- Активизация эфринов путем их кластеризации

Содержание
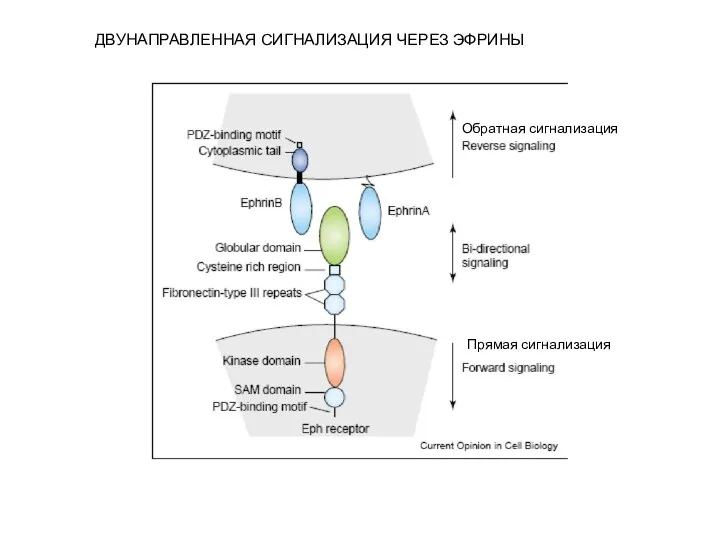
- 2. ДВУНАПРАВЛЕННАЯ СИГНАЛИЗАЦИЯ ЧЕРЕЗ ЭФРИНЫ Прямая сигнализация Обратная сигнализация
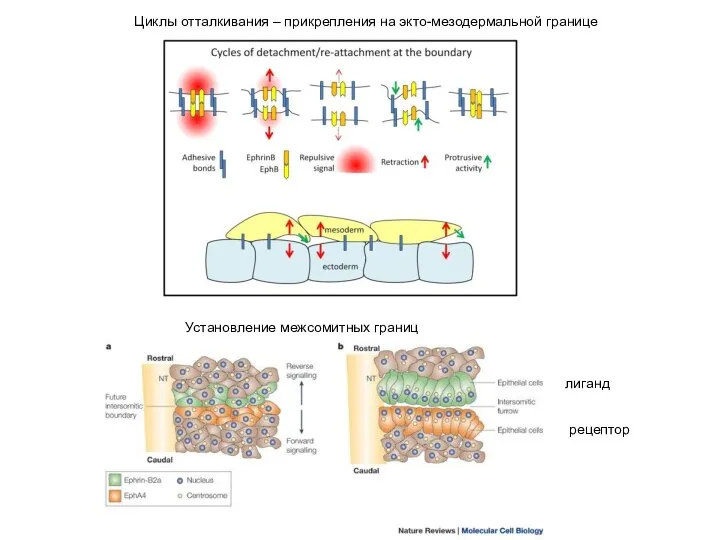
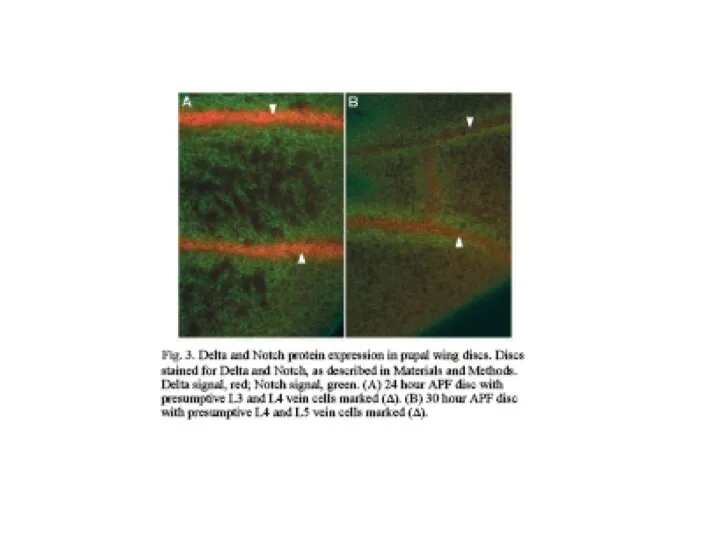
- 3. Циклы отталкивания – прикрепления на экто-мезодермальной границе Установление межсомитных границ лиганд рецептор
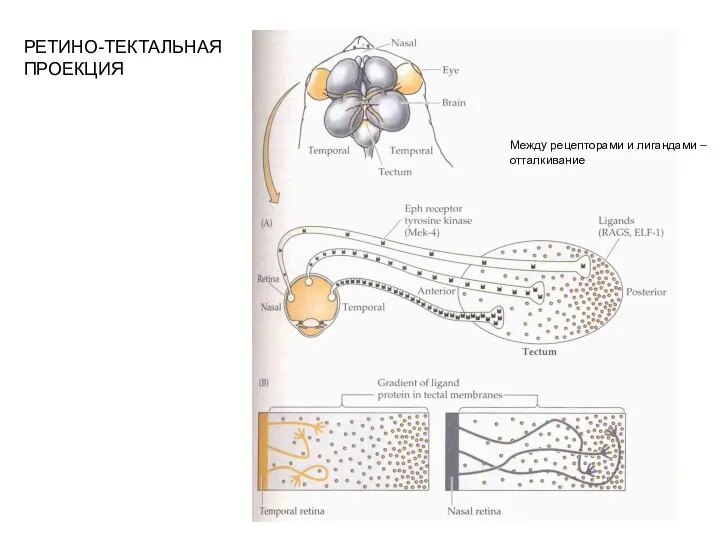
- 4. РЕТИНО-ТЕКТАЛЬНАЯ ПРОЕКЦИЯ Между рецепторами и лигандами – отталкивание
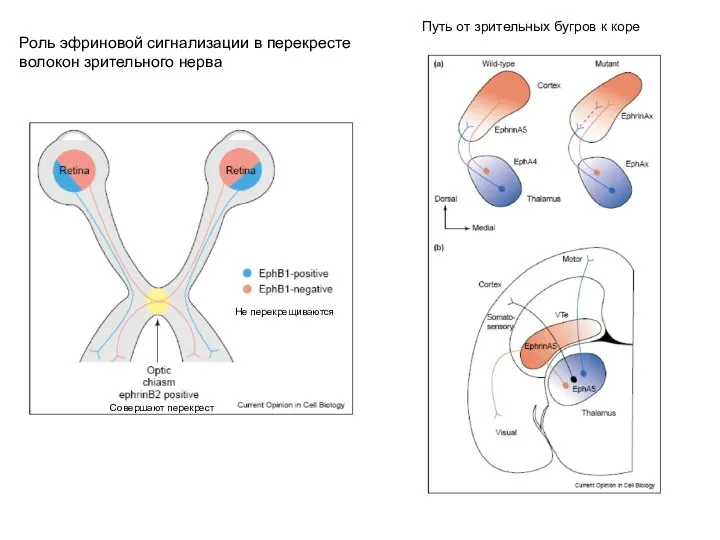
- 5. Роль эфриновой сигнализации в перекресте волокон зрительного нерва Путь от зрительных бугров к коре Совершают перекрест
- 6. СИГНАЛИЗАЦИЯ DELTA – NOTCH: ВЗАИМОДЕЙСТВИЕ МЕМБРАННО-СВЯЗАННОГО ЛИГАНДА DELTA (КРАСНЫЙ) НА ПОВЕРХНОСТИ БУДУЩИХ НЕРВНЫХ КЛЕТОК С РЕЦЕПТОРОМ
- 9. Delta-Notch signal WNT signal Effective non-effective DELTA-NOTCH и Wnt сигнализация в раннем развитии C. elegans E
- 10. DELTA-NOTCH ИНДУКЦИИ В ПОЗДНЕМ РАЗВИТИИ C.ELEGANS Для клеток левой стороны головы необходима индукция
