- МОЛЕКУЛЯРНЫЕ АСПЕКТЫ ПЕРЕДАЧИ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ

Содержание
- 2. Третичная структура ДНК: Начальный элемент упаковки – нуклеосома: сердцевину ее составляет октамер, состоящий из 4 димерных
- 3. Нуклеиновые кислоты и белки называют информационными молекулами, так как в чередовании их мономеров заложен определенный смысл.
- 4. Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности
- 5. РЕПЛИКАЦИЯ – процесс удвоения ДНК, суть которого состоит в образовании идентичных копий ДНК для передачи генетической
- 6. 1 - топоизомераза (гираза) - обратимая нуклеаза, сначала она разрывает цепь ДНК, а по окончании репликации
- 7. В настоящее время процесс репликации у прокариот достаточно изучен, в то время как многие аспекты эукариотической
- 8. Движение хеликазы усиливает скрученность (супер-спирализацию) впереди нее. Убирают образующиеся супервитки ферменты топоизомеразы. Они разрывают фосфатную перемычку
- 9. 3. Терминация. Заканчивается репликация тогда, когда вся матричная ДНК скопирована. В образованной таким образом двойной спирали
- 10. «Молекулярные часы клетки» - теломеры Особенность репликации линейных молекул эукариот связана со сложностью репликации концевых участков
- 11. РЕПАРАЦИЯ ПОВРЕЖДЕНИЙ И ОШИБОК ДНК Передача генетической информации в неискаженном виде - важнейшее условие сохранения вида.
- 12. Генные мутации Изменения генетической программы ДНК клеток называются мутациями. Различают хромосомные мутации (изменение числа хромосом, хромосомные
- 13. МОЛЕКУЛЯРНЫЕ ОСНОВЫ ТРАНСКРИПЦИИ Транскрипция - это синтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой
- 14. РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет только в направлении 5'
