- МОЛЕКУЛЯРНОЕ СТРОЕНИЕ ОПЕРОНА

Содержание
- 2. 1. Генетическая система координат. Строение оператора и промотора Методами молекулярной биологии в 70 х годах была
- 3. Строение оперона Рис. 1. Молекулярное строение и схема экспрессии оперона: а — принципиальная структура гена (р
- 4. На рисунке 1 использована принятая в молекулярной гене-тике система координат, в которой нуклеотиды нумеруются от границы
- 5. В состав промоторов всех изученных оперонов входят два участка с относительно постоянным составом и последовательностью нуклеотидов
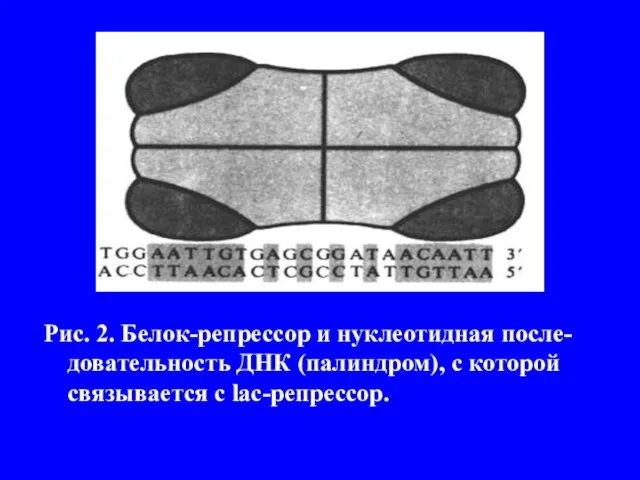
- 6. Последовательность нуклеотидов оператора (область ДНК от +1 до +24). лактозного оперона (Lac-оперона) образует симметричный палиндром (рис.
- 7. Рис. 2. Белок-репрессор и нуклеотидная после-довательность ДНК (палиндром), с которой связывается с lac-репрессор.
- 8. 2. Структура транскрибируемой области оперона. Терминация транскрипции. Результат транскрипции Транскрибируемая область оперона включает не только опе-ратор
- 9. Примерно с 37-го нуклеотида начинается ДНК первого структурного гена. Первый триплет (кодон в мРНК) называется инициирующим
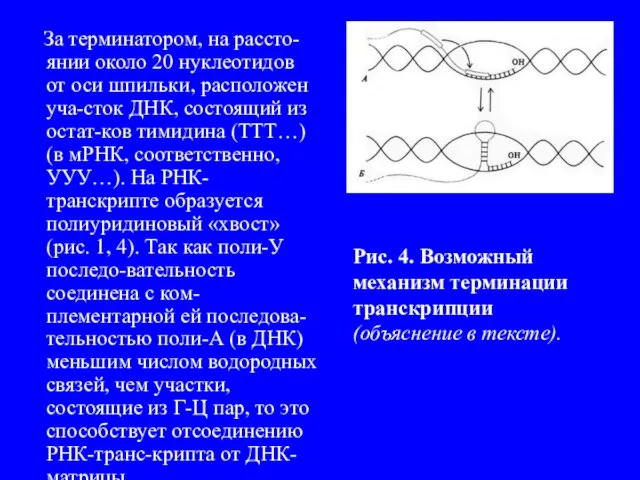
- 10. Предполагается, что образование шпилек в синтезированной моле-куле мРНК способствует её отсо-единению от ДНК-матрицы. Движение РНК-полимеразы по
- 11. За терминатором, на рассто-янии около 20 нуклеотидов от оси шпильки, расположен уча-сток ДНК, состоящий из остат-ков
- 12. На рис. 5 показана схема экспрессии лактозного оперона E.coli. Образовавша-яся в ходе транс-крипции полигенная (полицистронная) молекула
- 13. Трансляция мРНК начинается с посадки рибосом на мРНК, которая осуществляется благодаря комплементарности 3’–конца 16S рибосомной РНК
- 14. 3. Регулоны и аттенюаторы В ДНК бактерий существует и другой тип организа-ции генов. Некоторые группы структурных
- 15. Аттенюатор – это нуклеотидная последовательность с инвертированными повторами, расположенная между промотором и первым геном оперона. Вторичная
- 16. 4. Молекулярный механизм работы белка-репрессора Молекула активного Lac-репрессора это тетрамер, постро-енный из четырёх одинаковых полипептидных цепей
- 17. Удивительна точность, с которой молекула репрессора «опознаёт» операторный участок. Длина молекулы ДНК E. coli составляет 3,2
- 18. Она достаточно сильна, чтобы удерживать молекулы репрес-сора в постоянном контакте с нитью ДНК, но позволяет им
- 19. Поиск оператора молекулами репрессора облегчается благодаря особенностям строения как самого репрессора, так и операторного участка ДНК.
- 21. Скачать презентацию
Слайд 21. Генетическая система координат. Строение оператора и промотора
Методами молекулярной биологии в
1. Генетическая система координат. Строение оператора и промотора
Методами молекулярной биологии в

Слайд 3Строение оперона
Рис. 1. Молекулярное строение и схема экспрессии оперона: а — принципиальная
Строение оперона
Рис. 1. Молекулярное строение и схема экспрессии оперона: а — принципиальная
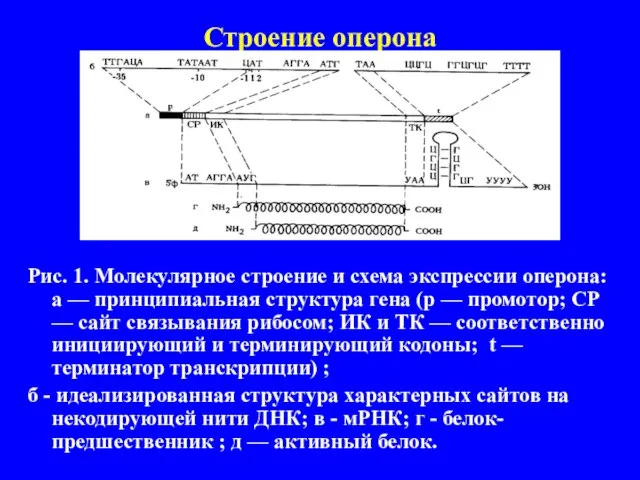
Слайд 4 На рисунке 1 использована принятая в молекулярной гене-тике система координат, в
На рисунке 1 использована принятая в молекулярной гене-тике система координат, в

Слайд 5В состав промоторов всех изученных оперонов входят два участка с относительно постоянным
В состав промоторов всех изученных оперонов входят два участка с относительно постоянным

Слайд 6 Последовательность нуклеотидов оператора (область ДНК от +1 до +24). лактозного оперона
Последовательность нуклеотидов оператора (область ДНК от +1 до +24). лактозного оперона

Слайд 7Рис. 2. Белок-репрессор и нуклеотидная после-довательность ДНК (палиндром), с которой связывается с
Рис. 2. Белок-репрессор и нуклеотидная после-довательность ДНК (палиндром), с которой связывается с
Слайд 82. Структура транскрибируемой области оперона. Терминация транскрипции. Результат транскрипции
Транскрибируемая область оперона
2. Структура транскрибируемой области оперона. Терминация транскрипции. Результат транскрипции
Транскрибируемая область оперона

Слайд 9 Примерно с 37-го нуклеотида начинается ДНК первого структурного гена. Первый триплет
Примерно с 37-го нуклеотида начинается ДНК первого структурного гена. Первый триплет

Слайд 10Предполагается, что образование шпилек в синтезированной моле-куле мРНК способствует её отсо-единению от
Предполагается, что образование шпилек в синтезированной моле-куле мРНК способствует её отсо-единению от

Слайд 11 За терминатором, на рассто-янии около 20 нуклеотидов от оси шпильки, расположен
За терминатором, на рассто-янии около 20 нуклеотидов от оси шпильки, расположен
Слайд 12 На рис. 5 показана схема экспрессии лактозного оперона E.coli. Образовавша-яся в
На рис. 5 показана схема экспрессии лактозного оперона E.coli. Образовавша-яся в

Слайд 13 Трансляция мРНК начинается с посадки рибосом на мРНК, которая осуществляется благодаря
Трансляция мРНК начинается с посадки рибосом на мРНК, которая осуществляется благодаря

Слайд 143. Регулоны и аттенюаторы
В ДНК бактерий существует и другой тип организа-ции
3. Регулоны и аттенюаторы
В ДНК бактерий существует и другой тип организа-ции

Слайд 15 Аттенюатор – это нуклеотидная последовательность с инвертированными повторами, расположенная между промотором
Аттенюатор – это нуклеотидная последовательность с инвертированными повторами, расположенная между промотором

Слайд 164. Молекулярный механизм работы белка-репрессора
Молекула активного Lac-репрессора это тетрамер, постро-енный
4. Молекулярный механизм работы белка-репрессора
Молекула активного Lac-репрессора это тетрамер, постро-енный

