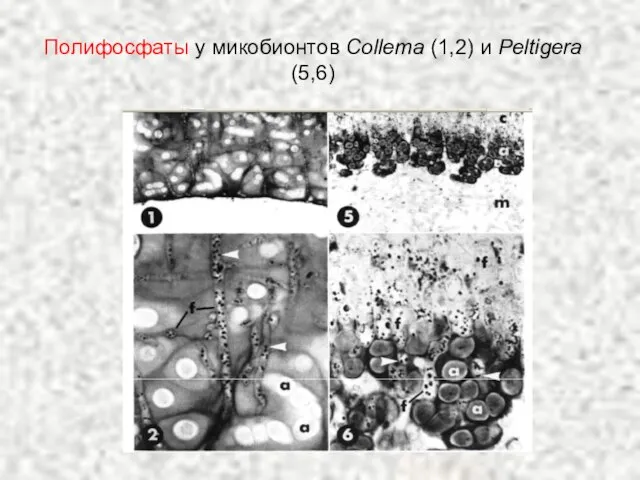
Слайд 63Полифосфаты у микобионтов Collema (1,2) и Peltigera (5,6)
Полифосфаты у микобионтов Collema (1,2) и Peltigera (5,6)

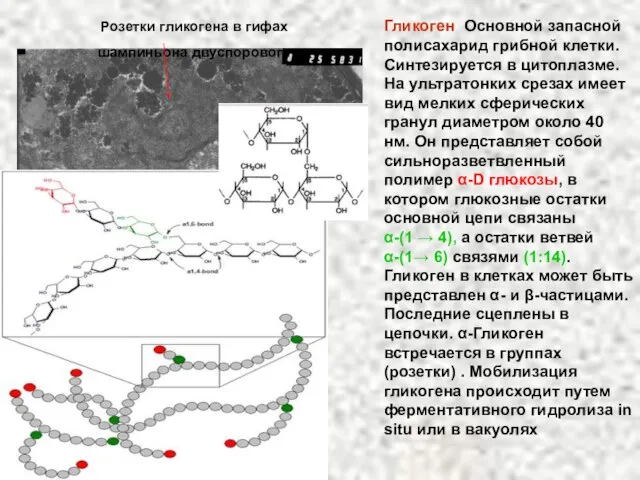
Слайд 66Гликоген Основной запасной полисахарид грибной клетки. Синтезируется в цитоплазме. На ультратонких срезах
Гликоген Основной запасной полисахарид грибной клетки. Синтезируется в цитоплазме. На ультратонких срезах
Слайд 67Запасные липиды (олеосомы)
В цитоплазме могут накапливаться триацилглицерины (в молекуле глицерина образуются эфирные
Запасные липиды (олеосомы)
В цитоплазме могут накапливаться триацилглицерины (в молекуле глицерина образуются эфирные

