- Процесс разложения органических азотистых соединений

Содержание
- 2. Нитрифицирующие бактерии получают энергию за счет окисления восстановленных форм азота (аммиак, азотистая кислота). Впервые определил хемолитоавтотрофную
- 3. Содержание доступного растениям азота в почве определяется не только процессами аммонификации, нитрификации, азотфиксации и вымыванием его
- 4. Способностью к фиксации молекулярного азота обладают исключительно прокариоты. Основная масса азота, содержащегося в живых организмах, своим
- 5. К группе симбиотических азотфиксаторов относятся бактерии родов Azorhizobium, Bradyrhizobium, Photorhizobium, Rhizobium и Sinorhizobium, образующие клубеньки на
- 6. Nod-факторы. Процесс симбиотической азотфиксации идет в клубеньках — специализированных органах растения-хозяина (рис. 6.5). КЛУБЕНЬКОВЫЕ БАКТЕРИИ Рис.
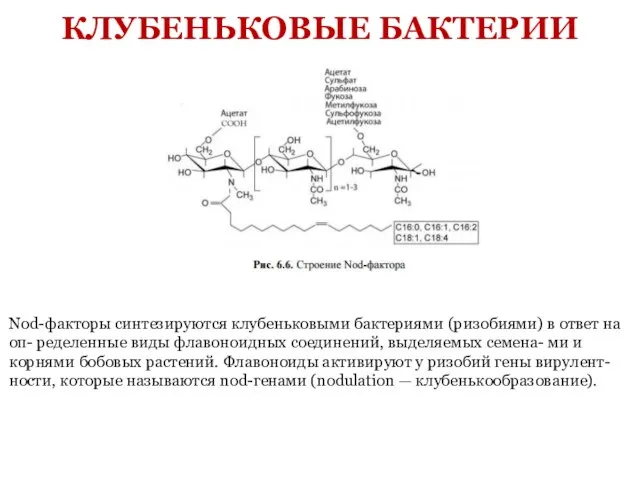
- 7. КЛУБЕНЬКОВЫЕ БАКТЕРИИ Nod-факторы синтезируются клубеньковыми бактериями (ризобиями) в ответ на оп- ределенные виды флавоноидных соединений, выделяемых
- 8. КЛУБЕНЬКОВЫЕ БАКТЕРИИ Найдено около 50 nod-генов, мутации в которых нарушают способность ризобий к формированию клубеньков. Большинство
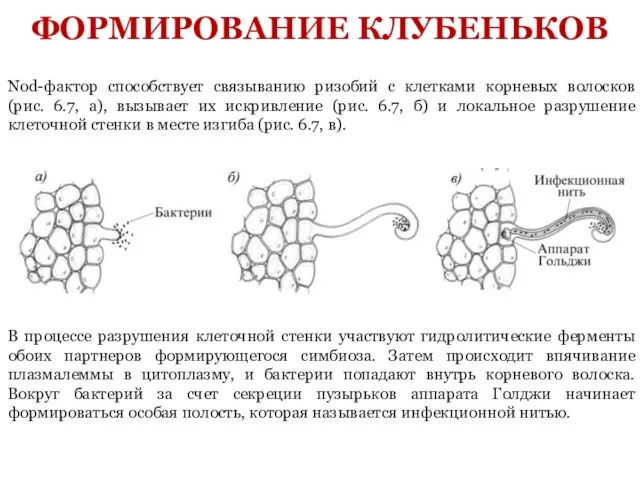
- 9. Nod-фактор способствует связыванию ризобий с клетками корневых волосков (рис. 6.7, а), вызывает их искривление (рис. 6.7,
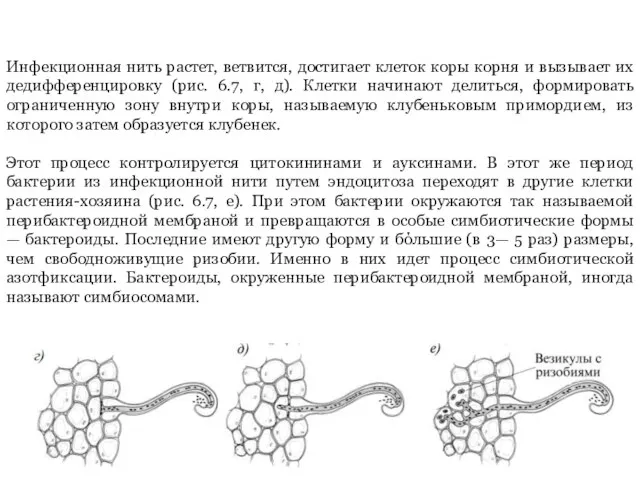