Слайд 17Удивительна точность, с которой молекула репрессора «опознаёт» операторный участок. Длина молекулы ДНК
Удивительна точность, с которой молекула репрессора «опознаёт» операторный участок. Длина молекулы ДНК

Слайд 18Она достаточно сильна, чтобы удерживать молекулы репрес-сора в постоянном контакте с нитью
Она достаточно сильна, чтобы удерживать молекулы репрес-сора в постоянном контакте с нитью
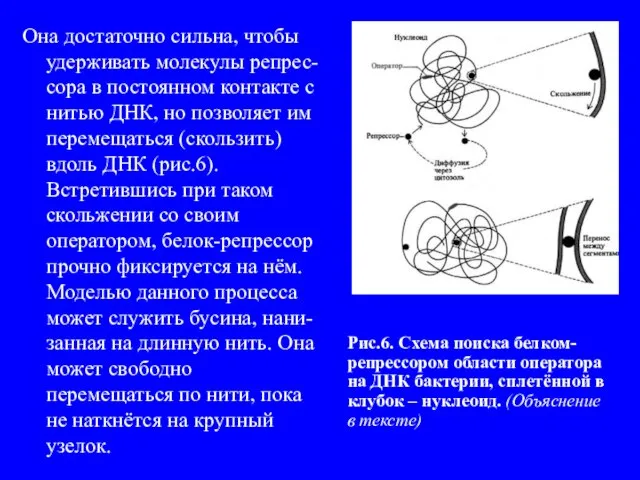
Слайд 19Поиск оператора молекулами репрессора облегчается благодаря особенностям строения как самого репрессора, так
Поиск оператора молекулами репрессора облегчается благодаря особенностям строения как самого репрессора, так

 Заключительный урок по теме «Жизнь и творчество Владимира Маяковского»
Заключительный урок по теме «Жизнь и творчество Владимира Маяковского» «Социализация обучающихся через предпрофильную подготовку»
«Социализация обучающихся через предпрофильную подготовку» Знакомьтесь: фотобанки (микростоки, фотостоки)
Знакомьтесь: фотобанки (микростоки, фотостоки) Тест по теме Строительство
Тест по теме Строительство «Современные образовательные технологии в преподавании географии»
«Современные образовательные технологии в преподавании географии» Станция «Детская библиотека»
Станция «Детская библиотека» История китайской живописи
История китайской живописи МЕЖДУНАРОДНЫЙ НАЛОГОВЫЙ КОНГРЕСС Заемное финансирование – практические аспекты Контролируемая задолженность: понятие и сложн
МЕЖДУНАРОДНЫЙ НАЛОГОВЫЙ КОНГРЕСС Заемное финансирование – практические аспекты Контролируемая задолженность: понятие и сложн Качественное образование и успешная профессиональная карьера
Качественное образование и успешная профессиональная карьера ПАКТУ РЕРИХА – 70 лет!
ПАКТУ РЕРИХА – 70 лет! Людмила Гаврилюк-Єнсен координатор проекту в ТНЕУ Канадсько-український проект «Регіональне навчання та розвиток консультативни
Людмила Гаврилюк-Єнсен координатор проекту в ТНЕУ Канадсько-український проект «Регіональне навчання та розвиток консультативни Презентация на тему Общая характеристика ИНДИИ
Презентация на тему Общая характеристика ИНДИИ Презентация на тему Морские рыбы
Презентация на тему Морские рыбы  Исследовательский проект на тему:
Исследовательский проект на тему: Основные понятия теории менеджмента
Основные понятия теории менеджмента Математика в футболе.Презентация подготовлена учеником 10 класса завазальским а.
Математика в футболе.Презентация подготовлена учеником 10 класса завазальским а. хаврошечка
хаврошечка Сертификат об участии в Региональном семинаре для судей по конькобежному спорту
Сертификат об участии в Региональном семинаре для судей по конькобежному спорту Guzenko_Vladislav_Ibaskf-191
Guzenko_Vladislav_Ibaskf-191 Вирус сквернословия
Вирус сквернословия Физическая культура и закаливание
Физическая культура и закаливание Доходность euro / usd
Доходность euro / usd Презентация на тему Моя Россия
Презентация на тему Моя Россия 15 Unusual Places You Won’t Believe Existed In India
15 Unusual Places You Won’t Believe Existed In India К здоровой семье через детский сад
К здоровой семье через детский сад Громко прозвенел звонок - начинается урок
Громко прозвенел звонок - начинается урок История Кубани. Краткий обзор.
История Кубани. Краткий обзор. Альтернативная коммуникация
Альтернативная коммуникация