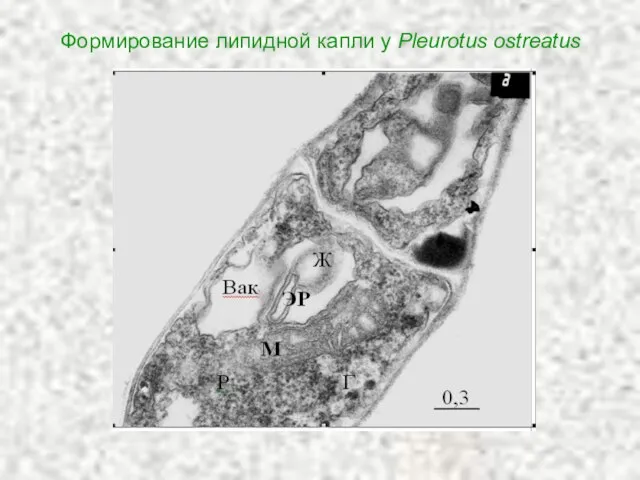
Слайд 68Формирование липидной капли у Pleurotus ostreatus
Формирование липидной капли у Pleurotus ostreatus
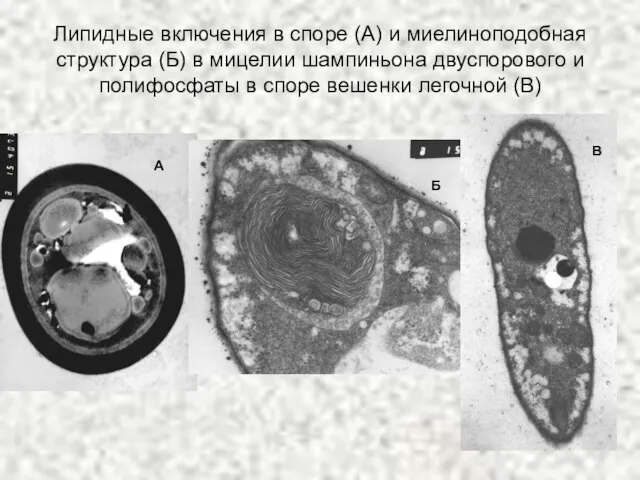
Слайд 69Липидные включения в споре (А) и миелиноподобная структура (Б) в мицелии шампиньона
Липидные включения в споре (А) и миелиноподобная структура (Б) в мицелии шампиньона
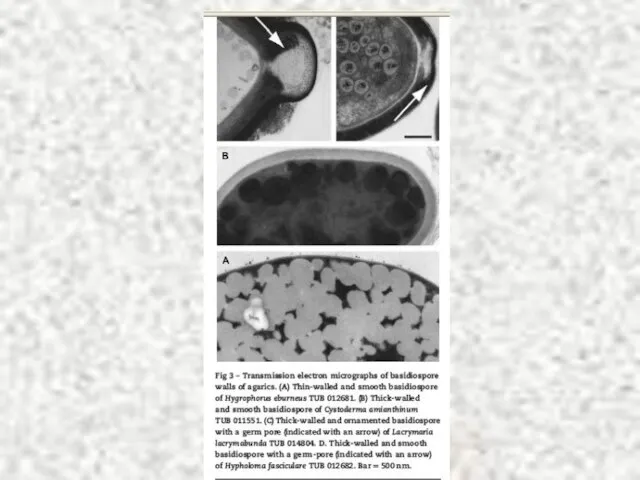
Слайд 71R.Bras.Ci.Solo 2008. v.32:1531-1540
R.Bras.Ci.Solo 2008. v.32:1531-1540
R.Bras.Ci.Solo 2008. v.32:1531-1540
R.Bras.Ci.Solo 2008. v.32:1531-1540

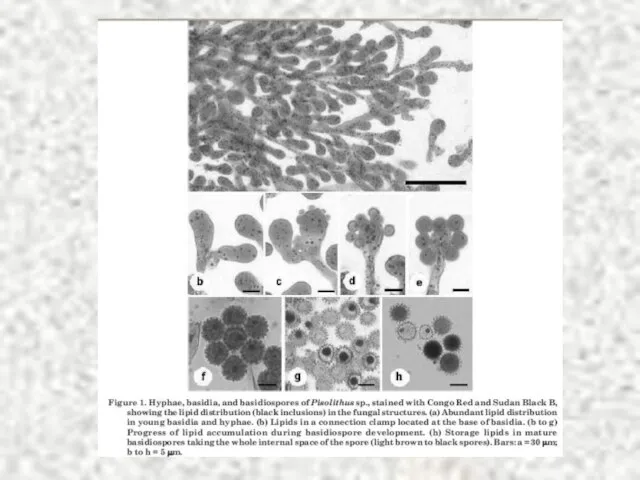
Слайд 73Липидные капли обнаружены у большинства эукариот и некоторых прокариот LDs (lipid droplets,
Липидные капли обнаружены у большинства эукариот и некоторых прокариот LDs (lipid droplets,

Слайд 74Функции липидных капель
резервный источник углерода,
удаление лишних белков из других компартментов для
Функции липидных капель
резервный источник углерода,
удаление лишних белков из других компартментов для

Слайд 75Трегалоза
Важный запасной дисахарид в вегетативных клетках и в спорах грибов.
Трегалоза является основным
Трегалоза
Важный запасной дисахарид в вегетативных клетках и в спорах грибов.
Трегалоза является основным