- 11. Figure 9. A working model for the involvement of DELTA-NOTCH signalling in the transition from proliferation
- 12. МЕХАНИЧЕСКИЕ СИЛЫ, ДЕЙСТВУЮЩИЕ НА КЛЕТКИ
- 13. Натяжение предотвращает апоптоз МЕХАНИЧЕСКИЕ НАПРЯЖЕНИЯ КАК ФАКТОР ВЫЖИВАНИЯ И ДИФФЕРЕНЦИРОВКИ: КЛЕТКИ НА «ОСТРОВКАХ» Chen et al.
- 14. РАСТЯЖЕНИЕ СПОСОБСТВУЕТ РОСТУ, СЖАТИЕ ПОДАВЛЯЕТ РОСТ ЧРЕЗМЕРНЫЕ НАПРЯЖЕНИЯ ВЕДУТ К АПОПТОЗУ
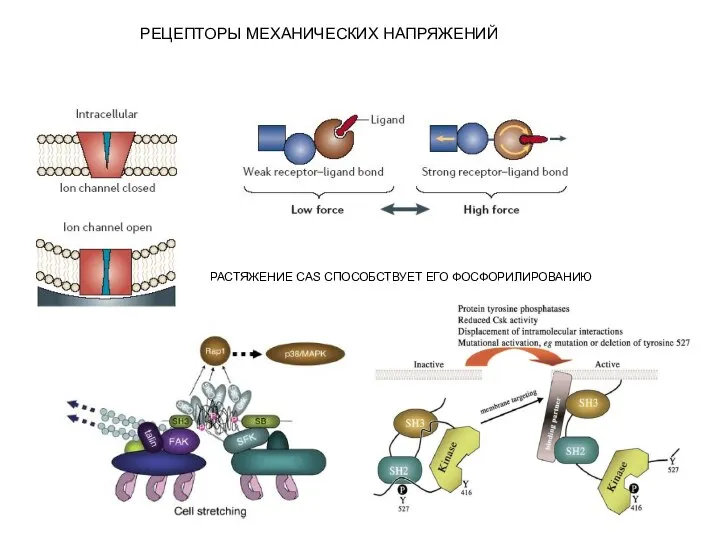
- 16. РАСТЯЖЕНИЕ CAS СПОСОБСТВУЕТ ЕГО ФОСФОРИЛИРОВАНИЮ РЕЦЕПТОРЫ МЕХАНИЧЕСКИХ НАПРЯЖЕНИЙ
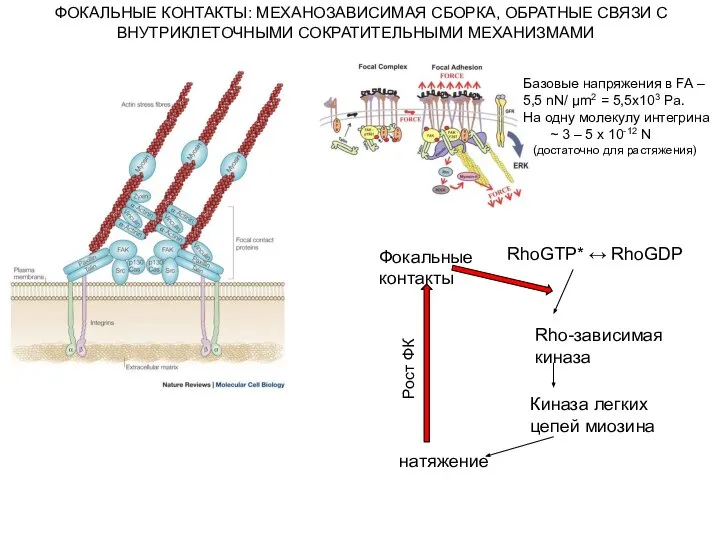
- 17. Базовые напряжения в FA – 5,5 nN/ µm2 = 5,5x103 Pa. На одну молекулу интегрина ~
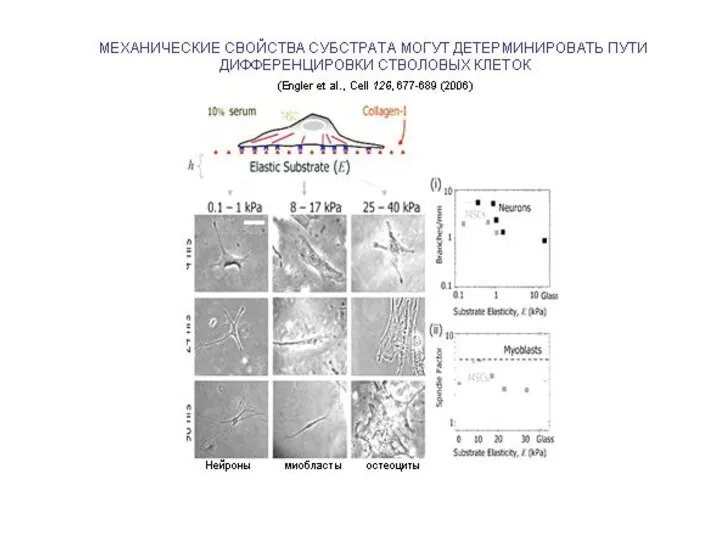
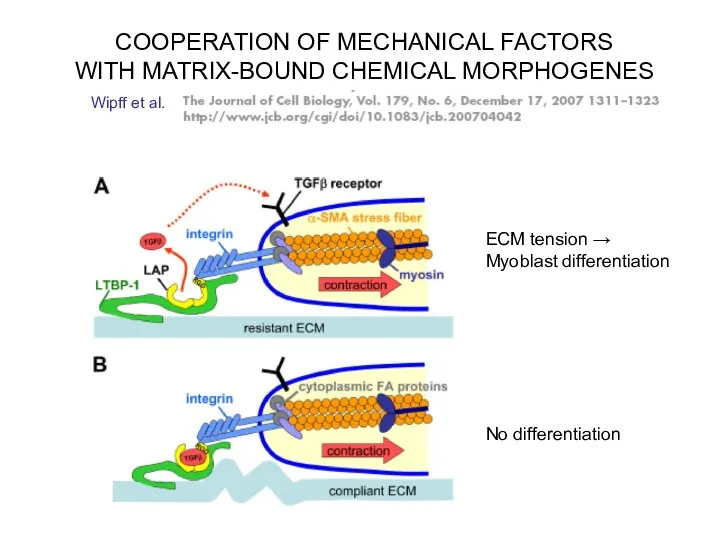
- 18. COOPERATION OF MECHANICAL FACTORS WITH MATRIX-BOUND CHEMICAL MORPHOGENES Wipff et al. ECM tension → Myoblast differentiation
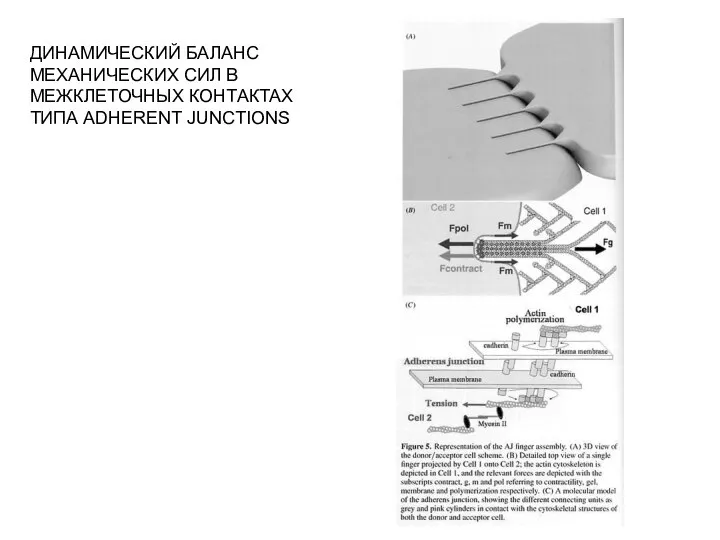
- 19. ДИНАМИЧЕСКИЙ БАЛАНС МЕХАНИЧЕСКИХ СИЛ В МЕЖКЛЕТОЧНЫХ КОНТАКТАХ ТИПА ADHERENT JUNCTIONS
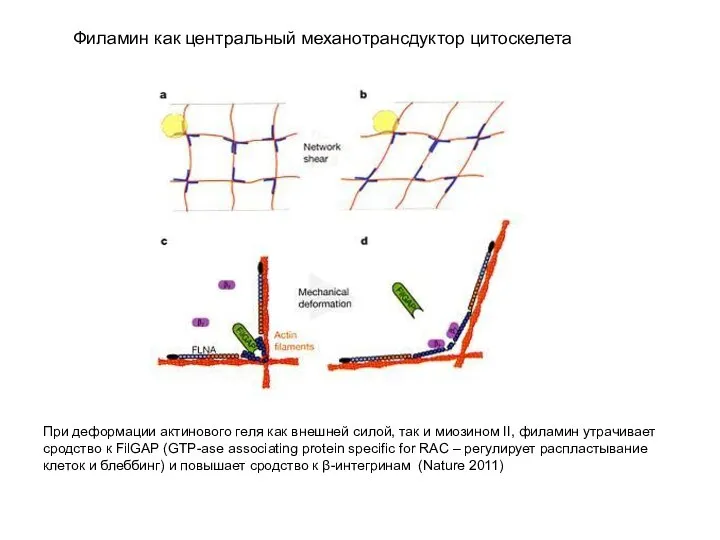
- 20. Филамин как центральный механотрансдуктор цитоскелета При деформации актинового геля как внешней силой, так и миозином II,
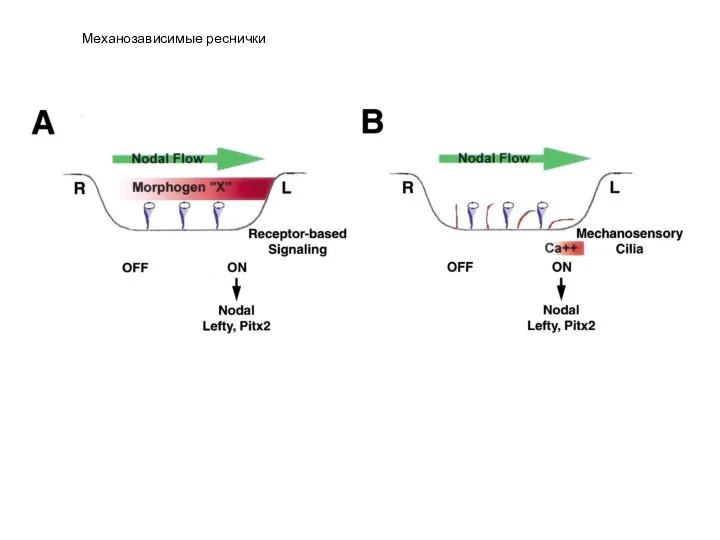
- 21. Механозависимые реснички
- 24. СИГНАЛЬНЫЕ ПУТИ, ИДУЩИЕ ОТ ФОКАЛЬНЫХ КОНТАКТОВ МЕХАНИЧЕСКОЕ РАСТЯЖЕНИЕ УСИЛИВАЕТ ТИРОЗИН-ФОСФОРИЛИРОВАНИЕ p130 CAS nesprin Промежут филамент актин
- 26. Скачать презентацию
Слайд 3Циклы отталкивания – прикрепления на экто-мезодермальной границе
Установление межсомитных границ
лиганд
рецептор
Циклы отталкивания – прикрепления на экто-мезодермальной границе
Установление межсомитных границ
лиганд
рецептор
Слайд 4РЕТИНО-ТЕКТАЛЬНАЯ
ПРОЕКЦИЯ
Между рецепторами и лигандами –
отталкивание
РЕТИНО-ТЕКТАЛЬНАЯ
ПРОЕКЦИЯ
Между рецепторами и лигандами –
отталкивание
Слайд 5Роль эфриновой сигнализации в перекресте
волокон зрительного нерва
Путь от зрительных бугров
Роль эфриновой сигнализации в перекресте
волокон зрительного нерва
Путь от зрительных бугров
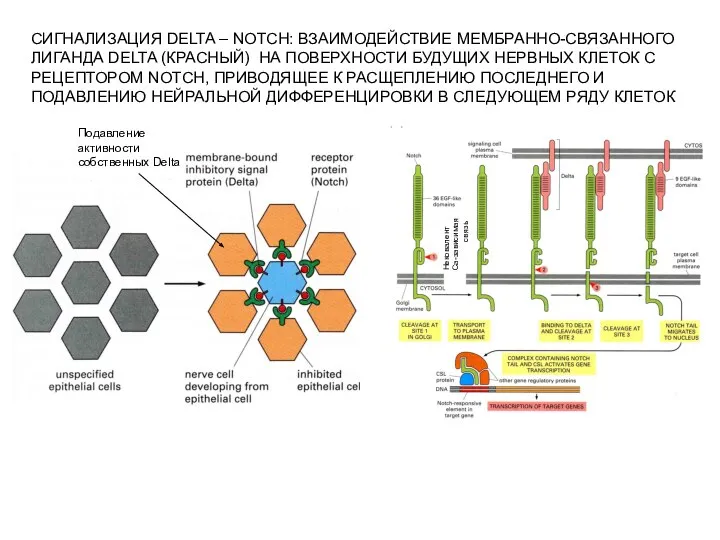
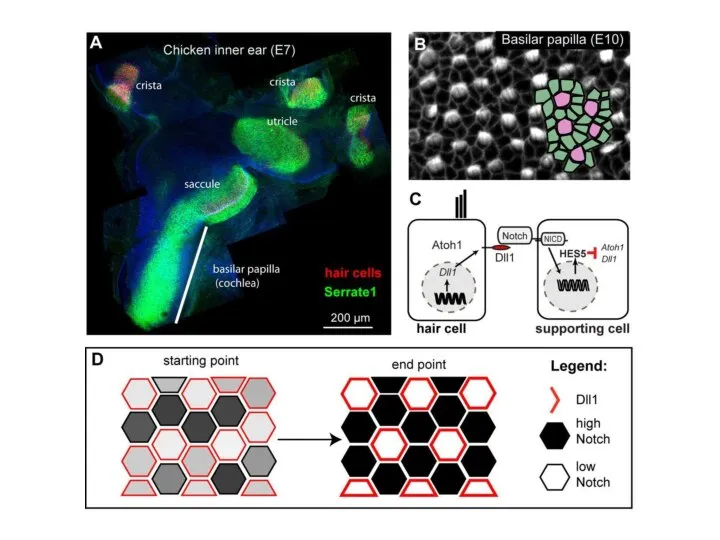
Слайд 6СИГНАЛИЗАЦИЯ DELTA – NOTCH: ВЗАИМОДЕЙСТВИЕ МЕМБРАННО-СВЯЗАННОГО
ЛИГАНДА DELTA (КРАСНЫЙ) НА ПОВЕРХНОСТИ БУДУЩИХ НЕРВНЫХ
СИГНАЛИЗАЦИЯ DELTA – NOTCH: ВЗАИМОДЕЙСТВИЕ МЕМБРАННО-СВЯЗАННОГО
ЛИГАНДА DELTA (КРАСНЫЙ) НА ПОВЕРХНОСТИ БУДУЩИХ НЕРВНЫХ
Слайд 9Delta-Notch signal
WNT signal
Effective non-effective
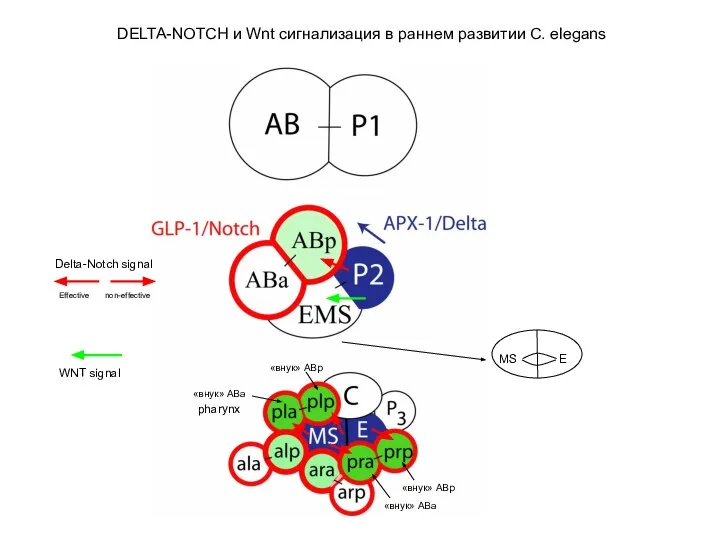
DELTA-NOTCH и Wnt сигнализация в раннем развитии C. elegans
E
MS
«внук»
Delta-Notch signal
WNT signal
Effective non-effective
DELTA-NOTCH и Wnt сигнализация в раннем развитии C. elegans
E
MS
«внук»
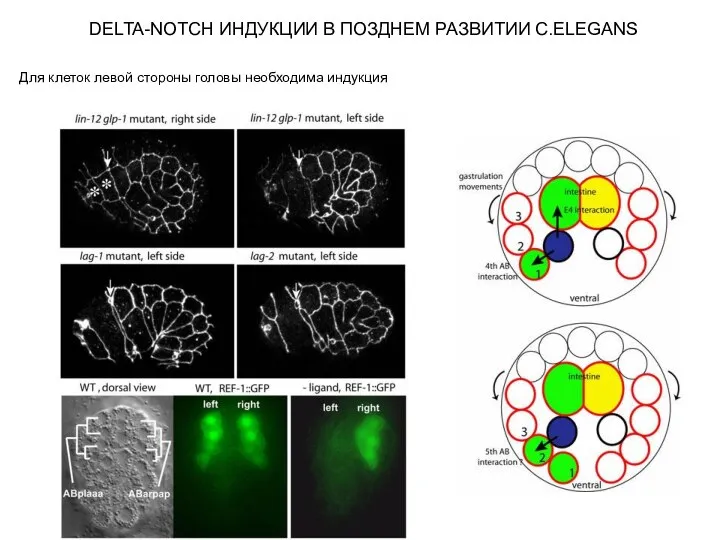
Слайд 10DELTA-NOTCH ИНДУКЦИИ В ПОЗДНЕМ РАЗВИТИИ C.ELEGANS
Для клеток левой стороны головы необходима индукция
DELTA-NOTCH ИНДУКЦИИ В ПОЗДНЕМ РАЗВИТИИ C.ELEGANS
Для клеток левой стороны головы необходима индукция
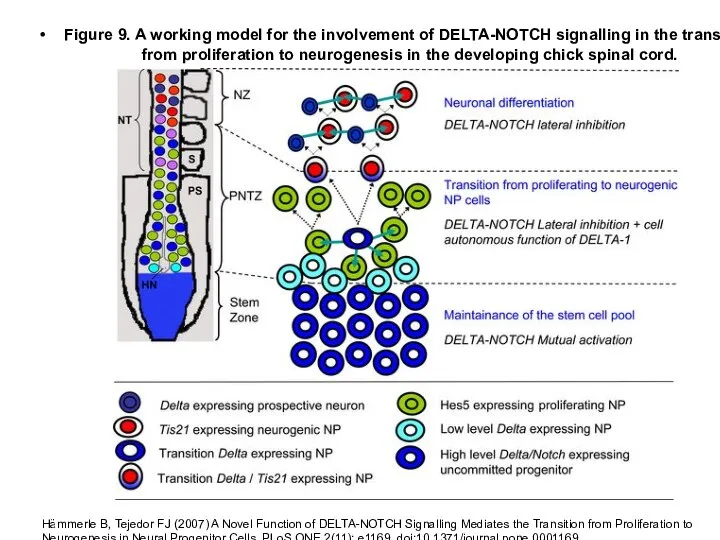
Слайд 11Figure 9. A working model for the involvement of DELTA-NOTCH signalling in
Figure 9. A working model for the involvement of DELTA-NOTCH signalling in
