- 832648_№3-realizaciya-nasledstvennoi-informacii

Содержание
- 2. ПЛАН ЛЕКЦИИ: Клеточный цикл: характеристика фаз, биологический смысл. Разновидности митоза. Мейоз, характеристика 1-го и 2-го делений.
- 3. СХЕМА МЕЙОЗА. ОСНОВНЫЕ ФАЗЫ. МЕЙОЗ – способ деления эукариотической клетки, при котором происходит редукция (уменьшение) числа
- 4. СХЕМА МЕЙОЗА МЕЙОЗ состоит из: ИНТЕРФАЗА МЕЙОЗ 1 МЕЙОЗ 2 Пресинтетический период профаза 1 профаза 2
- 7. ПРОФАЗА 1. ЛЕПТОТЕНА и ЗИГОТЕНА Хромосомы спирализуются, утолщаются и укорачиваются. Начало конъюгации гомологичных хромосом, которые объединяются
- 8. ПРОФАЗА 1. ПАХИТЕНА. Спирализация продолжается. КРОССИНГОВЕР. 1п бив 4хр4с.
- 9. ПРОФАЗА 1 МЕЙОЗА. КРОССИНГОВЕР.
- 10. ПРОФАЗА 1. ДИПЛОТЕНА. Возникновение сил отталкивания между гомологами, которые начинают отделяться сначала в области центромер. Образование
- 11. ПРОФАЗА 1. ДИАКИНЕЗ. Хромосомы удерживаются в отдельных точках. Окончательного разрушения бивалентов не происходит. 1п бив 4хр4с.
- 12. ПРОФАЗА 1. ДИКТИОТЕНА. Только в овогенезе. Хромосомы принимают форму «ламповых щеток» и прекращают структурные изменения до
- 13. М Е Й О З. П Р О Ф А З А 1 .
- 14. МЕТАФАЗА 1. БИВАЛЕНТЫ В ЭКВАТОРИАЛЬНОЙ ПЛОСКОСТИ. 2n4C
- 15. АНАФАЗА 1. 1. ГОМОЛОГИ РАСХОДЯТСЯ ПО ПОЛЮСАМ. 2. РАСХОЖДЕНИЕ ХРОМОСОМ НОСИТ НЕЗАВИСИМЫЙ ХАРАКТЕР. 3. (1п2хр2с) х
- 17. МЕЙОЗ 2. Эквационное деление. По типу митоза. Профаза 2 короткая. Метафаза 2 – на экваторе двухроматидные
- 18. Биологическое значение мейоза Рекомбинация генетического материала (кроссинговер в профазу 1 и независимое расхождение гомологичных хромосом по
- 19. Молекулярная биология Экспрессия гена — процесс реализации информации, закодированной в гене. Состоит из двух основных стадий
- 20. 3. ОСНОВНЫЕ ЭТАПЫ БИОСИНТЕЗА БЕЛКА. РОЛЬ РНК. СХЕМА БИОСИНТЕЗА: ТРАНСКРИПЦИЯ (ЯДРО). СОЗРЕВАНИЕ м-РНК. ТРАНСЛЯЦИЯ (РИБОСОМА).
- 21. Центральная догма молекулярной биологии ДНК → РНК → белок
- 22. Транскрипционный аппарат клетки Транскрипция — синтез РНК на матрице ДНК. Транскрипт — продукт транскрипции, т. е.
- 23. Транскрипционный аппарат клетки Этапы транскрипции: Присоединение РНК-полимеразы Инициация Элонгация Терминация
- 24. Транскрипционный аппарат клетки Промотор — регуляторный участок гена, к которому присоединяется РНК-полимераза с тем, чтобы начать
- 25. Транскрипционный аппарат клетки Элонгация – удлинение цепи РНК за счет комплементарного присоединения новых нуклеотидов
- 26. Транскрипционный аппарат клетки Терминатор – это участок, где прекращается дальнейший рост цепи РНК и происходит ее
- 27. Схема транскрипции
- 28. ТРАНСКРИПЦИЯ - процесс переписывания информации с ДНК на и-РНК (м-РНК). Синтез начинается с обнаружения РНК-полимеразой в
- 29. ТРАНСКРИПЦИЯ 4. Синтез и-РНК по принципу комплементарности: 5. Синтез продолжается до терминатора тринскрипции. 6. Фрагмент ДНК
- 30. Транскрипционный аппарат клетки Процессинг – совокупность событий, связанных с претрансляционным преобразованием первичного РНК-транскрипта
- 31. Транскрипционный аппарат клетки К 5′-концу РНК добавляется кэп (метилированный гуаниновый нуклеотид), защищающий транскрипт от деградации.
- 32. Транскрипционный аппарат клетки К 3′-концу РНК присоединяется «поли-А-хвост» - последовательность из 100-200 остатков адениловой кислоты, которая
- 33. Транскрипционный аппарат клетки Экзон — значащий участок гена, на котором записана информация о порядке аминокислот в
- 34. Транскрипционный аппарат клетки Интрон — некодирующий участок гена, который переписывается на иРНК, а затем удаляется из
- 35. Транскрипционный аппарат клетки Сплайсинг — процесс формирования зрелой и-РНК путем удаления внутренних частей молекулы — интронов.
- 36. Схема сплайсинга
- 37. Схема сплайсинга
- 38. Общая схема транскрипции, процессинга gРНК и трансляции
- 39. Альтернативный сплайсинг дает возможность синтеза различных молекул белка на базе одной нуклеотидной последовательности
- 40. Транскрипционный аппарат клетки Неактивное состояние гена может быть обусловлено компактизацией хроматина. Иногда компактизацию хроматина объясняют метилированием
- 41. Молекулярно-генетический уровень организации жизни Трансляционный аппарат клетки
- 42. Трансляционный аппарат клетки Трансляция — процесс биосинтеза белка, определяемый матричной РНК.
- 43. Трансляционный аппарат клетки В 1968 г. За открытие генетического кода Р.Хорана, Р.Холли и М.Ниренберг получили Нобелевскую
- 44. Трансляционный аппарат клетки Генетический код – это способ записи информации об аминокислотном составе белка с помощью
- 45. ТРАНСЛЯЦИЯ. т-РНК. Синтезируется на матрице ДНК. Число нуклеотидов – 75-95. Лист клевера, в котором выделяют 4
- 46. ТРАНСЛЯЦИЯ. т-РНК. 5. ИНОЗИН в составе антикодона способен комплементарно соединяться с У, Ц и А м-РНК
- 47. СОЕДИНЕНИЕ т-РНК С АМИНОКИСЛОТОЙ А\к-та взаимодействует с АТФ, активация ее и образование аденилированной аминокислоты. Аденилированная аминокислота
- 48. Строение рибосом
- 49. Трансляционный аппарат клетки В рибосоме имеются три различных участка, с которыми связывается РНК: один для мРНК
- 50. Трансляционный аппарат клетки Участки для т-РНК называются Р -(пептидильный) и А- (акцепторный или аминоацильный) участки
- 51. Трансляционный аппарат клетки В фазе инициации субъединицы рибосомы объединяются с мРНК и в систему поступает первая
- 52. ИНИЦИАЦИЯ БЕЛКОВОГО СИНТЕЗА
- 53. Рибосомы
- 54. Трансляционный аппарат клетки Элонгация (удлинение) – циклически повторяющиеся события, связанные с включением аминокислот в белковую цепочку.
- 55. ФАЗА ЭЛОНГАЦИИ При соединении антикодона т-РНК с кодоном м-РНК транспортируемая аминокислота располагается на А-участке. Шаг рибосомы
- 56. Элонгация
- 57. Трансляционный аппарат клетки Терминация (окончание биосинтеза) связана с поступлением в рибосому одного из нонсенс-кодонов: УАА, УАГ
- 58. Полирибосома (полисома)
- 59. ФАЗЫ ТРАНСЛЯЦИОННОГО СИНТЕЗА: инициации, элонгации, терминации Фаза инициации: Рибосомные РНК обеспечивают взаимосвязь с м-РНК (рамка считывания).
- 60. Белки в эволюции и онтогенезе Бактериальные и-РНК полицистронны, т.е.кодируют несколько белков по одной и-РНК, а эукариотические
- 61. Трансляционный аппарат клетки Действие многих эффективных антибиотиков основано на подавлении биосинтеза белков.
- 62. Трансляционный аппарат клетки Посттрансляционная модификация заключается в укладке первичной структуры белка в структуры высшего порядка.
- 63. 4. Особенности организации и экспрессии у эукариот и прокариот ЕДИНСТВО ПРОИСХОЖДЕНИЯ: Генетический материал у прокариот и
- 64. ОТЛИЧИЯ В ЭКСПРЕССИИ ГЕНОВ У ПРО- и ЭУКАРИОТ
- 65. РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ У ПРОКАРИОТ МОДЕЛЬ ОПЕРОНА. 1. Начало 60-х годов 20-го века: Ф. ЖАКОБ, А.
- 66. ИНДУКЦИЯ 4. Процесс индукции включает следующие фазы: А) ПРОИЗВОДСТВО МОЛЕКУЛЫ-РЕПРЕССОРА: ген-регулятор синтезирует репрессор; при отсутствии субстрата
- 67. ИНДУКЦИЯ В) Транскрипция генов и производство ферментов: РНК-полимераза получает доступ к оператору: синтез фермента.
- 68. ИНДУКЦИЯ. РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ.
- 70. Скачать презентацию
Слайд 2ПЛАН ЛЕКЦИИ:
Клеточный цикл: характеристика фаз, биологический смысл. Разновидности митоза.
Мейоз, характеристика 1-го и
ПЛАН ЛЕКЦИИ:
Клеточный цикл: характеристика фаз, биологический смысл. Разновидности митоза.
Мейоз, характеристика 1-го и

Слайд 3СХЕМА МЕЙОЗА. ОСНОВНЫЕ ФАЗЫ.
МЕЙОЗ – способ деления эукариотической клетки, при котором происходит
СХЕМА МЕЙОЗА. ОСНОВНЫЕ ФАЗЫ.
МЕЙОЗ – способ деления эукариотической клетки, при котором происходит

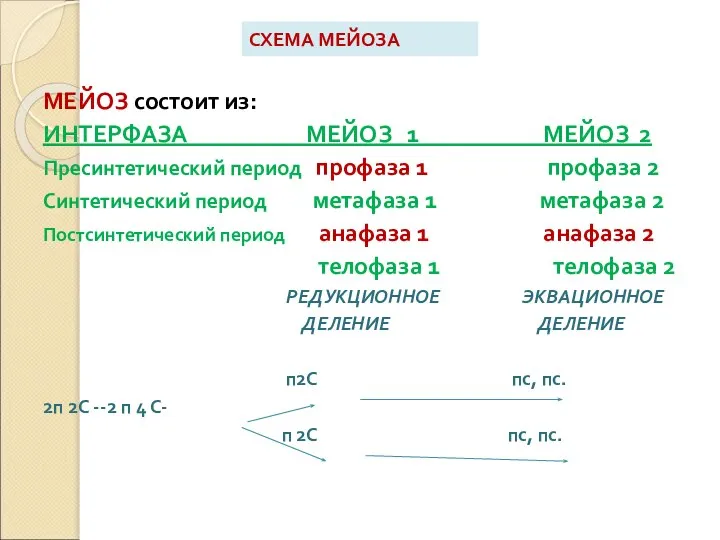
Слайд 4СХЕМА МЕЙОЗА
МЕЙОЗ состоит из:
ИНТЕРФАЗА МЕЙОЗ 1 МЕЙОЗ 2
Пресинтетический период профаза 1 профаза
СХЕМА МЕЙОЗА
МЕЙОЗ состоит из:
ИНТЕРФАЗА МЕЙОЗ 1 МЕЙОЗ 2
Пресинтетический период профаза 1 профаза
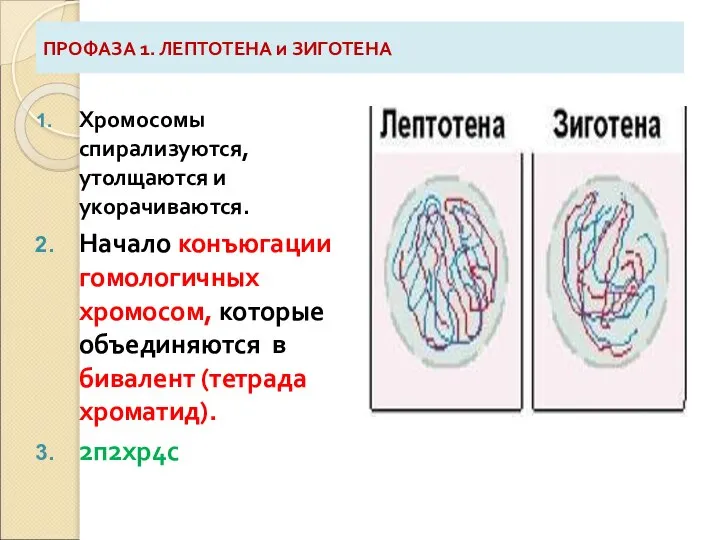
Слайд 7ПРОФАЗА 1. ЛЕПТОТЕНА и ЗИГОТЕНА
Хромосомы спирализуются, утолщаются и укорачиваются.
Начало конъюгации гомологичных хромосом,
ПРОФАЗА 1. ЛЕПТОТЕНА и ЗИГОТЕНА
Хромосомы спирализуются, утолщаются и укорачиваются.
Начало конъюгации гомологичных хромосом,
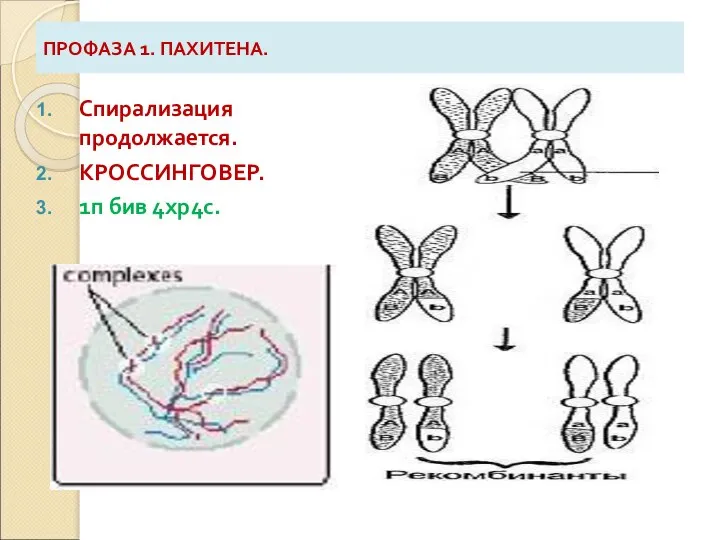
Слайд 8ПРОФАЗА 1. ПАХИТЕНА.
Спирализация продолжается.
КРОССИНГОВЕР.
1п бив 4хр4с.
ПРОФАЗА 1. ПАХИТЕНА.
Спирализация продолжается.
КРОССИНГОВЕР.
1п бив 4хр4с.
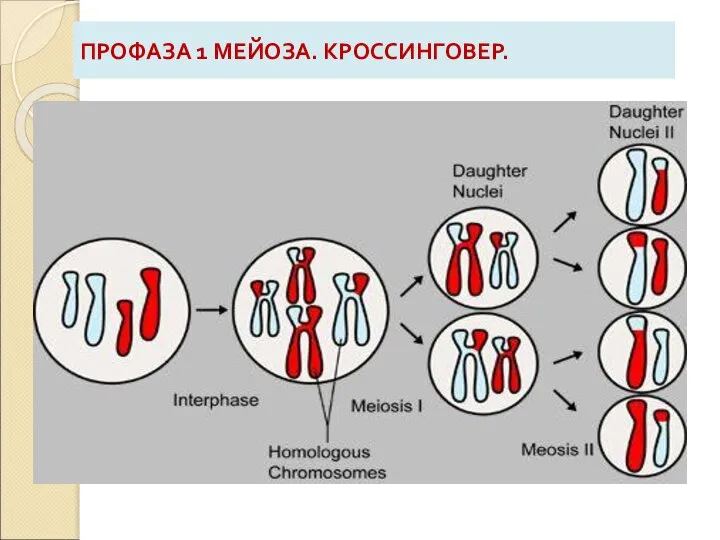
Слайд 9ПРОФАЗА 1 МЕЙОЗА. КРОССИНГОВЕР.
ПРОФАЗА 1 МЕЙОЗА. КРОССИНГОВЕР.
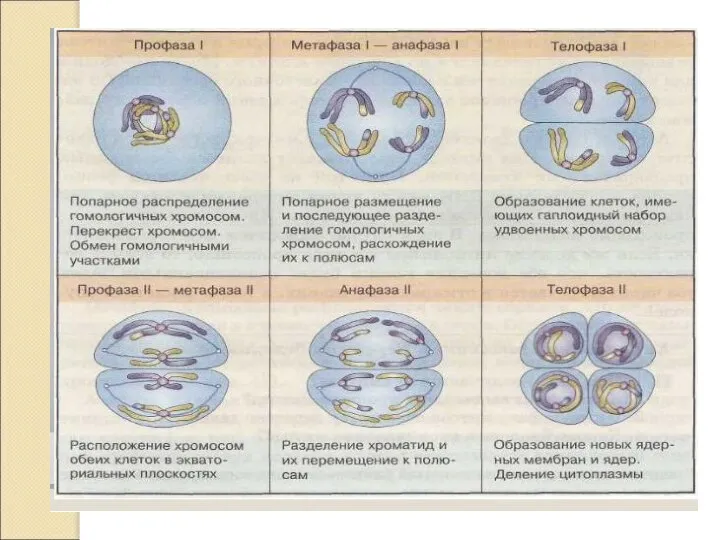
Слайд 10ПРОФАЗА 1. ДИПЛОТЕНА.
Возникновение сил отталкивания между гомологами, которые начинают отделяться сначала в
ПРОФАЗА 1. ДИПЛОТЕНА.
Возникновение сил отталкивания между гомологами, которые начинают отделяться сначала в

Слайд 11ПРОФАЗА 1. ДИАКИНЕЗ.
Хромосомы удерживаются в отдельных точках.
Окончательного разрушения бивалентов не происходит.
1п бив
ПРОФАЗА 1. ДИАКИНЕЗ.
Хромосомы удерживаются в отдельных точках.
Окончательного разрушения бивалентов не происходит.
1п бив

Слайд 12ПРОФАЗА 1. ДИКТИОТЕНА.
Только в овогенезе.
Хромосомы принимают форму «ламповых щеток» и прекращают структурные
ПРОФАЗА 1. ДИКТИОТЕНА.
Только в овогенезе.
Хромосомы принимают форму «ламповых щеток» и прекращают структурные

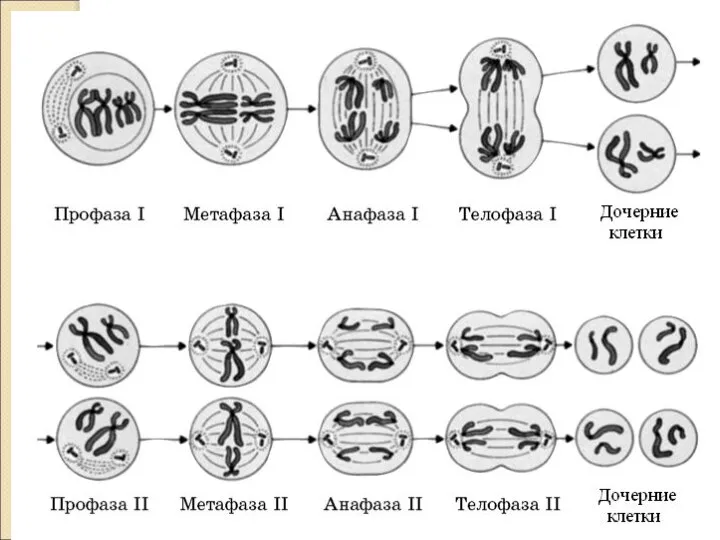
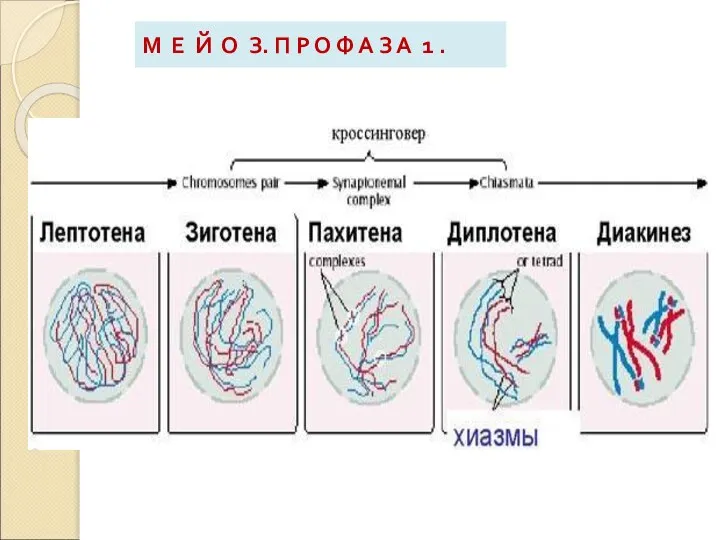
Слайд 13М Е Й О З. П Р О Ф А З А
М Е Й О З. П Р О Ф А З А
Слайд 14МЕТАФАЗА 1.
БИВАЛЕНТЫ В ЭКВАТОРИАЛЬНОЙ ПЛОСКОСТИ.
2n4C
МЕТАФАЗА 1.
БИВАЛЕНТЫ В ЭКВАТОРИАЛЬНОЙ ПЛОСКОСТИ.
2n4C

Слайд 15АНАФАЗА 1.
1. ГОМОЛОГИ РАСХОДЯТСЯ ПО ПОЛЮСАМ.
2. РАСХОЖДЕНИЕ ХРОМОСОМ НОСИТ НЕЗАВИСИМЫЙ ХАРАКТЕР.
3. (1п2хр2с)
АНАФАЗА 1.
1. ГОМОЛОГИ РАСХОДЯТСЯ ПО ПОЛЮСАМ.
2. РАСХОЖДЕНИЕ ХРОМОСОМ НОСИТ НЕЗАВИСИМЫЙ ХАРАКТЕР.
3. (1п2хр2с)

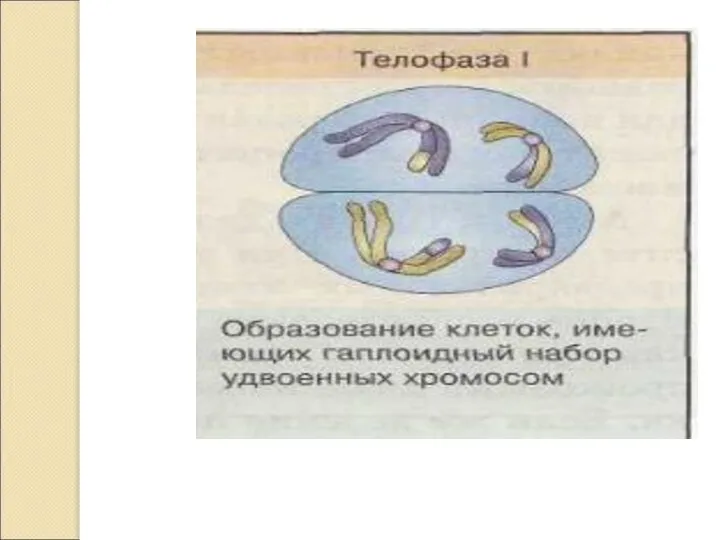
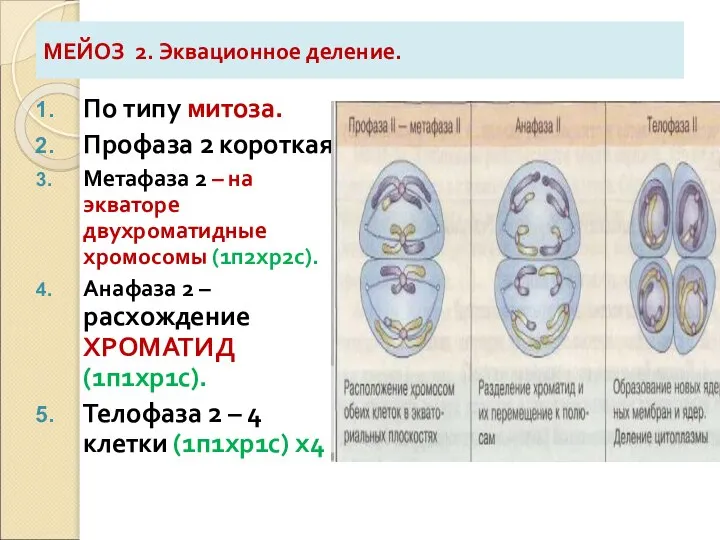
Слайд 17МЕЙОЗ 2. Эквационное деление.
По типу митоза.
Профаза 2 короткая.
Метафаза 2 – на экваторе
МЕЙОЗ 2. Эквационное деление.
По типу митоза.
Профаза 2 короткая.
Метафаза 2 – на экваторе
Слайд 18Биологическое значение мейоза
Рекомбинация генетического материала (кроссинговер в профазу 1 и независимое расхождение
Биологическое значение мейоза
Рекомбинация генетического материала (кроссинговер в профазу 1 и независимое расхождение

Слайд 19
Молекулярная биология
Экспрессия гена — процесс реализации информации, закодированной в гене. Состоит
Молекулярная биология
Экспрессия гена — процесс реализации информации, закодированной в гене. Состоит

Слайд 203. ОСНОВНЫЕ ЭТАПЫ БИОСИНТЕЗА БЕЛКА. РОЛЬ РНК.
СХЕМА БИОСИНТЕЗА:
ТРАНСКРИПЦИЯ (ЯДРО).
СОЗРЕВАНИЕ м-РНК.
ТРАНСЛЯЦИЯ (РИБОСОМА).
3. ОСНОВНЫЕ ЭТАПЫ БИОСИНТЕЗА БЕЛКА. РОЛЬ РНК.
СХЕМА БИОСИНТЕЗА:
ТРАНСКРИПЦИЯ (ЯДРО).
СОЗРЕВАНИЕ м-РНК.
ТРАНСЛЯЦИЯ (РИБОСОМА).

Слайд 21Центральная догма молекулярной биологии
ДНК → РНК → белок
Центральная догма молекулярной биологии
ДНК → РНК → белок

Слайд 22Транскрипционный
аппарат клетки
Транскрипция — синтез РНК на матрице ДНК.
Транскрипт — продукт транскрипции,
Транскрипционный
аппарат клетки
Транскрипция — синтез РНК на матрице ДНК. Транскрипт — продукт транскрипции,

Слайд 23Транскрипционный
аппарат клетки
Этапы транскрипции:
Присоединение РНК-полимеразы
Инициация
Элонгация
Терминация
Транскрипционный
аппарат клетки
Этапы транскрипции:
Присоединение РНК-полимеразы
Инициация
Элонгация
Терминация

Слайд 24Транскрипционный
аппарат клетки
Промотор — регуляторный участок гена, к которому присоединяется РНК-полимераза с
Транскрипционный
аппарат клетки
Промотор — регуляторный участок гена, к которому присоединяется РНК-полимераза с

Слайд 25Транскрипционный
аппарат клетки
Элонгация – удлинение цепи РНК за счет комплементарного присоединения новых
Транскрипционный
аппарат клетки
Элонгация – удлинение цепи РНК за счет комплементарного присоединения новых

Слайд 26Транскрипционный
аппарат клетки
Терминатор – это участок, где прекращается дальнейший рост цепи РНК
Транскрипционный
аппарат клетки
Терминатор – это участок, где прекращается дальнейший рост цепи РНК

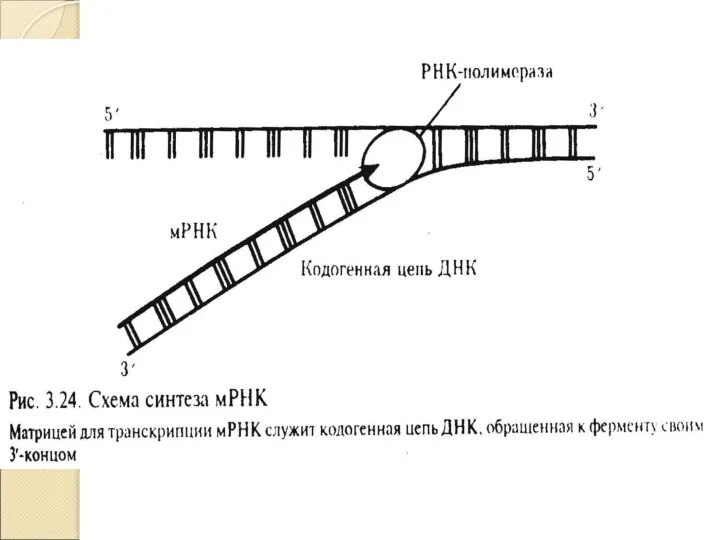
Слайд 27Схема транскрипции
Схема транскрипции
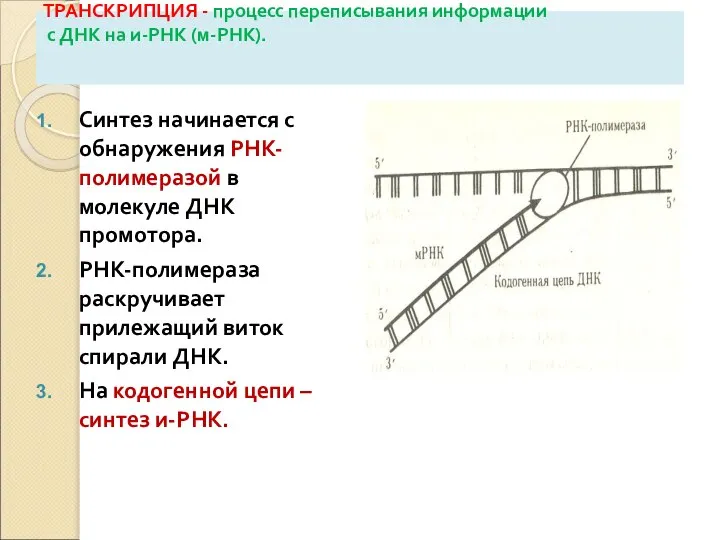
Слайд 28ТРАНСКРИПЦИЯ - процесс переписывания информации
с ДНК на и-РНК (м-РНК).
Синтез начинается с
ТРАНСКРИПЦИЯ - процесс переписывания информации
с ДНК на и-РНК (м-РНК).
Синтез начинается с
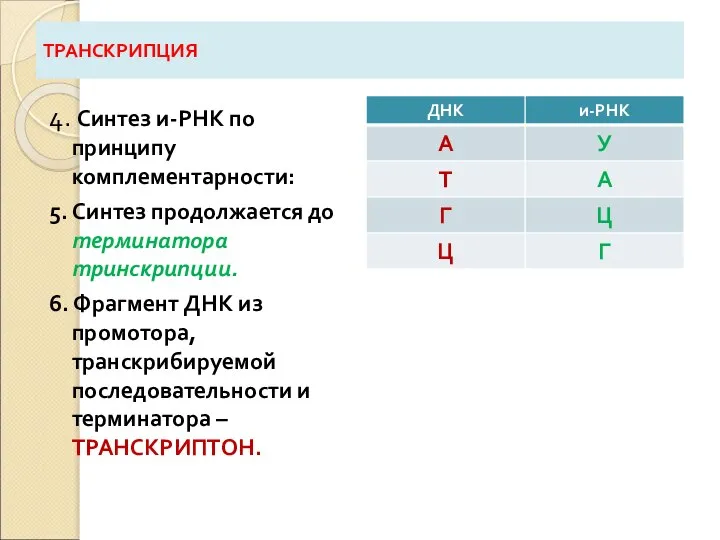
Слайд 29ТРАНСКРИПЦИЯ
4. Синтез и-РНК по принципу комплементарности:
5. Синтез продолжается до терминатора тринскрипции.
6. Фрагмент
ТРАНСКРИПЦИЯ
4. Синтез и-РНК по принципу комплементарности:
5. Синтез продолжается до терминатора тринскрипции.
6. Фрагмент
Слайд 30Транскрипционный
аппарат клетки
Процессинг – совокупность событий, связанных с претрансляционным преобразованием первичного РНК-транскрипта
Транскрипционный
аппарат клетки
Процессинг – совокупность событий, связанных с претрансляционным преобразованием первичного РНК-транскрипта

Слайд 31Транскрипционный
аппарат клетки
К 5′-концу РНК добавляется кэп (метилированный гуаниновый нуклеотид), защищающий транскрипт
Транскрипционный
аппарат клетки
К 5′-концу РНК добавляется кэп (метилированный гуаниновый нуклеотид), защищающий транскрипт

Слайд 32Транскрипционный
аппарат клетки
К 3′-концу РНК присоединяется «поли-А-хвост» - последовательность из 100-200 остатков
Транскрипционный
аппарат клетки
К 3′-концу РНК присоединяется «поли-А-хвост» - последовательность из 100-200 остатков

Слайд 33Транскрипционный
аппарат клетки
Экзон — значащий участок гена, на котором записана информация о
Транскрипционный
аппарат клетки
Экзон — значащий участок гена, на котором записана информация о

Слайд 34Транскрипционный
аппарат клетки
Интрон — некодирующий участок гена, который переписывается на иРНК, а
Транскрипционный
аппарат клетки
Интрон — некодирующий участок гена, который переписывается на иРНК, а

Слайд 35Транскрипционный
аппарат клетки
Сплайсинг — процесс формирования зрелой и-РНК путем удаления внутренних частей
Транскрипционный
аппарат клетки
Сплайсинг — процесс формирования зрелой и-РНК путем удаления внутренних частей

Слайд 36Схема сплайсинга
Схема сплайсинга
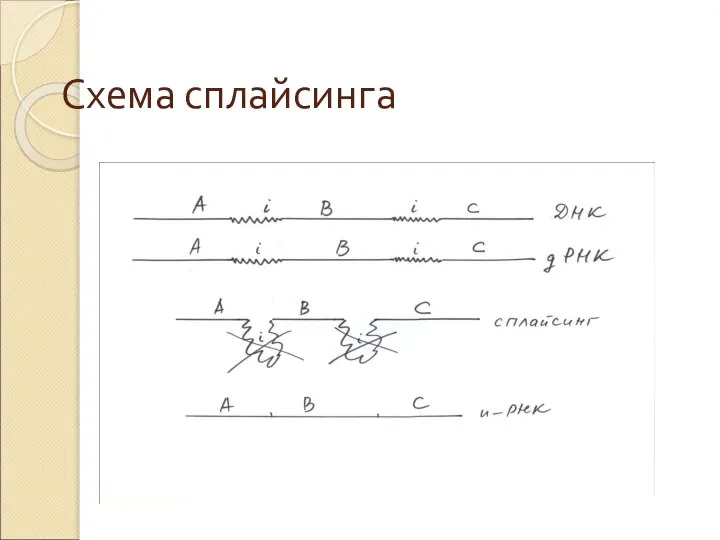
Слайд 37Схема сплайсинга
Схема сплайсинга
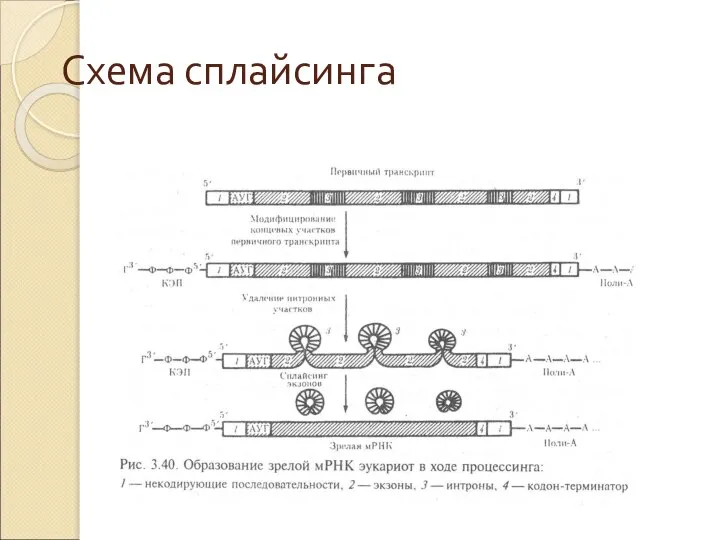
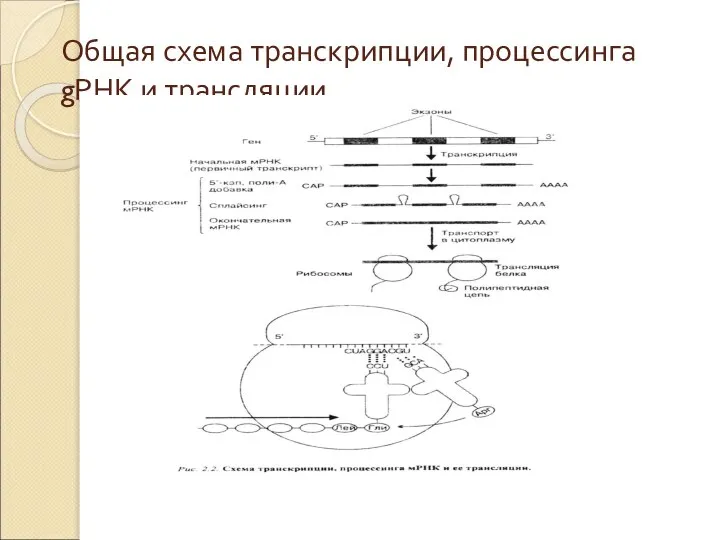
Слайд 38Общая схема транскрипции, процессинга gРНК и трансляции
Общая схема транскрипции, процессинга gРНК и трансляции
Слайд 39Альтернативный сплайсинг дает возможность синтеза различных молекул белка на базе одной нуклеотидной
Альтернативный сплайсинг дает возможность синтеза различных молекул белка на базе одной нуклеотидной
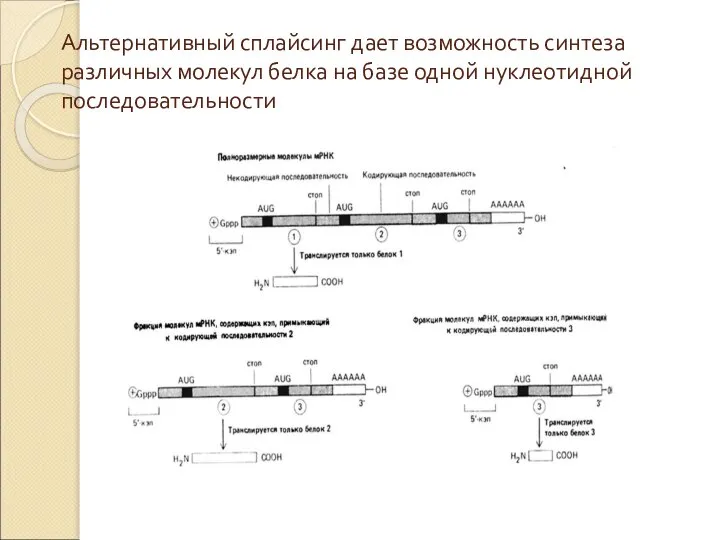
Слайд 40Транскрипционный
аппарат клетки
Неактивное состояние гена может быть обусловлено компактизацией хроматина. Иногда компактизацию
Транскрипционный
аппарат клетки
Неактивное состояние гена может быть обусловлено компактизацией хроматина. Иногда компактизацию