Слайд 76Ранее трегалозу считали лишь одним из запасных углеводов, но по современным представлениям
Ранее трегалозу считали лишь одним из запасных углеводов, но по современным представлениям
 Мини - волейбол. Игра по упращенным правилам
Мини - волейбол. Игра по упращенным правилам Страхование ВЭД
Страхование ВЭД Непрерывное медицинское и фармацевтическое образование
Непрерывное медицинское и фармацевтическое образование Презентация на тему Франц Шуберт
Презентация на тему Франц Шуберт  Аномия и девиантное поведение
Аномия и девиантное поведение Додавання і віднімання дробів з різними знаменниками
Додавання і віднімання дробів з різними знаменниками Исаак Ильич Левитан
Исаак Ильич Левитан Магазин музыкальных инструментов Rock Line Magazi
Магазин музыкальных инструментов Rock Line Magazi Веганская уха
Веганская уха Реклама - распространяемая в любой форме, с помощью любых средств информация о физическом или юридическом лице, товарах, идеях и н
Реклама - распространяемая в любой форме, с помощью любых средств информация о физическом или юридическом лице, товарах, идеях и н Пещерные храмы Китая
Пещерные храмы Китая Контекстная реклама МетаКонтекст
Контекстная реклама МетаКонтекст Исследование почвы на наличие в ней тяжелых металлов химическим путем
Исследование почвы на наличие в ней тяжелых металлов химическим путем Носители информации
Носители информации Великий, могучий русский язык. Агитационная акция С книгой в 21 век
Великий, могучий русский язык. Агитационная акция С книгой в 21 век Монголо-татары
Монголо-татары Население
Население Масиви Object Delphi
Масиви Object Delphi “Природоохранные технологии” ООО. Варианты размещения КНС 4200 м3/сут мкр.Купелинка
“Природоохранные технологии” ООО. Варианты размещения КНС 4200 м3/сут мкр.Купелинка Устройство детей-сирот, детей, оставшихся без попечения родителей, в семьи на территории Пермского края
Устройство детей-сирот, детей, оставшихся без попечения родителей, в семьи на территории Пермского края 技术和科学的新形式在因特网的作用, 互联网实际应用 以及建设领域和科学知识的传播和流行 Применение Интернета в распространении научно-популярных знаний Лекция №6
技术和科学的新形式在因特网的作用, 互联网实际应用 以及建设领域和科学知识的传播和流行 Применение Интернета в распространении научно-популярных знаний Лекция №6 Три состояния воды
Три состояния воды Презентация на тему Англия и Шотландия
Презентация на тему Англия и Шотландия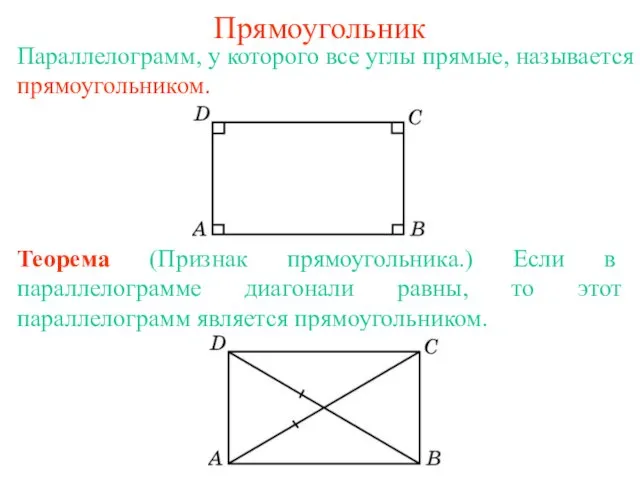 Прямоугольник
Прямоугольник Методы планирования. Экстраполяция. Балансовый метод. Методы календарного планирования
Методы планирования. Экстраполяция. Балансовый метод. Методы календарного планирования Федеральный законодательный процесс
Федеральный законодательный процесс Spot wizerunkowy
Spot wizerunkowy Тема урока:
Тема урока: