Физиология как наука. Биоэлектрические явления в возбудимых тканях. Физиология синапсов. Межнейронные связи
- Физиология как наука. Биоэлектрические явления в возбудимых тканях. Физиология синапсов. Межнейронные связи

Содержание
- 2. Физиология - наука о закономерностях жизнедеятельности организма во взаимосвязи с внешней средой. Различают следующие физиологические дисциплины:
- 4. Рудольф Гейденгайн (1834-1897) , выдающийся немецкий физиолог и гистолог . Основные работы посвящены физиологии обмена веществ
- 5. Английский врач Уильям Гарвей (1578-1657) на основании своих экспериментов описал работу сердца , малое и большое
- 6. Введенский Николай Евгеньевич (1852-1922) , русский физиолог . Впервые в 1884 году прослушал с помощью телефонного
- 7. Строение клеточной мембраны внешняя поверхность
- 8. Основные виды транспорта 1. Пассивный транспорт: простая диффузия облегченная диффузия осмос фильтрация 2. Активный транспорт: первичный
- 9. Основные механизмы транспорта
- 10. Ворота белкового канала обеспечивают контроль ионной проницаемости
- 11. Активный транспорт (Механизм работы Na + - K + насоса)
- 12. Экзоцытоз та эндоцытоз
- 14. Любая нервная клетка организма ограничена липопротеиновой мембраной, которая является хорошим электрическим изолятором. Если внутрь клетки ввести
- 15. Мембранный потенциал покоя - это разница биоэлектрических потенциалов между наружной и внутренней поверхностью мембраны , которая
- 16. Виды раздражителей (по природе): химические (растворы кислот, щелочей, солей, органических соединений), механические (удар, сжатие, укол), температурные
- 17. После воздействия допорогового раздражителя на мембрану , в месте ее раздражения возникает деполяризация . Эти изменения
- 18. Потенциал действия нейрона 1 - мембранный потенциал покоя, 2 - локальный ответ 3 - критический уровень
- 20. Співвідношення фаз потенціалу дії (А) та збудливості (В)
- 21. Условия и законы проведения возбуждения аксонами Условия: 1. Анатомическая целостность нервного волокна. Травма, перерезка нерва нарушает
- 22. Аксон Миелиновая оболочка Шванновских клеток Перехват Ранвье является Ядро шванновских клеток Строение миелиновых волокон
- 23. Проведение возбуждения безмиелиновыми и миелиновыми нервными волокнами Безмиелиновых нервными волокнами возбуждения распространяется непрерывно, а миелиновимы от
- 25. Послідовність процесів скорочення та розслаблення м'язів Сокращение Генерация ПД мотонейроном Высвобождение ацетилхолина в концевой пластинке Связывание
- 26. Суммация сокращений и тетанус мышцы В ответ на одинокий импульс происходит сокращение , по которому наблюдается
- 27. Нейромоторного фазные (а) и тонические (б) единицы
- 28. Одинокие сокращения (а), зубчатый (б) и гладкий (в) тетанус. Зубчатый тетанус возникает при такой частоте, когда
- 29. Быстрый потенциал действия кардиомиоцитов Фаза 0 - быстрый вход Na + в клетку; Фаза 1 -
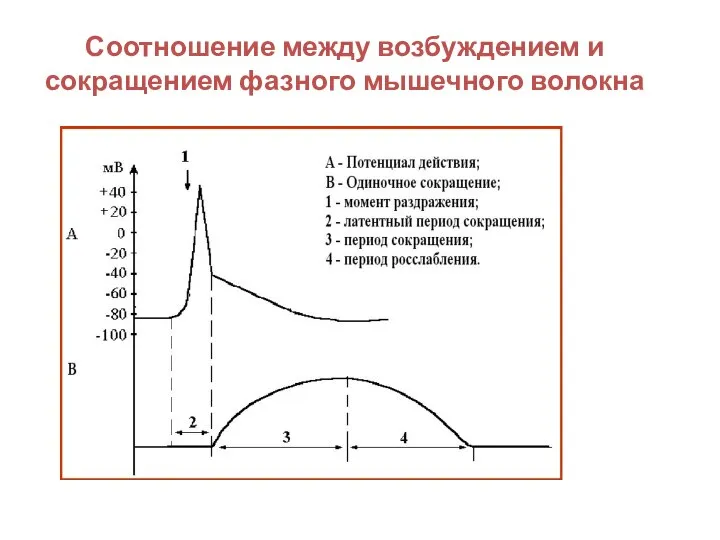
- 30. Потенциал действия фазного мышечного волокна
- 31. Соотношение между возбуждением и сокращением фазного мышечного волокна
- 32. Классификация рецепторов 1. По строению и физиологическими свойствами : ▪ первичные (первичночуствительные); ▪ вторичные (вторичночуствительные). 2.
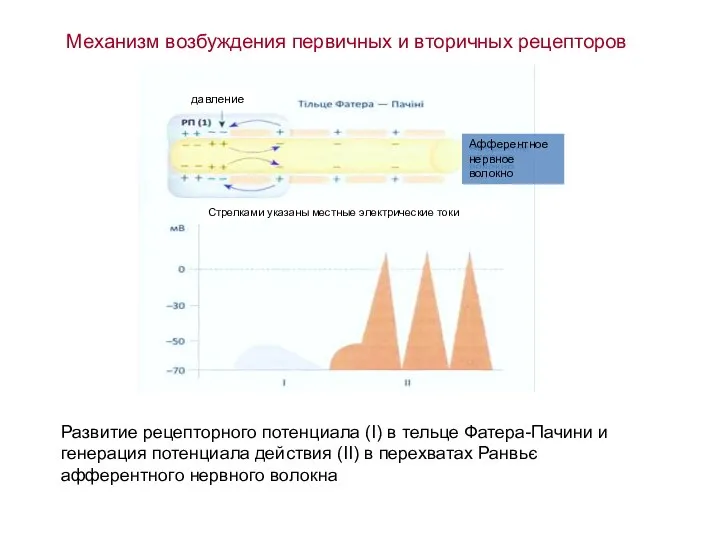
- 33. Механизм возбуждения первичных и вторичных рецепторов Развитие рецепторного потенциала (I) в тельце Фатера-Пачини и генерация потенциала
- 34. Понятие о рецепторе, рецепторном участке Главной частью периферического отдела сенсорных систем является рецепторы - "окна" ЦНС.
- 35. Функции рецептора : восприятие раздражения , его первичное различение , формирования кода . Кодирование информации -
- 37. Скачать презентацию

Слайд 4Рудольф Гейденгайн (1834-1897) , выдающийся немецкий физиолог и гистолог . Основные работы
Рудольф Гейденгайн (1834-1897) , выдающийся немецкий физиолог и гистолог . Основные работы

Слайд 5Английский врач Уильям Гарвей (1578-1657) на основании своих экспериментов описал работу сердца
Английский врач Уильям Гарвей (1578-1657) на основании своих экспериментов описал работу сердца

Слайд 6Введенский Николай Евгеньевич (1852-1922) , русский физиолог . Впервые в 1884 году
Введенский Николай Евгеньевич (1852-1922) , русский физиолог . Впервые в 1884 году

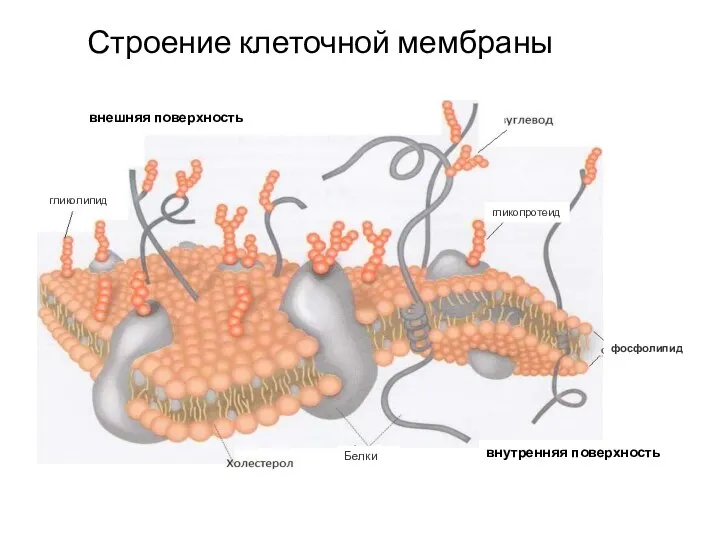
Слайд 7Строение клеточной мембраны
внешняя поверхность
Строение клеточной мембраны
внешняя поверхность
Слайд 8 Основные виды транспорта
1. Пассивный транспорт:
простая диффузия
облегченная диффузия
осмос
фильтрация
2. Активный транспорт:
первичный
вторичный
везикулярный механизм транспортировки (эндоцитоз,
Основные виды транспорта
1. Пассивный транспорт:
простая диффузия
облегченная диффузия
осмос
фильтрация
2. Активный транспорт:
первичный
вторичный
везикулярный механизм транспортировки (эндоцитоз,

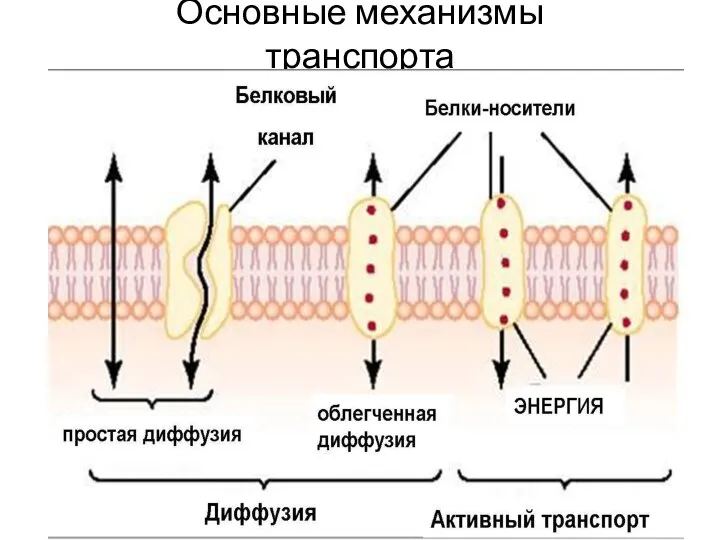
Слайд 9Основные механизмы транспорта
Основные механизмы транспорта
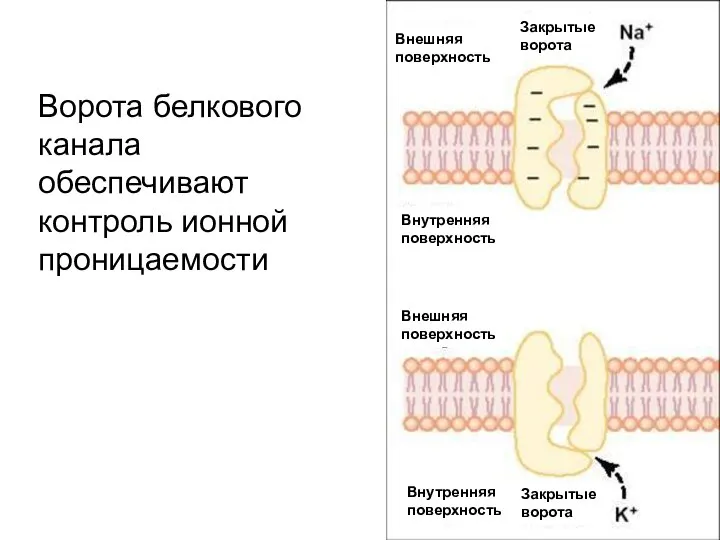
Слайд 10Ворота белкового канала обеспечивают контроль ионной проницаемости
Ворота белкового канала обеспечивают контроль ионной проницаемости
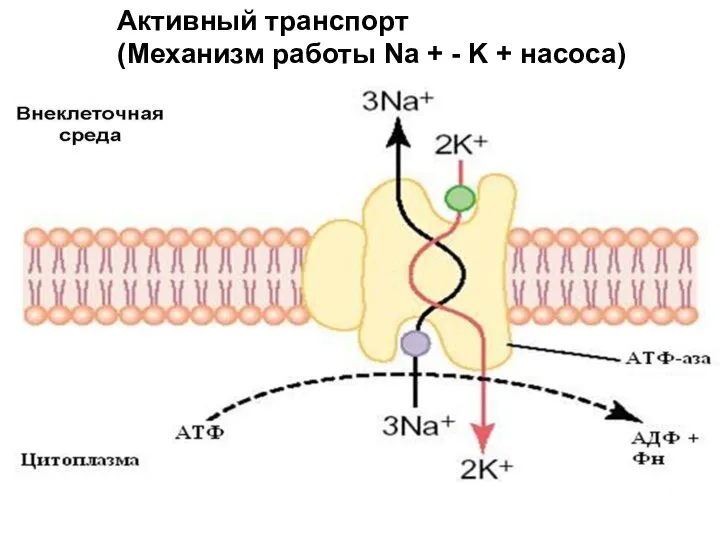
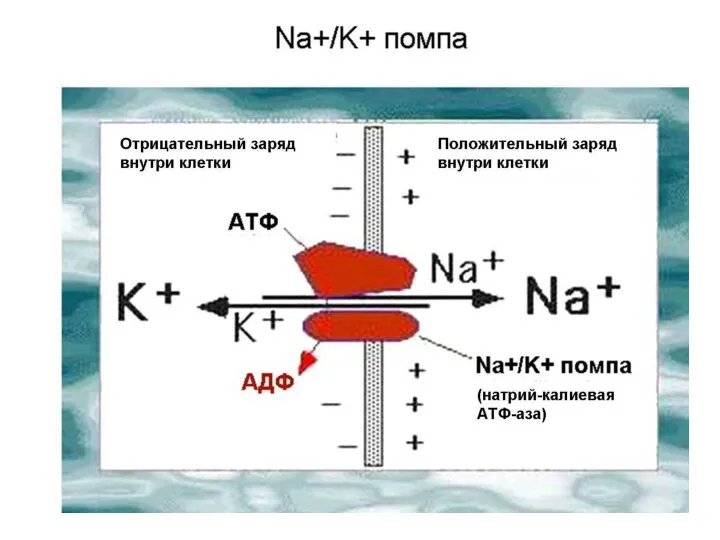
Слайд 11Активный транспорт
(Механизм работы Na + - K + насоса)
Активный транспорт
(Механизм работы Na + - K + насоса)
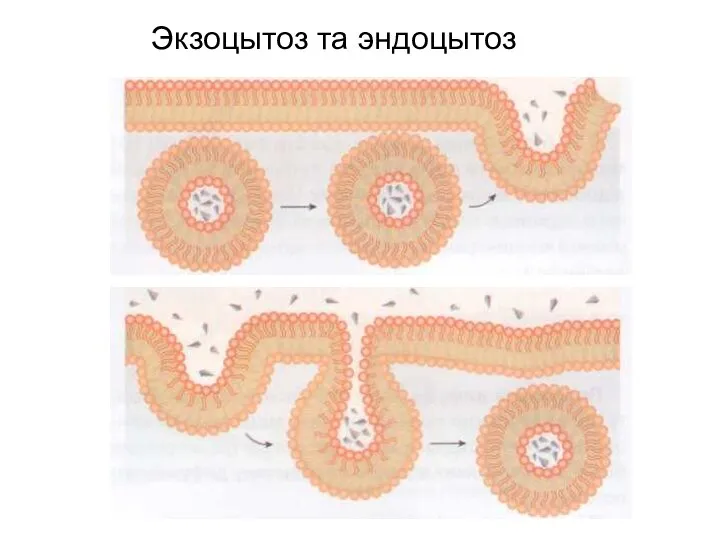
Слайд 12Экзоцытоз та эндоцытоз
Экзоцытоз та эндоцытоз
Слайд 14Любая нервная клетка организма ограничена липопротеиновой мембраной, которая является хорошим электрическим изолятором.
Любая нервная клетка организма ограничена липопротеиновой мембраной, которая является хорошим электрическим изолятором.
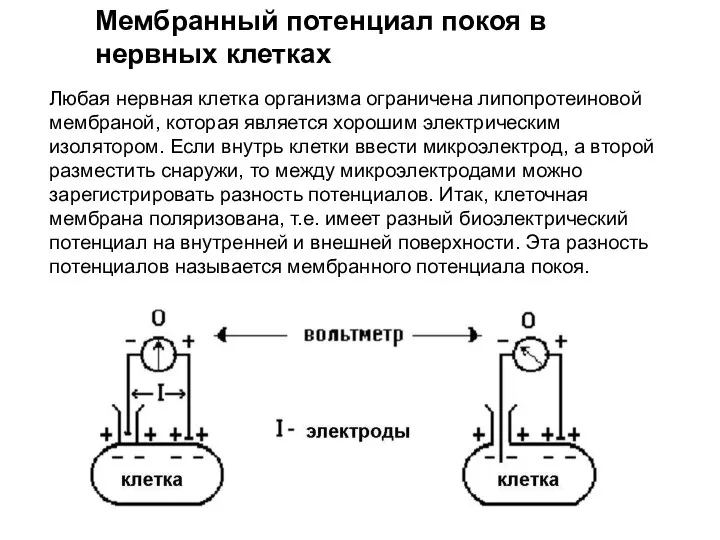
Слайд 15 Мембранный потенциал покоя - это разница биоэлектрических потенциалов между наружной и внутренней
Мембранный потенциал покоя - это разница биоэлектрических потенциалов между наружной и внутренней

Слайд 16Виды раздражителей (по природе):
химические (растворы кислот, щелочей, солей, органических соединений),
механические (удар, сжатие,
Виды раздражителей (по природе):
химические (растворы кислот, щелочей, солей, органических соединений),
механические (удар, сжатие,

Слайд 17 После воздействия допорогового раздражителя на мембрану , в месте ее раздражения возникает
После воздействия допорогового раздражителя на мембрану , в месте ее раздражения возникает

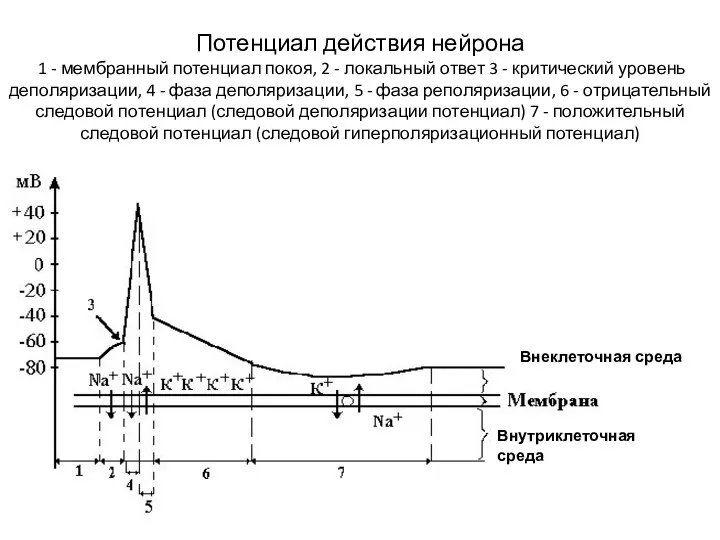
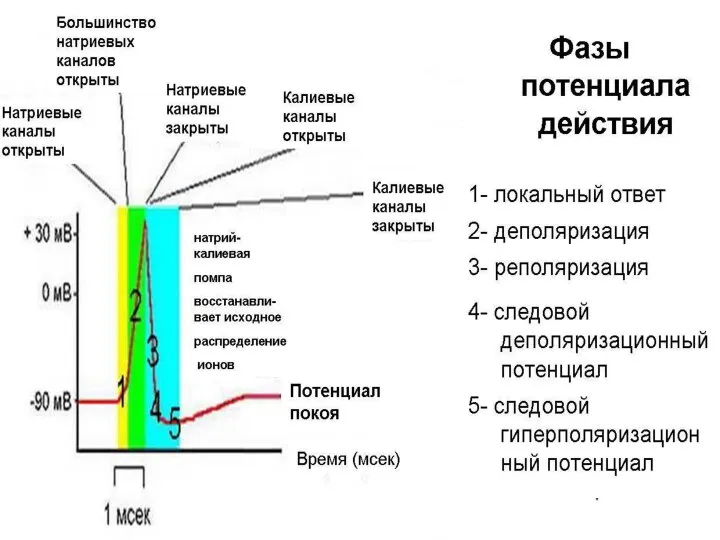
Слайд 18Потенциал действия нейрона
1 - мембранный потенциал покоя, 2 - локальный ответ
Потенциал действия нейрона 1 - мембранный потенциал покоя, 2 - локальный ответ
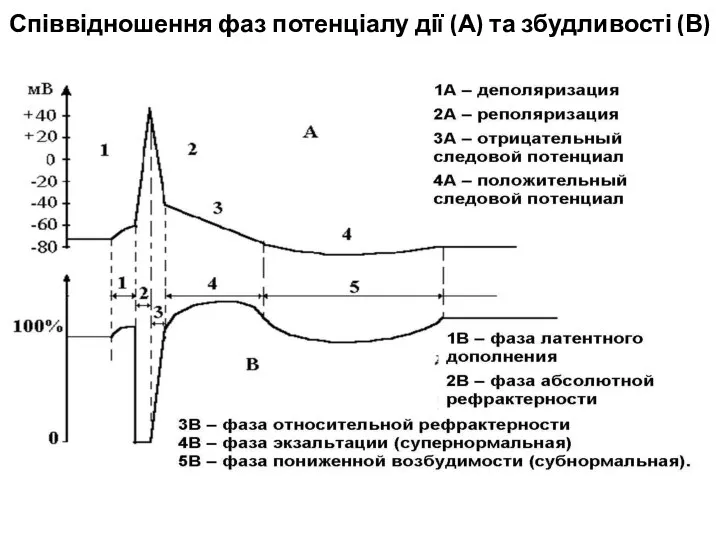
Слайд 20Співвідношення фаз потенціалу дії (А) та збудливості (В)
Співвідношення фаз потенціалу дії (А) та збудливості (В)
Слайд 21Условия и законы проведения возбуждения аксонами
Условия:
1. Анатомическая целостность нервного волокна. Травма, перерезка
Условия и законы проведения возбуждения аксонами
Условия:
1. Анатомическая целостность нервного волокна. Травма, перерезка