- 10. Инфекционная нить растет, ветвится, достигает клеток коры корня и вызывает их дедифференцировку (рис. 6.7, г, д).
- 11. По мере развития в клубеньке формируется собственная сосудистая система и очень плотный, "кожистый" слой клеток, который
- 12. Молекула азота N2 (N ≡ N) чрезвычайно прочна и химически инертна. Тем не менее большая группа
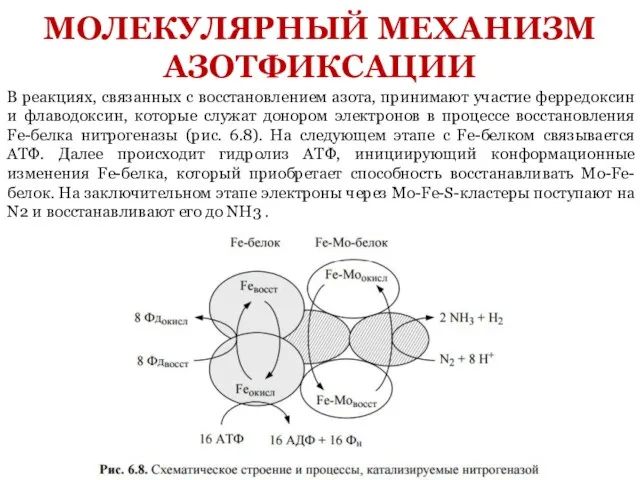
- 13. В реакциях, связанных с восстановлением азота, принимают участие ферредоксин и флаводоксин, которые служат донором электронов в
- 14. Поскольку нитрогеназа разрушается в присутствии кислорода, у азотфиксирующих микроорганизмов имеется ряд механизмов ее защиты. У ризобий
- 15. Расчеты показывают, что на связывание 1 г молекулярного азота растение затрачи- вает до 12 г углерода
- 16. Нитрат, попадая в растение, восстанавливается до аммония, который затем включается в аминокислоты. Процесс редукции нитрата в
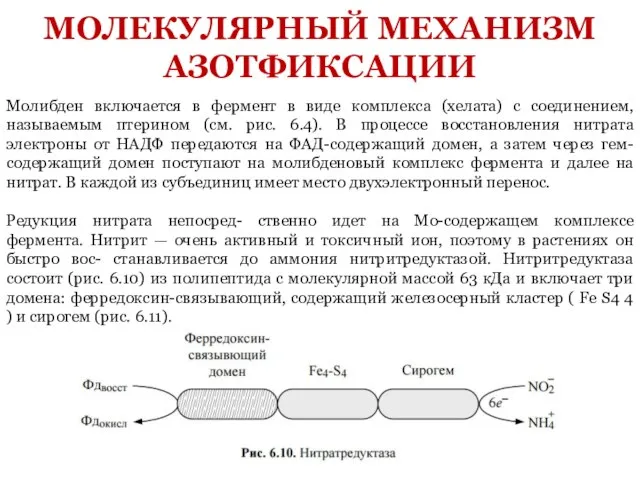
- 17. Молибден включается в фермент в виде комплекса (хелата) с соединением, называемым птерином (см. рис. 6.4). В
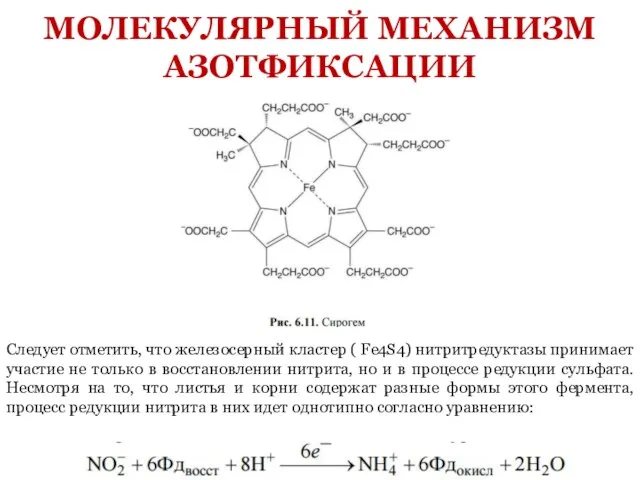
- 18. Следует отметить, что железосерный кластер ( Fe4S4) нитритредуктазы принимает участие не только в восстановлении нитрита, но
- 20. Скачать презентацию
Слайд 2Нитрифицирующие бактерии получают энергию за счет окисления восстановленных форм азота (аммиак, азотистая
Нитрифицирующие бактерии получают энергию за счет окисления восстановленных форм азота (аммиак, азотистая