- 15. РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов. В промоторе узнается взаимное расположение двух AT-богатых участков. В
- 16. Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну tРНК,
- 17. На всех этапах транскрипции необходимо участие специальных белковых факторов: А, В – факторов инициации, Е, Н,
- 18. На этом этапе действует ингибитор элонгации - стрептолидигин 3. Терминация У E.coli . большинство генов терминируются
- 19. У прокариот м-РНК синтезируется сразу в зрелом виде. У эукариот все первичные транскрипты , образующиеся с
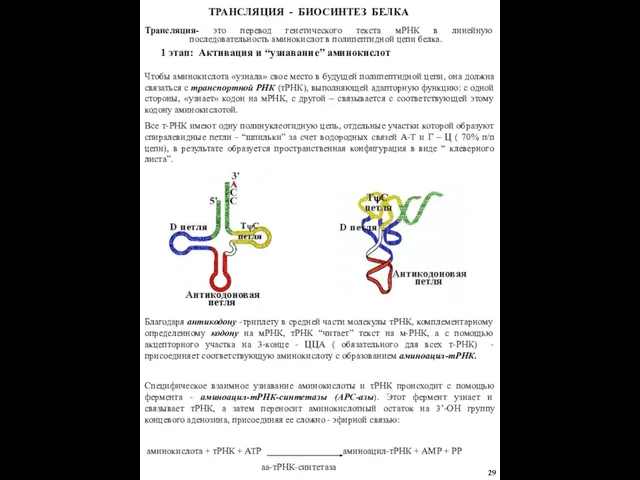
- 20. Благодаря антикодону -триплету в средней части молекулы тРНК, комплементарному определенному кодону на мРНК, тРНК “читает” текст
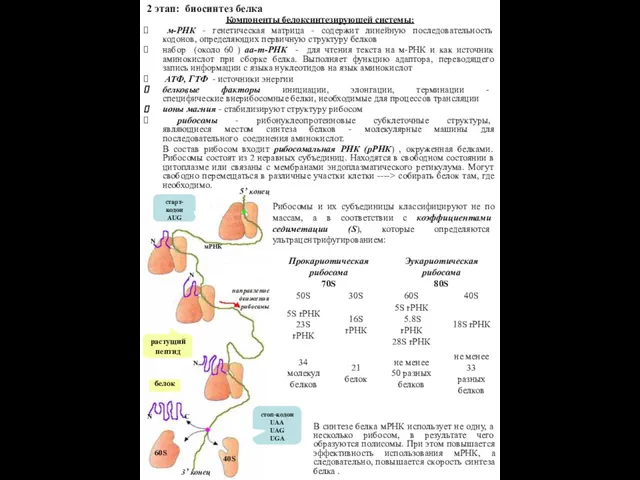
- 21. Компоненты белоксинтезирующей системы: м-РНК - генетическая матрица - содержит линейную последовательность кодонов, определяющих первичную структуру белков
- 22. 1. ИНИЦИАЦИЯ ТРАНСЛЯЦИИ На рибосоме имеются два участка: П-сайт или пептидильный сайт, служащий для связывания с
- 23. 2. ЭЛОНГАЦИЯ ТРАНСЛЯЦИИ Элонгация - это рост полипептидной цепи от N-конца к С-концу, ступенчатое удлинение синтезируемой
- 24. Во время элонгации происходит важное событие - формирование пространственной структуры полипептидной цепи, от чего в значительной
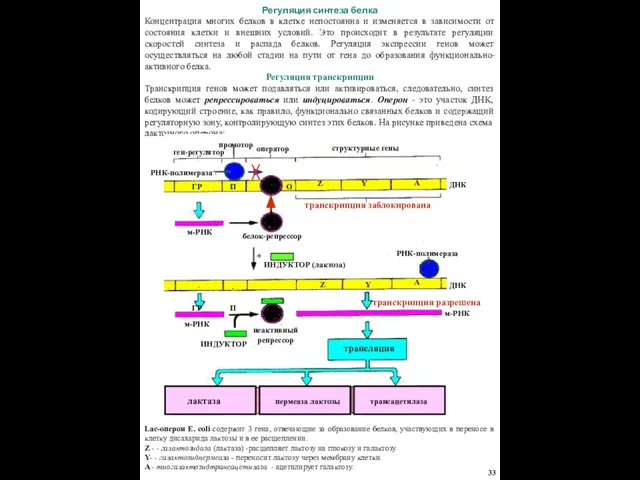
- 25. 33 Регуляция синтеза белка Концентрация многих белков в клетке непостоянна и изменяется в зависимости от состояния
- 27. Скачать презентацию
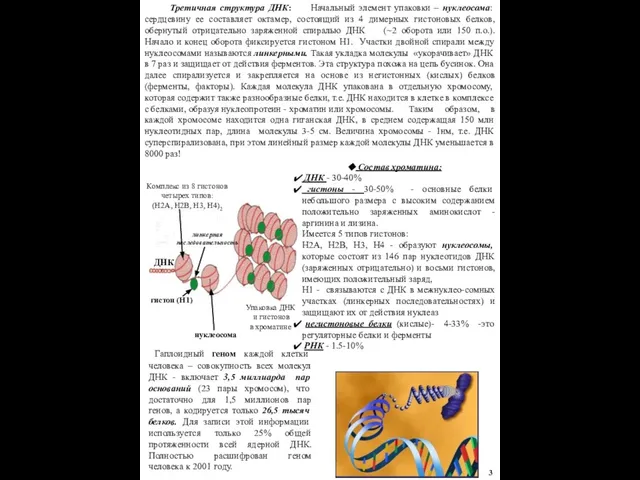
Слайд 3Нуклеиновые кислоты и белки называют информационными молекулами, так как в чередовании их
Нуклеиновые кислоты и белки называют информационными молекулами, так как в чередовании их