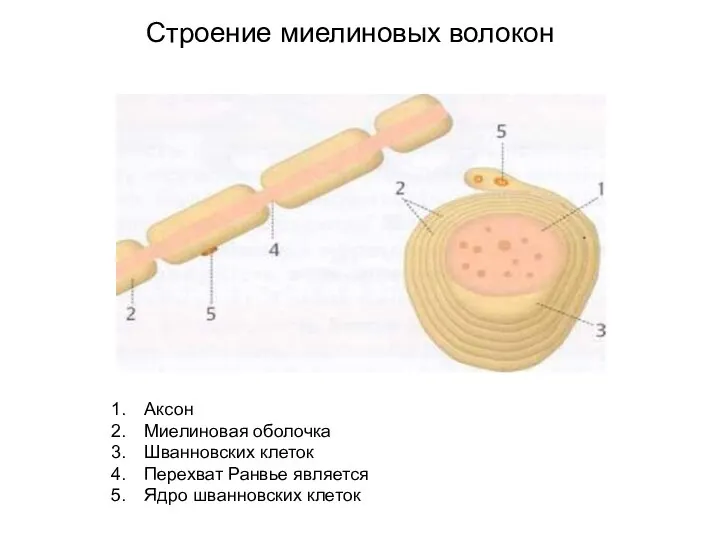
Слайд 22Аксон
Миелиновая оболочка
Шванновских клеток
Перехват Ранвье является
Ядро шванновских клеток
Строение миелиновых волокон
Аксон
Миелиновая оболочка
Шванновских клеток
Перехват Ранвье является
Ядро шванновских клеток
Строение миелиновых волокон
Слайд 23Проведение возбуждения безмиелиновыми и миелиновыми нервными волокнами
Безмиелиновых нервными волокнами возбуждения распространяется непрерывно,
Проведение возбуждения безмиелиновыми и миелиновыми нервными волокнами
Безмиелиновых нервными волокнами возбуждения распространяется непрерывно,


Слайд 25Послідовність процесів скорочення та розслаблення м'язів
Сокращение
Генерация ПД мотонейроном
Высвобождение ацетилхолина в концевой пластинке
Связывание
Послідовність процесів скорочення та розслаблення м'язів
Сокращение
Генерация ПД мотонейроном
Высвобождение ацетилхолина в концевой пластинке
Связывание

Слайд 26Суммация сокращений и тетанус мышцы
В ответ на одинокий импульс происходит сокращение ,
Суммация сокращений и тетанус мышцы
В ответ на одинокий импульс происходит сокращение ,