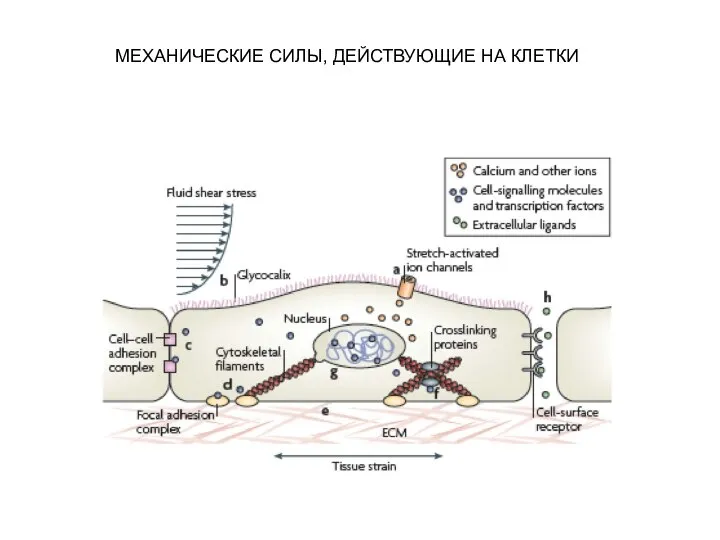
Слайд 12МЕХАНИЧЕСКИЕ СИЛЫ, ДЕЙСТВУЮЩИЕ НА КЛЕТКИ
МЕХАНИЧЕСКИЕ СИЛЫ, ДЕЙСТВУЮЩИЕ НА КЛЕТКИ
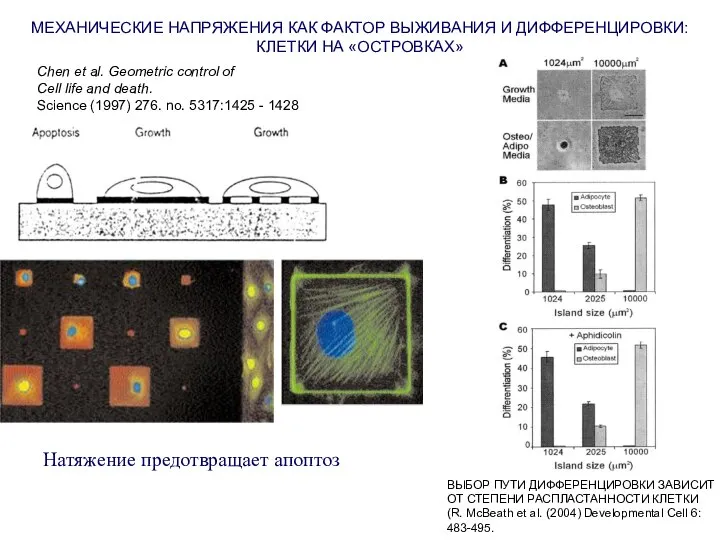
Слайд 13Натяжение предотвращает апоптоз
МЕХАНИЧЕСКИЕ НАПРЯЖЕНИЯ КАК ФАКТОР ВЫЖИВАНИЯ И ДИФФЕРЕНЦИРОВКИ:
КЛЕТКИ НА «ОСТРОВКАХ»
Chen et
Натяжение предотвращает апоптоз
МЕХАНИЧЕСКИЕ НАПРЯЖЕНИЯ КАК ФАКТОР ВЫЖИВАНИЯ И ДИФФЕРЕНЦИРОВКИ:
КЛЕТКИ НА «ОСТРОВКАХ»
Chen et
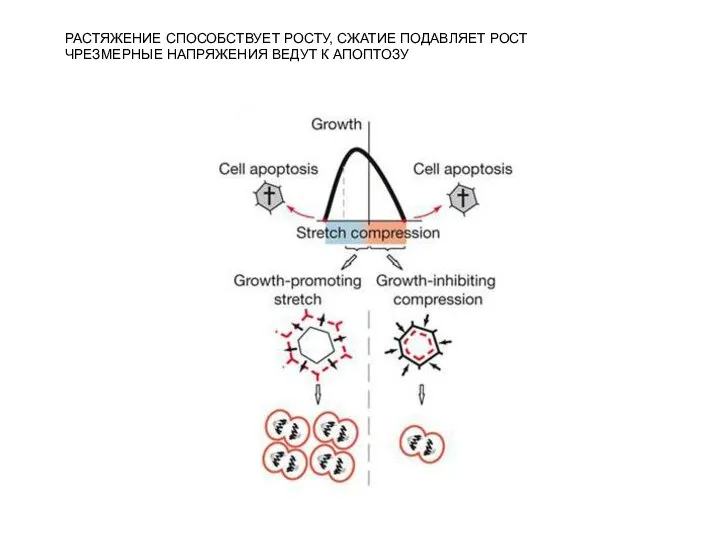
Слайд 14РАСТЯЖЕНИЕ СПОСОБСТВУЕТ РОСТУ, СЖАТИЕ ПОДАВЛЯЕТ РОСТ
ЧРЕЗМЕРНЫЕ НАПРЯЖЕНИЯ ВЕДУТ К АПОПТОЗУ
РАСТЯЖЕНИЕ СПОСОБСТВУЕТ РОСТУ, СЖАТИЕ ПОДАВЛЯЕТ РОСТ
ЧРЕЗМЕРНЫЕ НАПРЯЖЕНИЯ ВЕДУТ К АПОПТОЗУ
Слайд 16РАСТЯЖЕНИЕ CAS СПОСОБСТВУЕТ ЕГО ФОСФОРИЛИРОВАНИЮ
РЕЦЕПТОРЫ МЕХАНИЧЕСКИХ НАПРЯЖЕНИЙ
РАСТЯЖЕНИЕ CAS СПОСОБСТВУЕТ ЕГО ФОСФОРИЛИРОВАНИЮ
РЕЦЕПТОРЫ МЕХАНИЧЕСКИХ НАПРЯЖЕНИЙ
Слайд 17Базовые напряжения в FA –
5,5 nN/ µm2 = 5,5x103 Pa.
На одну молекулу
Базовые напряжения в FA –
5,5 nN/ µm2 = 5,5x103 Pa.
На одну молекулу