Слайд 4 Генетический код - это система записи информации о последовательности расположения аминокислот в
Генетический код - это система записи информации о последовательности расположения аминокислот в

Слайд 5 РЕПЛИКАЦИЯ – процесс удвоения ДНК, суть которого состоит в образовании идентичных копий
РЕПЛИКАЦИЯ – процесс удвоения ДНК, суть которого состоит в образовании идентичных копий

Слайд 6 1 - топоизомераза (гираза) - обратимая нуклеаза, сначала она разрывает цепь
1 - топоизомераза (гираза) - обратимая нуклеаза, сначала она разрывает цепь
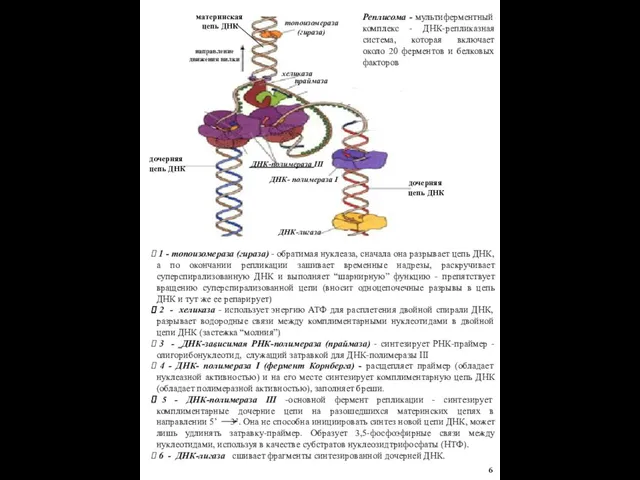
Слайд 7 В настоящее время процесс репликации у прокариот достаточно изучен, в то время
В настоящее время процесс репликации у прокариот достаточно изучен, в то время

Слайд 8Движение хеликазы усиливает скрученность (супер-спирализацию) впереди нее. Убирают образующиеся супервитки ферменты топоизомеразы.
Движение хеликазы усиливает скрученность (супер-спирализацию) впереди нее. Убирают образующиеся супервитки ферменты топоизомеразы.

Слайд 93. Терминация.
Заканчивается репликация тогда, когда вся матричная ДНК скопирована. В образованной
3. Терминация.
Заканчивается репликация тогда, когда вся матричная ДНК скопирована. В образованной

Слайд 10«Молекулярные часы клетки» - теломеры
Особенность репликации линейных молекул эукариот связана со сложностью
«Молекулярные часы клетки» - теломеры
Особенность репликации линейных молекул эукариот связана со сложностью

Слайд 11РЕПАРАЦИЯ ПОВРЕЖДЕНИЙ И ОШИБОК ДНК
Передача генетической информации в неискаженном виде - важнейшее
РЕПАРАЦИЯ ПОВРЕЖДЕНИЙ И ОШИБОК ДНК
Передача генетической информации в неискаженном виде - важнейшее

Слайд 12
Генные мутации
Изменения генетической программы ДНК клеток называются мутациями. Различают хромосомные мутации (изменение
Генные мутации
Изменения генетической программы ДНК клеток называются мутациями. Различают хромосомные мутации (изменение

Слайд 13МОЛЕКУЛЯРНЫЕ ОСНОВЫ ТРАНСКРИПЦИИ
Транскрипция - это синтез всех видов РНК по матрице ДНК,
МОЛЕКУЛЯРНЫЕ ОСНОВЫ ТРАНСКРИПЦИИ
Транскрипция - это синтез всех видов РНК по матрице ДНК,

Слайд 14РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет
РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет

Слайд 15РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов. В промоторе узнается взаимное расположение
РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов. В промоторе узнается взаимное расположение

Слайд 16Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев -
Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев -

Слайд 17 На всех этапах транскрипции необходимо участие специальных белковых факторов:
А,
На всех этапах транскрипции необходимо участие специальных белковых факторов:
А,

Слайд 18На этом этапе действует ингибитор элонгации - стрептолидигин
3. Терминация
У E.coli . большинство
На этом этапе действует ингибитор элонгации - стрептолидигин
3. Терминация
У E.coli . большинство

Слайд 19У прокариот м-РНК синтезируется сразу в зрелом виде.
У эукариот все первичные транскрипты
У прокариот м-РНК синтезируется сразу в зрелом виде.
У эукариот все первичные транскрипты