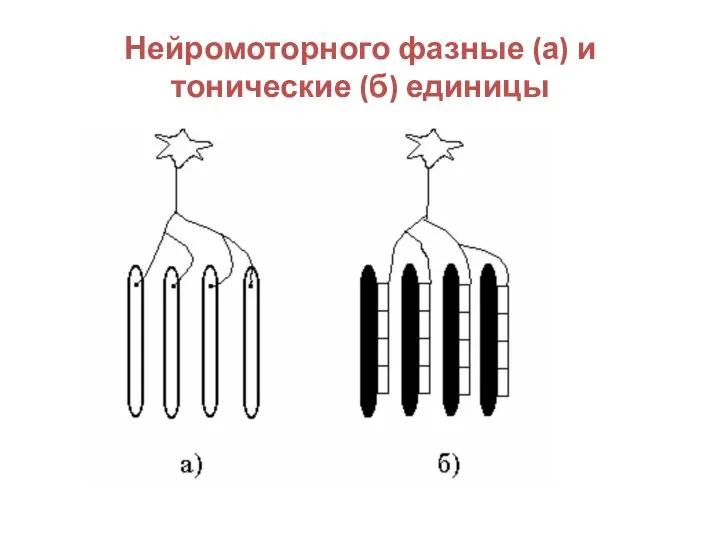
Слайд 27Нейромоторного фазные (а) и тонические (б) единицы
Нейромоторного фазные (а) и тонические (б) единицы
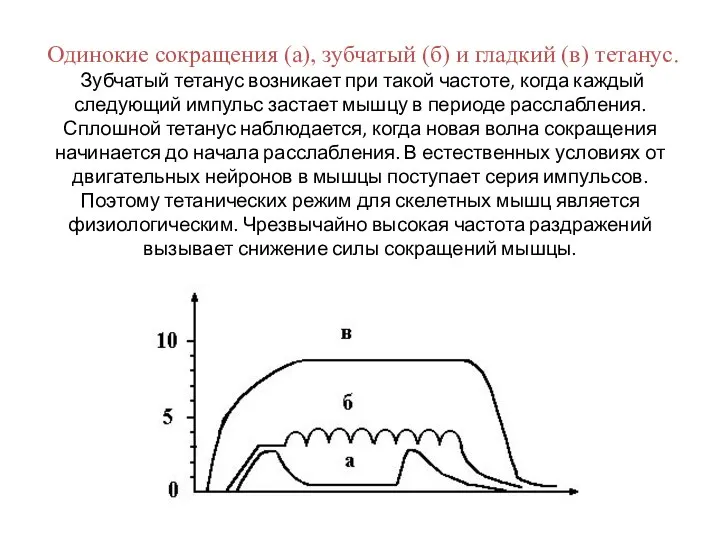
Слайд 28Одинокие сокращения (а), зубчатый (б) и гладкий (в) тетанус.
Зубчатый тетанус возникает
Одинокие сокращения (а), зубчатый (б) и гладкий (в) тетанус. Зубчатый тетанус возникает
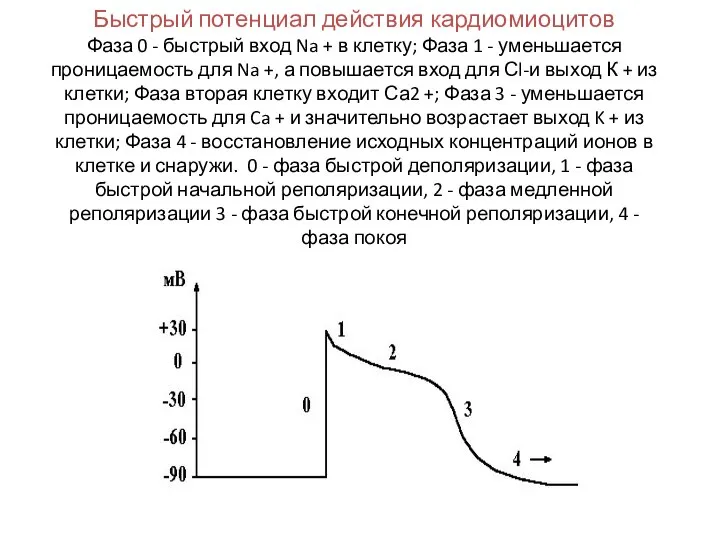
Слайд 29Быстрый потенциал действия кардиомиоцитов
Фаза 0 - быстрый вход Na + в
Быстрый потенциал действия кардиомиоцитов Фаза 0 - быстрый вход Na + в
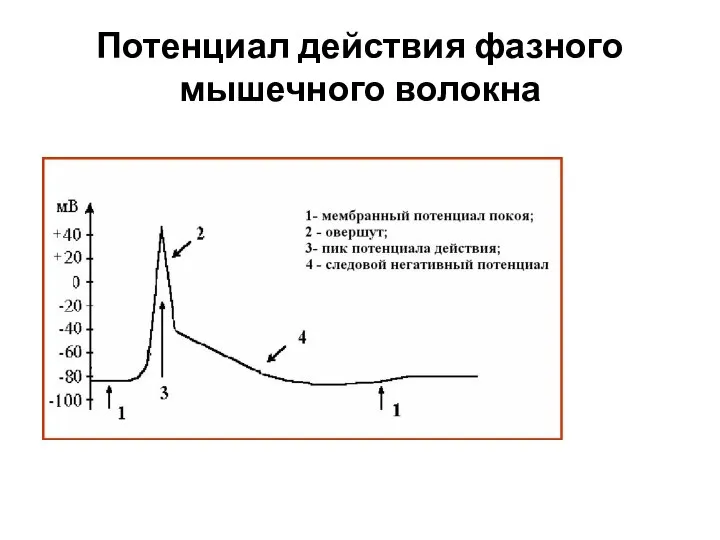
Слайд 30Потенциал действия фазного мышечного волокна
Потенциал действия фазного мышечного волокна
Слайд 31Соотношение между возбуждением и сокращением фазного мышечного волокна
Соотношение между возбуждением и сокращением фазного мышечного волокна
Слайд 32Классификация рецепторов
1. По строению и физиологическими свойствами :
▪ первичные (первичночуствительные);
▪ вторичные (вторичночуствительные).
2.
Классификация рецепторов
1. По строению и физиологическими свойствами :
▪ первичные (первичночуствительные);
▪ вторичные (вторичночуствительные).
2.