Слайд 41Молекулярно-генетический уровень организации жизни
Трансляционный аппарат клетки
Молекулярно-генетический уровень организации жизни
Трансляционный аппарат клетки

Слайд 42Трансляционный аппарат клетки
Трансляция — процесс биосинтеза белка, определяемый матричной РНК.
Трансляционный аппарат клетки
Трансляция — процесс биосинтеза белка, определяемый матричной РНК.

Слайд 43Трансляционный аппарат клетки
В 1968 г. За открытие генетического кода Р.Хорана, Р.Холли и
Трансляционный аппарат клетки
В 1968 г. За открытие генетического кода Р.Хорана, Р.Холли и

Слайд 44Трансляционный аппарат клетки
Генетический код – это способ записи информации об аминокислотном составе
Трансляционный аппарат клетки
Генетический код – это способ записи информации об аминокислотном составе

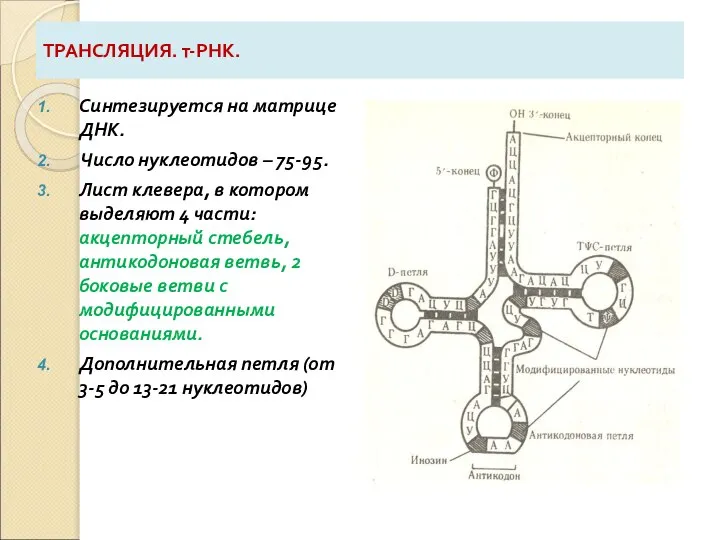
Слайд 45ТРАНСЛЯЦИЯ. т-РНК.
Синтезируется на матрице ДНК.
Число нуклеотидов – 75-95.
Лист клевера, в котором
ТРАНСЛЯЦИЯ. т-РНК.
Синтезируется на матрице ДНК.
Число нуклеотидов – 75-95.
Лист клевера, в котором
Слайд 46ТРАНСЛЯЦИЯ. т-РНК.
5. ИНОЗИН в составе антикодона способен комплементарно соединяться с У, Ц
ТРАНСЛЯЦИЯ. т-РНК.
5. ИНОЗИН в составе антикодона способен комплементарно соединяться с У, Ц

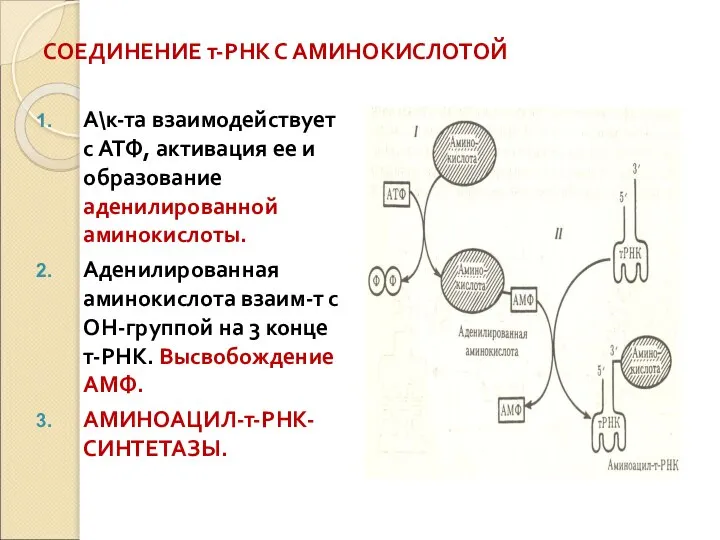
Слайд 47СОЕДИНЕНИЕ т-РНК С АМИНОКИСЛОТОЙ
А\к-та взаимодействует с АТФ, активация ее и образование аденилированной
СОЕДИНЕНИЕ т-РНК С АМИНОКИСЛОТОЙ
А\к-та взаимодействует с АТФ, активация ее и образование аденилированной
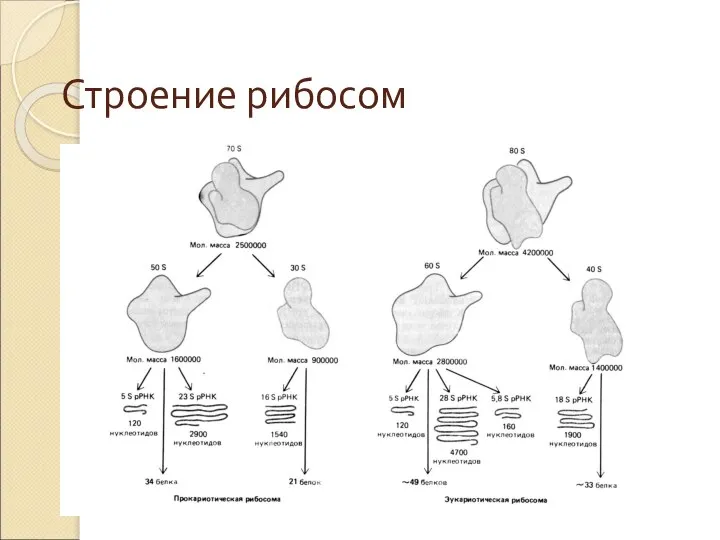
Слайд 48Строение рибосом
Строение рибосом
Слайд 49Трансляционный аппарат клетки
В рибосоме имеются три различных участка, с которыми связывается РНК:
Трансляционный аппарат клетки
В рибосоме имеются три различных участка, с которыми связывается РНК:

Слайд 50Трансляционный аппарат клетки
Участки для т-РНК называются Р -(пептидильный) и
А- (акцепторный
Трансляционный аппарат клетки
Участки для т-РНК называются Р -(пептидильный) и
А- (акцепторный

Слайд 51Трансляционный аппарат клетки
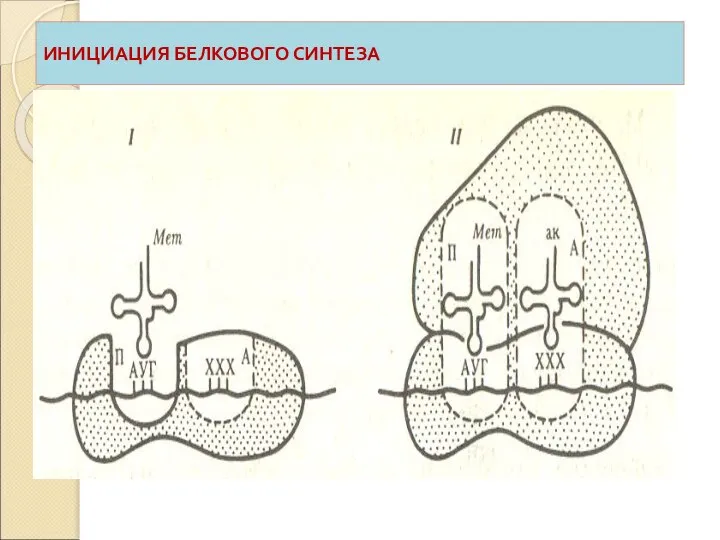
В фазе инициации субъединицы рибосомы объединяются с мРНК и в
Трансляционный аппарат клетки
В фазе инициации субъединицы рибосомы объединяются с мРНК и в