Слайд 18COOPERATION OF MECHANICAL FACTORS
WITH MATRIX-BOUND CHEMICAL MORPHOGENES
Wipff et al.
ECM tension →
Myoblast
COOPERATION OF MECHANICAL FACTORS
WITH MATRIX-BOUND CHEMICAL MORPHOGENES
Wipff et al.
ECM tension →
Myoblast
Слайд 19ДИНАМИЧЕСКИЙ БАЛАНС
МЕХАНИЧЕСКИХ СИЛ В
МЕЖКЛЕТОЧНЫХ КОНТАКТАХ
ТИПА ADHERENT JUNCTIONS
ДИНАМИЧЕСКИЙ БАЛАНС
МЕХАНИЧЕСКИХ СИЛ В
МЕЖКЛЕТОЧНЫХ КОНТАКТАХ
ТИПА ADHERENT JUNCTIONS
Слайд 20Филамин как центральный механотрансдуктор цитоскелета
При деформации актинового геля как внешней силой, так
Филамин как центральный механотрансдуктор цитоскелета
При деформации актинового геля как внешней силой, так
Слайд 21Механозависимые реснички
Механозависимые реснички
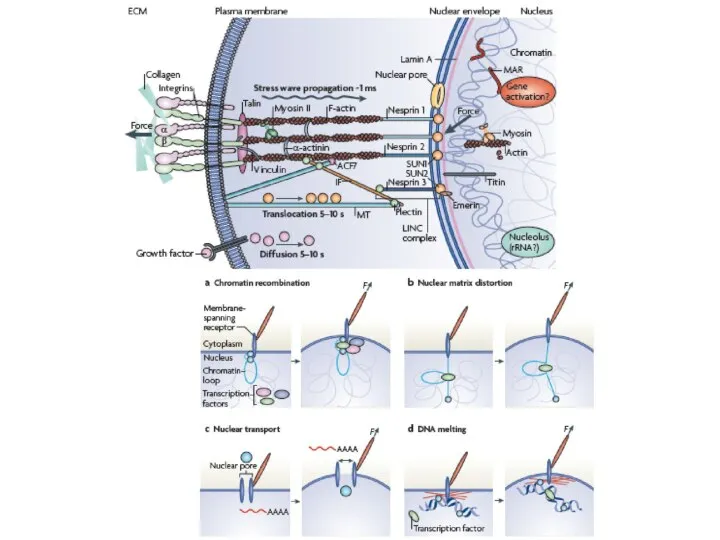
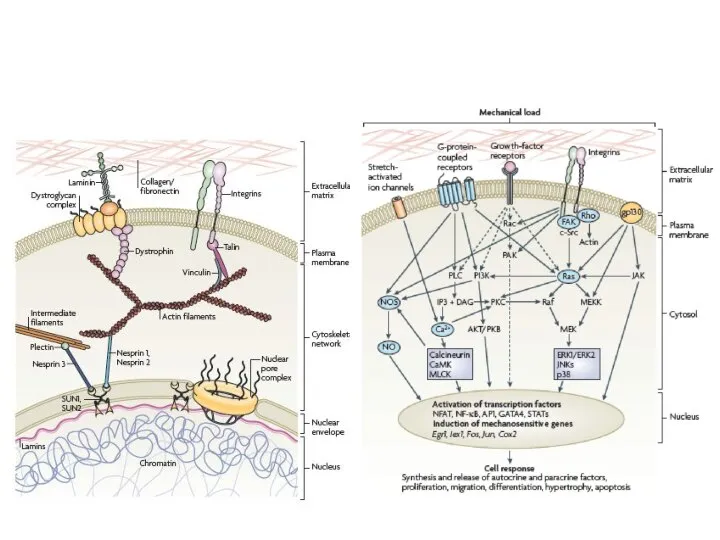
Слайд 24СИГНАЛЬНЫЕ ПУТИ, ИДУЩИЕ ОТ ФОКАЛЬНЫХ КОНТАКТОВ
МЕХАНИЧЕСКОЕ РАСТЯЖЕНИЕ УСИЛИВАЕТ
ТИРОЗИН-ФОСФОРИЛИРОВАНИЕ p130 CAS
nesprin
Промежут филамент
актин
ламины
хроматин
МЕХАНИЧЕСКИЕ СИГНАЛЫ
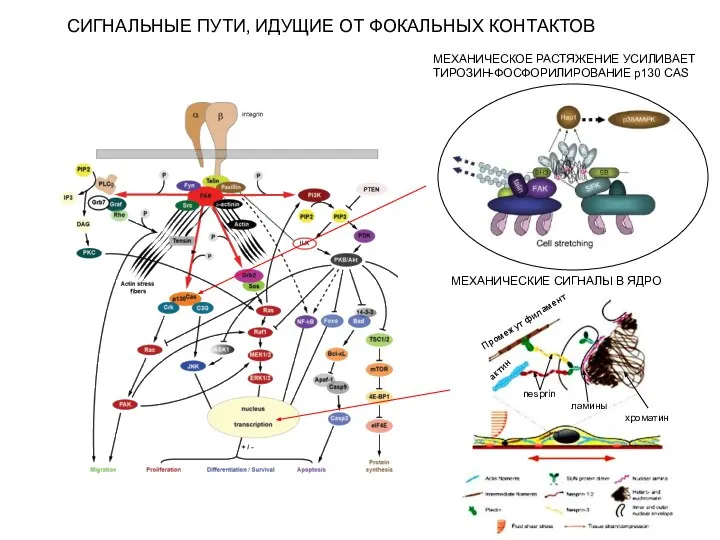