Слайд 20
Благодаря антикодону -триплету в средней части молекулы тРНК, комплементарному определенному кодону на
Благодаря антикодону -триплету в средней части молекулы тРНК, комплементарному определенному кодону на
Слайд 21Компоненты белоксинтезирующей системы:
м-РНК - генетическая матрица - содержит линейную последовательность кодонов,
Компоненты белоксинтезирующей системы:
м-РНК - генетическая матрица - содержит линейную последовательность кодонов,
Слайд 221. ИНИЦИАЦИЯ ТРАНСЛЯЦИИ
На рибосоме имеются два участка: П-сайт или пептидильный сайт, служащий
1. ИНИЦИАЦИЯ ТРАНСЛЯЦИИ
На рибосоме имеются два участка: П-сайт или пептидильный сайт, служащий

Слайд 232. ЭЛОНГАЦИЯ ТРАНСЛЯЦИИ
Элонгация - это рост полипептидной цепи от N-конца
2. ЭЛОНГАЦИЯ ТРАНСЛЯЦИИ
Элонгация - это рост полипептидной цепи от N-конца

Слайд 24 Во время элонгации происходит важное событие - формирование пространственной структуры полипептидной цепи,
Во время элонгации происходит важное событие - формирование пространственной структуры полипептидной цепи,

Слайд 2533
Регуляция синтеза белка
Концентрация многих белков в клетке непостоянна и изменяется в
33
Регуляция синтеза белка
Концентрация многих белков в клетке непостоянна и изменяется в
 Змістовий модуль ІІ. Теорія вихованняТема 1.Суть і зміст процесу виховання. Процес виховання в національній школі
Змістовий модуль ІІ. Теорія вихованняТема 1.Суть і зміст процесу виховання. Процес виховання в національній школі Фитодизайн. Композиции
Фитодизайн. Композиции Лекция-3-русс
Лекция-3-русс Как сберечь свое здоровье?
Как сберечь свое здоровье? Объёмные открытки
Объёмные открытки Использование ЦОР в учебном процессе
Использование ЦОР в учебном процессе АНО «Молодежный театр «Light» Использование театральных методик в профилактике ВИЧ-инфекции
АНО «Молодежный театр «Light» Использование театральных методик в профилактике ВИЧ-инфекции (вычерчивание фигуры непрерывной линией)
(вычерчивание фигуры непрерывной линией) Союз как часть речи
Союз как часть речи Дети Беслана.
Дети Беслана. Юный разработчик. Занятие №9. Сборка. Динозавр
Юный разработчик. Занятие №9. Сборка. Динозавр Кондратий Фёдорович Рылеев
Кондратий Фёдорович Рылеев ЗВЕЗДООБРАЗОВАНИЕ
ЗВЕЗДООБРАЗОВАНИЕ Олимпийская символика
Олимпийская символика Государство и право
Государство и право Презентация на тему Арабы в средние века. Возникновение ислама
Презентация на тему Арабы в средние века. Возникновение ислама  Жаз айлары тақырыбындағы сабақ
Жаз айлары тақырыбындағы сабақ Употребление заглавной буквы в словах 2 класс
Употребление заглавной буквы в словах 2 класс Процессы управления проектами
Процессы управления проектами Мои первые словари
Мои первые словари Население Урала
Население Урала Вам дали это. Это и сейчас принадлежит вам. Вы его никогда никому не передавали, но им пользуются все ваши знакомые. Что это такое?
Вам дали это. Это и сейчас принадлежит вам. Вы его никогда никому не передавали, но им пользуются все ваши знакомые. Что это такое? Информация об областном конкурсе научных работ по проблемам развития агропромышленного комплекса.
Информация об областном конкурсе научных работ по проблемам развития агропромышленного комплекса. Внешняя политика России во второй половине 19 века.
Внешняя политика России во второй половине 19 века. Toxicology 1 (4)
Toxicology 1 (4) СОВЕТСКАЯ МУЗЫКА
СОВЕТСКАЯ МУЗЫКА Магниторецепция у человека: теория и практическое применение
Магниторецепция у человека: теория и практическое применение ПИЩЕВЫЕ ДОБАВКИ
ПИЩЕВЫЕ ДОБАВКИ