Слайд 52ИНИЦИАЦИЯ БЕЛКОВОГО СИНТЕЗА
ИНИЦИАЦИЯ БЕЛКОВОГО СИНТЕЗА
Слайд 53Рибосомы
Рибосомы

Слайд 54Трансляционный аппарат клетки
Элонгация (удлинение) – циклически повторяющиеся события, связанные с включением аминокислот
Трансляционный аппарат клетки
Элонгация (удлинение) – циклически повторяющиеся события, связанные с включением аминокислот

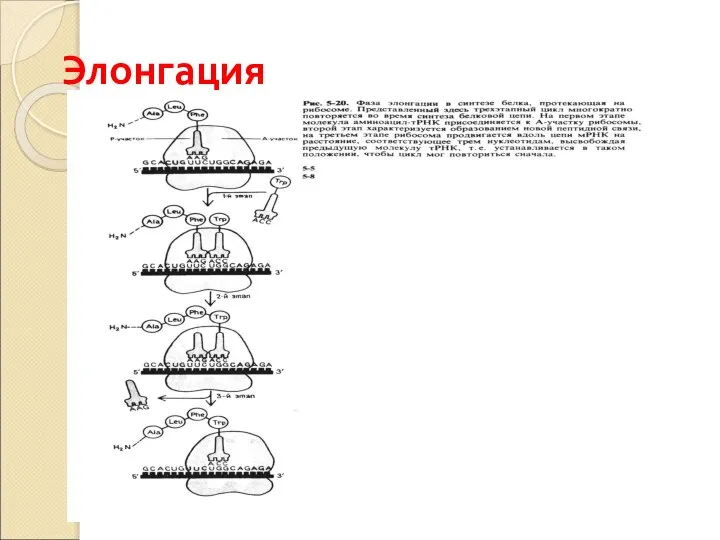
Слайд 55ФАЗА ЭЛОНГАЦИИ
При соединении антикодона т-РНК с кодоном м-РНК транспортируемая аминокислота располагается на
ФАЗА ЭЛОНГАЦИИ
При соединении антикодона т-РНК с кодоном м-РНК транспортируемая аминокислота располагается на

Слайд 56Элонгация
Элонгация
Слайд 57Трансляционный аппарат клетки
Терминация (окончание биосинтеза) связана с поступлением в рибосому одного из
Трансляционный аппарат клетки
Терминация (окончание биосинтеза) связана с поступлением в рибосому одного из

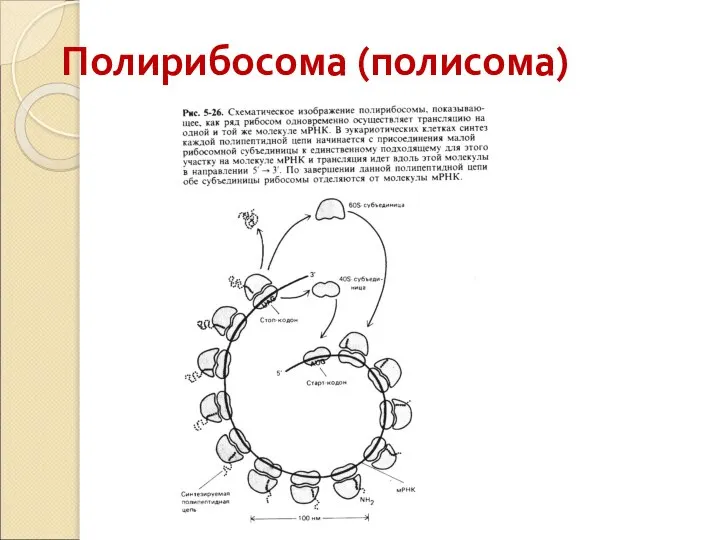
Слайд 58Полирибосома (полисома)
Полирибосома (полисома)
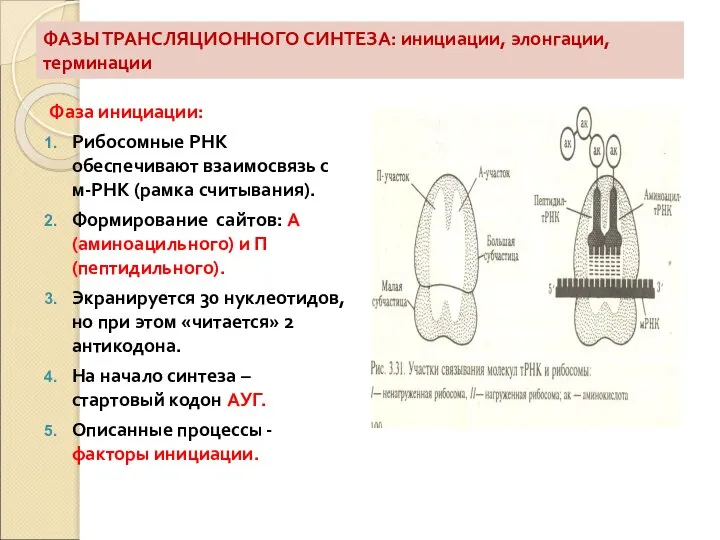
Слайд 59ФАЗЫ ТРАНСЛЯЦИОННОГО СИНТЕЗА: инициации, элонгации, терминации
Фаза инициации:
Рибосомные РНК обеспечивают взаимосвязь с м-РНК
ФАЗЫ ТРАНСЛЯЦИОННОГО СИНТЕЗА: инициации, элонгации, терминации
Фаза инициации:
Рибосомные РНК обеспечивают взаимосвязь с м-РНК
Слайд 60Белки в эволюции и онтогенезе
Бактериальные и-РНК полицистронны, т.е.кодируют несколько белков по одной
Белки в эволюции и онтогенезе
Бактериальные и-РНК полицистронны, т.е.кодируют несколько белков по одной

Слайд 61Трансляционный аппарат клетки
Действие многих эффективных антибиотиков основано на подавлении биосинтеза белков.
Трансляционный аппарат клетки
Действие многих эффективных антибиотиков основано на подавлении биосинтеза белков.

Слайд 62Трансляционный аппарат клетки
Посттрансляционная модификация заключается в укладке первичной структуры белка в структуры
Трансляционный аппарат клетки
Посттрансляционная модификация заключается в укладке первичной структуры белка в структуры

Слайд 634. Особенности организации и экспрессии у эукариот и прокариот
ЕДИНСТВО ПРОИСХОЖДЕНИЯ:
Генетический материал у
4. Особенности организации и экспрессии у эукариот и прокариот
ЕДИНСТВО ПРОИСХОЖДЕНИЯ:
Генетический материал у

Слайд 64ОТЛИЧИЯ В ЭКСПРЕССИИ ГЕНОВ У ПРО- и ЭУКАРИОТ
ОТЛИЧИЯ В ЭКСПРЕССИИ ГЕНОВ У ПРО- и ЭУКАРИОТ

Слайд 65РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ У ПРОКАРИОТ МОДЕЛЬ ОПЕРОНА.
1. Начало 60-х годов 20-го века:
РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ У ПРОКАРИОТ МОДЕЛЬ ОПЕРОНА.
1. Начало 60-х годов 20-го века:

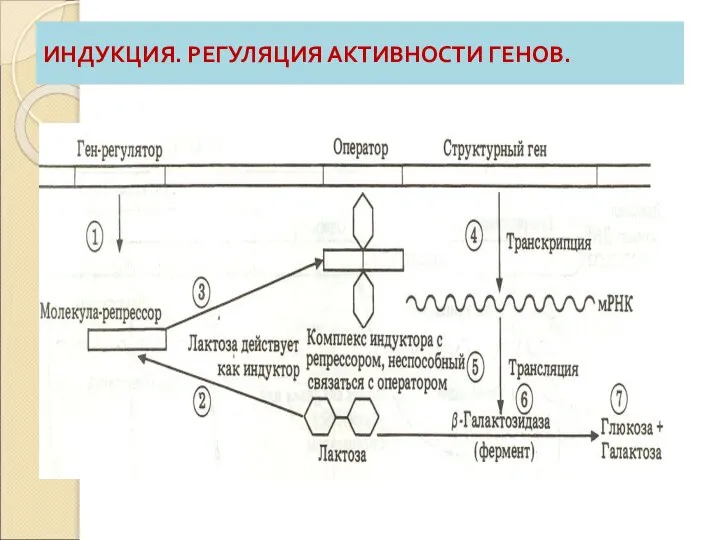
Слайд 66ИНДУКЦИЯ
4. Процесс индукции включает следующие фазы:
А) ПРОИЗВОДСТВО МОЛЕКУЛЫ-РЕПРЕССОРА:
ген-регулятор синтезирует репрессор;
при отсутствии субстрата
ИНДУКЦИЯ
4. Процесс индукции включает следующие фазы:
А) ПРОИЗВОДСТВО МОЛЕКУЛЫ-РЕПРЕССОРА:
ген-регулятор синтезирует репрессор;
при отсутствии субстрата

Слайд 67ИНДУКЦИЯ
В) Транскрипция генов и производство ферментов:
РНК-полимераза получает доступ к оператору:
синтез фермента.
ИНДУКЦИЯ
В) Транскрипция генов и производство ферментов:
РНК-полимераза получает доступ к оператору:
синтез фермента.

Слайд 68ИНДУКЦИЯ. РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ.
ИНДУКЦИЯ. РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ.
 Северные животные
Северные животные Клеточные мембраны. Транспортные системы
Клеточные мембраны. Транспортные системы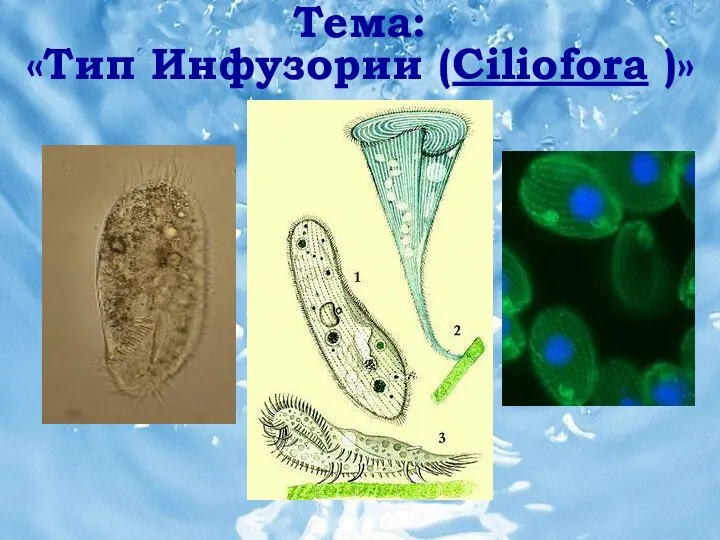 Тип инфузории (ciliofora)
Тип инфузории (ciliofora) Северо- Осетинский заповедник
Северо- Осетинский заповедник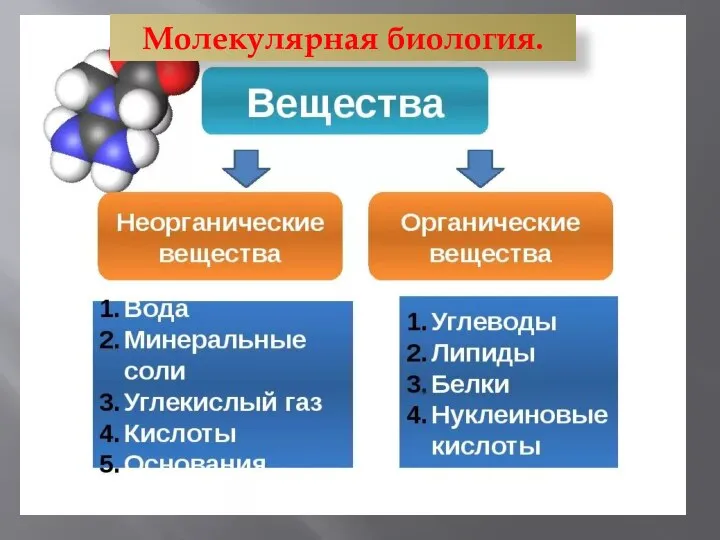 Молекулярная биология. Углеводы. Липиды
Молекулярная биология. Углеводы. Липиды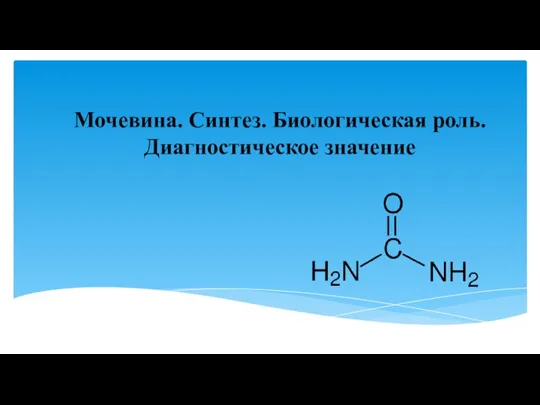 Мочевина. Синтез. Биологическая роль. Диагностическое значение
Мочевина. Синтез. Биологическая роль. Диагностическое значение Mein Fach ist Biologie
Mein Fach ist Biologie Микробиологический анализ морской воды платного и муниципального пляжей г. Сочи
Микробиологический анализ морской воды платного и муниципального пляжей г. Сочи Кто такие земноводные
Кто такие земноводные Отряды насекомых таракановые, прямокрылые, уховёртки, подёнки
Отряды насекомых таракановые, прямокрылые, уховёртки, подёнки Топ-5 самых больших бабочек в мире
Топ-5 самых больших бабочек в мире 1
1 Презентация по биологии на тему _Этапы развития жизни на Земле_
Презентация по биологии на тему _Этапы развития жизни на Земле_ Лисы
Лисы Разноцветный мир растений. 5 класс
Разноцветный мир растений. 5 класс Окружающая среда
Окружающая среда Транспортные системы организма
Транспортные системы организма Основы геоботаники (фитоценологии). Экосистема. Биогеоценоз (лекция 1)
Основы геоботаники (фитоценологии). Экосистема. Биогеоценоз (лекция 1) Хромосомная теория наследственности
Хромосомная теория наследственности Цветок. Строение, формы цветоложа
Цветок. Строение, формы цветоложа Эволюция органического мира. Антропогенез. 11 класс
Эволюция органического мира. Антропогенез. 11 класс 文:[美]史蒂夫・危金斯
文:[美]史蒂夫・危金斯 Метод сравнительной геномной гибридизации
Метод сравнительной геномной гибридизации Систематика растений Архегониальные растения. Моховидные
Систематика растений Архегониальные растения. Моховидные Рослини-мандрівники
Рослини-мандрівники Корнепло́ды — часть растения, запасающая питательные вещества, ассоциированные с корневой системой
Корнепло́ды — часть растения, запасающая питательные вещества, ассоциированные с корневой системой 15. Тип Кольчатые черви. легкий (1)
15. Тип Кольчатые черви. легкий (1) Адамның жыныс жүйесінің құрылымы мен қызметі
Адамның жыныс жүйесінің құрылымы мен қызметі