Слайд 3Содержание доступного растениям азота в почве определяется не только процессами аммонификации, нитрификации,
Содержание доступного растениям азота в почве определяется не только процессами аммонификации, нитрификации,

Слайд 4Способностью к фиксации молекулярного азота обладают исключительно прокариоты. Основная масса азота, содержащегося
Способностью к фиксации молекулярного азота обладают исключительно прокариоты. Основная масса азота, содержащегося

Слайд 5К группе симбиотических азотфиксаторов относятся бактерии родов Azorhizobium, Bradyrhizobium, Photorhizobium, Rhizobium и
К группе симбиотических азотфиксаторов относятся бактерии родов Azorhizobium, Bradyrhizobium, Photorhizobium, Rhizobium и

Слайд 6Nod-факторы. Процесс симбиотической азотфиксации идет в клубеньках — специализированных органах растения-хозяина (рис.
Nod-факторы. Процесс симбиотической азотфиксации идет в клубеньках — специализированных органах растения-хозяина (рис.

Слайд 7КЛУБЕНЬКОВЫЕ БАКТЕРИИ
Nod-факторы синтезируются клубеньковыми бактериями (ризобиями) в ответ на оп- ределенные виды
КЛУБЕНЬКОВЫЕ БАКТЕРИИ
Nod-факторы синтезируются клубеньковыми бактериями (ризобиями) в ответ на оп- ределенные виды
Слайд 8КЛУБЕНЬКОВЫЕ БАКТЕРИИ
Найдено около 50 nod-генов, мутации в которых нарушают способность ризобий к
КЛУБЕНЬКОВЫЕ БАКТЕРИИ
Найдено около 50 nod-генов, мутации в которых нарушают способность ризобий к

Слайд 9Nod-фактор способствует связыванию ризобий с клетками корневых волосков (рис. 6.7, а), вызывает
Nod-фактор способствует связыванию ризобий с клетками корневых волосков (рис. 6.7, а), вызывает
Слайд 10Инфекционная нить растет, ветвится, достигает клеток коры корня и вызывает их дедифференцировку
Инфекционная нить растет, ветвится, достигает клеток коры корня и вызывает их дедифференцировку
Слайд 11По мере развития в клубеньке формируется собственная сосудистая система и очень плотный,
По мере развития в клубеньке формируется собственная сосудистая система и очень плотный,

Слайд 12Молекула азота N2 (N ≡ N) чрезвычайно прочна и химически инертна. Тем
Молекула азота N2 (N ≡ N) чрезвычайно прочна и химически инертна. Тем

Слайд 13В реакциях, связанных с восстановлением азота, принимают участие ферредоксин и флаводоксин, которые
В реакциях, связанных с восстановлением азота, принимают участие ферредоксин и флаводоксин, которые
Слайд 14Поскольку нитрогеназа разрушается в присутствии кислорода, у азотфиксирующих микроорганизмов имеется ряд механизмов
Поскольку нитрогеназа разрушается в присутствии кислорода, у азотфиксирующих микроорганизмов имеется ряд механизмов

