- Нуклеиновые кислоты

Содержание
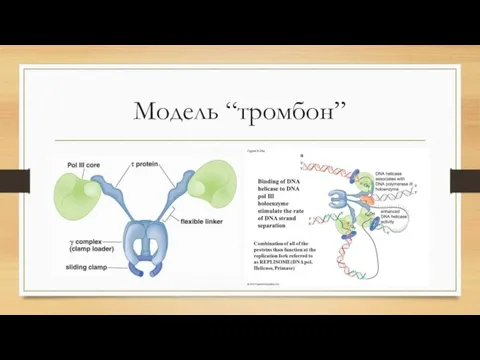
- 2. Модель “тромбон”
- 3. Топология ДНК Топология - раздел математики, имеющий своим назначением выяснение и исследование, в рамках математики, идеи
- 4. Математическое описание сверхспирализации ДНК В 1969 году Уайтом была предложена формула, которая связала порядок зацепления и
- 5. Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в результате которого ось двойной
- 6. Решение проблемы сверхспирализации ДНК Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию ДНК. Топоизомеразы способны релаксировать
- 7. Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы без затрат АТФ. Механизм
- 8. Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в виде димеров. Они осуществляют
- 9. Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины Топоизомеразы играют важную роль в процессах роста
- 10. Транскрипция РНК Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех
- 11. Свойства генетического кода Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон). Непрерывность
- 12. Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует
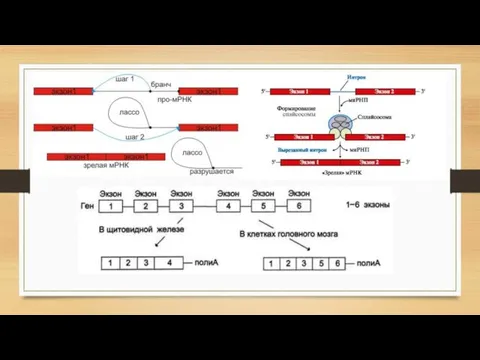
- 13. Процессинг РНК Кэпирование Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные связи в
- 14. 2) Полиаденилирование Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК (пре-мРНК). Иными словами,
- 16. 4) Редактирование Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется путём химической модификации
- 17. Общий вывод о созревании мРНК
- 18. Список использованной литературы 1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и др. 2) «Клетки»,
- 20. Скачать презентацию
Слайд 3Топология ДНК
Топология - раздел математики, имеющий своим назначением выяснение и исследование, в
Топология ДНК
Топология - раздел математики, имеющий своим назначением выяснение и исследование, в

Слайд 4Математическое описание сверхспирализации ДНК
В 1969 году Уайтом была предложена формула, которая связала
Математическое описание сверхспирализации ДНК
В 1969 году Уайтом была предложена формула, которая связала

Слайд 5Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в
Сверхспирализация ДНК — явление пере- или недоскручивания топологически замкнутых цепей ДНК, в

Слайд 6Решение проблемы сверхспирализации ДНК
Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию
Решение проблемы сверхспирализации ДНК
Топоизомеразы — класс ферментов-изомераз, которые влияют на топологию

Слайд 7Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы
Топоизомеразы I представляют собой мономерные белки. Они релаксируют ДНК, внося одноцепочечные разрывы

Слайд 8Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в
Топоизомеразы II функционируют у прокариот в виде тетрамеров, у эукариот — в

Слайд 9Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины
Топоизомеразы играют важную роль
Значение открытия топоизомераз в развитии фундаментальной и прикладной медицины
Топоизомеразы играют важную роль

Слайд 10Транскрипция РНК
Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы,
Транскрипция РНК
Транскрипция — процесс синтеза РНК с использованием ДНК в качестве матрицы,

Слайд 11Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет,
Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет,

Слайд 12Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA

Слайд 13Процессинг РНК
Кэпирование
Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные
Процессинг РНК
Кэпирование
Кэпирование защищает 5'-конец первичного транскрипта от действия рибонуклеаз, специфически разрезающих фосфодиэфирные

Слайд 142) Полиаденилирование
Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК
2) Полиаденилирование
Процесс присоединения большого количества остатков аденозинмонофосфата (поли(А)-хвоста) к 3'-концу первичной мРНК

Слайд 164) Редактирование
Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется
4) Редактирование
Молекулярно-биологический процесс, в ходе которого информация, содержащаяся в молекуле РНК изменяется

Слайд 17Общий вывод о созревании мРНК
Общий вывод о созревании мРНК

Слайд 18Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и
Список использованной литературы
1) «Общая биология», Л.В. Высоцкая, С.М. Глаголев, Г.М. Дымшиц и

 Презентация на тему Соседи восточных славян (6 класс)
Презентация на тему Соседи восточных славян (6 класс) Социальная политика занятости населения
Социальная политика занятости населения Лес – природное сообщество
Лес – природное сообщество Модель бизнеса
Модель бизнеса Сверление и зенкование
Сверление и зенкование ВКР: Разработка низкооборотистого асинхронного генератора электрического тока
ВКР: Разработка низкооборотистого асинхронного генератора электрического тока Конфликтные ситуации в семье
Конфликтные ситуации в семье Презентация на тему Житков "Как я ловил человечков" 4 класс
Презентация на тему Житков "Как я ловил человечков" 4 класс  Et Dieu crea la beaut
Et Dieu crea la beaut Суп. Ингредиенты
Суп. Ингредиенты ПЛАНИРУЙ ГЛАВНОЕ В СВОЕЙ ЖИЗНИ !!! (или о контрацепции для молодежи, позволяющей любить)
ПЛАНИРУЙ ГЛАВНОЕ В СВОЕЙ ЖИЗНИ !!! (или о контрацепции для молодежи, позволяющей любить) Презентация на тему Морские рыбы
Презентация на тему Морские рыбы  Экспертные системы
Экспертные системы Введение. Предмет и задачи курса. Сущность понятий менеджмент
Введение. Предмет и задачи курса. Сущность понятий менеджмент Образовательная программа ДОУ
Образовательная программа ДОУ 02_-_Suschnost_i_proiskhozhdenie_prava
02_-_Suschnost_i_proiskhozhdenie_prava Презентация на тему Морфологические признаки глагола
Презентация на тему Морфологические признаки глагола Биологические ритмы
Биологические ритмы Автоматическое формирование рубрикатора полнотекстовых документов
Автоматическое формирование рубрикатора полнотекстовых документов Гелий. Получение
Гелий. Получение Психологическая диагностика и коррекция аддиктивного поведения у подростков (из опыта работы психолога наркологического отделен
Психологическая диагностика и коррекция аддиктивного поведения у подростков (из опыта работы психолога наркологического отделен Химия в косметике
Химия в косметике Dr.Web Enterprise Suite
Dr.Web Enterprise Suite Управление рисками как система менеджмента
Управление рисками как система менеджмента Методические подходы к преподаванию курса обществознания по учебнику «Обществознание. Глобальный мир в XXI веке»
Методические подходы к преподаванию курса обществознания по учебнику «Обществознание. Глобальный мир в XXI веке» Презентация футболок для болельщиков футбольных сборных, участвующих на чемпионате мира 2018
Презентация футболок для болельщиков футбольных сборных, участвующих на чемпионате мира 2018 Абразивный инструмент
Абразивный инструмент Химическая промышленность
Химическая промышленность