СИГНАЛЬНЫЕ ПУТИ, ИДУЩИЕ ОТ ФОКАЛЬНЫХ КОНТАКТОВ
МЕХАНИЧЕСКОЕ РАСТЯЖЕНИЕ УСИЛИВАЕТ
ТИРОЗИН-ФОСФОРИЛИРОВАНИЕ p130 CAS
nesprin
Промежут филамент
актин
ламины
хроматин
МЕХАНИЧЕСКИЕ СИГНАЛЫ
 Скат Манта
Скат Манта Белый медведь. 1 класс
Белый медведь. 1 класс Лист. Строение и функции
Лист. Строение и функции Рост и развитие растений
Рост и развитие растений Образование половых клеток и оплодотворение
Образование половых клеток и оплодотворение Отряд ластоногие
Отряд ластоногие Прямое и непрямое деление клетки
Прямое и непрямое деление клетки Гаметогенез, эмбриогенез
Гаметогенез, эмбриогенез Турне в огород. Викторина о культурных растениях
Турне в огород. Викторина о культурных растениях Водоросли
Водоросли Кровососущие насекомые
Кровососущие насекомые Подборка заданий линии 23 (задания 2 части ЕГЭ с рисунком)
Подборка заданий линии 23 (задания 2 части ЕГЭ с рисунком) Изменчивость. Наследственная изменчивость
Изменчивость. Наследственная изменчивость Времена года
Времена года Predshestvenniki_cheloveka(1)
Predshestvenniki_cheloveka(1) Кумкват. Характеристика кумквата
Кумкват. Характеристика кумквата Викторина Мир вокруг
Викторина Мир вокруг Овуляція. Що таке овуляція
Овуляція. Що таке овуляція Самые необыкновенные морские рыбки
Самые необыкновенные морские рыбки За исследование каротиноидов и флавинов, а также за изучение витаминов А и В2
За исследование каротиноидов и флавинов, а также за изучение витаминов А и В2 Күріштің шығу тарихы. Күріштің Қазақстандағы сорттары. Қазақстандағы күріштің ген қоры және ұлттық коллекциясы
Күріштің шығу тарихы. Күріштің Қазақстандағы сорттары. Қазақстандағы күріштің ген қоры және ұлттық коллекциясы Презентация по биологии _Простейшие_
Презентация по биологии _Простейшие_ Презентация на тему Агроэкосистемы
Презентация на тему Агроэкосистемы  Морские уточки
Морские уточки Животный мир. Кто как готовится к зиме
Животный мир. Кто как готовится к зиме Augi vasarā
Augi vasarā Спинной мозг
Спинной мозг Лікарські рослини, які використовує в побуті моя сім’я
Лікарські рослини, які використовує в побуті моя сім’я