Слайд 15Расчеты показывают, что на связывание 1 г молекулярного азота растение затрачи- вает
Расчеты показывают, что на связывание 1 г молекулярного азота растение затрачи- вает

Слайд 16Нитрат, попадая в растение, восстанавливается до аммония, который затем включается в аминокислоты.
Нитрат, попадая в растение, восстанавливается до аммония, который затем включается в аминокислоты.

Слайд 17Молибден включается в фермент в виде комплекса (хелата) с соединением, называемым птерином
Молибден включается в фермент в виде комплекса (хелата) с соединением, называемым птерином
Слайд 18Следует отметить, что железосерный кластер ( Fe4S4) нитритредуктазы принимает участие не только
Следует отметить, что железосерный кластер ( Fe4S4) нитритредуктазы принимает участие не только
 Психология опознания
Психология опознания Мишка косолапый
Мишка косолапый Презентация на тему Подвигу народа жить в веках!
Презентация на тему Подвигу народа жить в веках! ПРАВИЛА УСПЕШНОГО РЕЗЮМЕ
ПРАВИЛА УСПЕШНОГО РЕЗЮМЕ Кальяны MYA
Кальяны MYA Звукоцвет
Звукоцвет Еженедельник «СЛОБОДА» ИНТЕРНЕТ-ПОРТАЛ MySLO.ru. - презентация
Еженедельник «СЛОБОДА» ИНТЕРНЕТ-ПОРТАЛ MySLO.ru. - презентация Разрешение конфликтных ситуаций в коллективе
Разрешение конфликтных ситуаций в коллективе Сотрудничество фирмы «1С» с учебными заведениями
Сотрудничество фирмы «1С» с учебными заведениями Страховая медицинская организация, осуществляющая деятельность в сфере ОМС
Страховая медицинская организация, осуществляющая деятельность в сфере ОМС Понятие философии. Особенности философского знания
Понятие философии. Особенности философского знания Покупка голосов, а также индифферентная позиция электората в избирательном праве
Покупка голосов, а также индифферентная позиция электората в избирательном праве Презентация на тему Навруз Байрам
Презентация на тему Навруз Байрам Kryvyi Rih Area
Kryvyi Rih Area Физика вокруг нас 5 класс
Физика вокруг нас 5 класс «Педагогическое взаимодействие как одно из важнейших условий компетентностного подхода в обучении»
«Педагогическое взаимодействие как одно из важнейших условий компетентностного подхода в обучении» Презентация на тему Древние германцы и Римская империя
Презентация на тему Древние германцы и Римская империя  философия древней индии
философия древней индии Волшебные голоса
Волшебные голоса ПРИЧИНЫ И ПРОФИЛАКТИКА ЭМОЦИОНАЛЬНЫХ СРЫВОВ «Наша жизнь – это наше здоровье Наше здоровье – это наше здоровье».
ПРИЧИНЫ И ПРОФИЛАКТИКА ЭМОЦИОНАЛЬНЫХ СРЫВОВ «Наша жизнь – это наше здоровье Наше здоровье – это наше здоровье». Массаж
Массаж Субкультуры
Субкультуры Компетентный родитель–счастливый ребенок
Компетентный родитель–счастливый ребенок Зачет по теме "Синтаксис и пунктуация"
Зачет по теме "Синтаксис и пунктуация" В каком банке выгоднее взять деньги?
В каком банке выгоднее взять деньги? Основы нефтедобычи
Основы нефтедобычи Предварительная обработка результатов функциональных испытаний гидромуфты «Фойт» на КС Чебоксарская 30.08.05г.
Предварительная обработка результатов функциональных испытаний гидромуфты «Фойт» на КС Чебоксарская 30.08.05г. МАШИНОВЕДЕНИЕ
МАШИНОВЕДЕНИЕ