Слайд 33Механизм возбуждения первичных и вторичных рецепторов
Развитие рецепторного потенциала (I) в тельце Фатера-Пачини
Механизм возбуждения первичных и вторичных рецепторов
Развитие рецепторного потенциала (I) в тельце Фатера-Пачини
Слайд 34Понятие о рецепторе, рецепторном участке
Главной частью периферического отдела сенсорных систем является рецепторы
Понятие о рецепторе, рецепторном участке
Главной частью периферического отдела сенсорных систем является рецепторы

Слайд 35Функции рецептора : восприятие раздражения , его первичное различение , формирования кода
Функции рецептора : восприятие раздражения , его первичное различение , формирования кода

 Постэмбриональное развитие организмов. Прямое и непрямое развитие
Постэмбриональное развитие организмов. Прямое и непрямое развитие Наследование групп крови
Наследование групп крови Презентация на тему Класс Хрящевые рыбы
Презентация на тему Класс Хрящевые рыбы  Решение задач с биологическим содержанием. История создания клеточной теории
Решение задач с биологическим содержанием. История создания клеточной теории Влияние физических упражнений на кровеносную, дыхательную и нервную системы
Влияние физических упражнений на кровеносную, дыхательную и нервную системы Прощание с ботаникой
Прощание с ботаникой Как заглянуть в митохондрию с помощью наночастиц
Как заглянуть в митохондрию с помощью наночастиц Классические методы селекции
Классические методы селекции Организм многоклеточного животного. Тип Кишечнополостные. 7 класс
Организм многоклеточного животного. Тип Кишечнополостные. 7 класс Презентация на тему Грибы - паразиты 6 класс
Презентация на тему Грибы - паразиты 6 класс  Мини-ландшафт в бутылке или флорариум
Мини-ландшафт в бутылке или флорариум Растения пустыни
Растения пустыни HydraMax. Оптимальная Гидратация
HydraMax. Оптимальная Гидратация Презентация на тему ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ ГЕННОЙ ИНЖЕНЕРИИ
Презентация на тему ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ ГЕННОЙ ИНЖЕНЕРИИ  Презентация на тему ОРГАНЫ ДЫХАНИЯ
Презентация на тему ОРГАНЫ ДЫХАНИЯ  Агатис южный
Агатис южный Ферменты
Ферменты Ткани
Ткани _Как работают в лаборатории
_Как работают в лаборатории Ядовитые растения
Ядовитые растения Глаз - оптическая система!
Глаз - оптическая система! Изменения строения и жизнедеятельности организмов. Основные направления эволюции
Изменения строения и жизнедеятельности организмов. Основные направления эволюции Обобщающий урок по теме: Жизнь растений
Обобщающий урок по теме: Жизнь растений Методы внутривидового типирования микроорганизмов
Методы внутривидового типирования микроорганизмов Биохимия мышц
Биохимия мышц Презентация на тему Вид. Критерии вида
Презентация на тему Вид. Критерии вида  Нарушение обмена витаминов
Нарушение обмена витаминов Современный этап эволюции человека
Современный этап эволюции человека