- Молекулярно-генетический уровень жизни

Содержание
- 2. Регуляция экспрессии гена и степени проявления признака В любой клетке различие между ее фенотипом и генотипом
- 3. Регуляция экспрессии (выражения) генов может осуществляться на нескольких уровнях: генном – изменение количества или локализации генов,
- 4. регуляция осуществляется за счёт увеличения или уменьшения количества копий генов (реже ДНК или хромосом), необходимых или
- 5. Путем: полиплоидизации – кратного увеличения числа хромосом, политенизации хромосом – возникновения многократно реплицированных хромосом, амплификации –
- 6. Прокариоты РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА. ТРАНСКРИПЦИОННЫЙ УРОВЕНЬ
- 7. Действующие в клетках прокариот регуляторные механизмы обеспечивают: Возможность включения (дерепрессии) или выключения (репрессии) экспрессии гена в
- 8. При первом типе регуляции: одни гены в норме репрессированы, и их дерепрессия происходит под влиянием индукторов,
- 9. * У E.coli ферменты, обеспечивающие утилизацию сахаров в качестве единственных источников углерода и азота, синтезируются лишь
- 10. 2. Регуляция второго типа: Обнаружена у фагов, инфицировавших клетки бактерий – обеспечивает запуск «цепной реакции» включения
- 11. Оба типа регуляции осуществляются в отношении лишь тех генов, постоянное функционирование которых нежелательно для клетки, поскольку
- 12. Оперонная регуляция Ф. Жакоб и Ж. Моно 1961 г. Регуляция экспрессии гена. Транскрипционный уровень. Прокариоты
- 13. Оперонная регуляция – транскрипция группы структурных генов, кодирующих полипептиды, тесно связанные между собой функционально, регулируется двумя
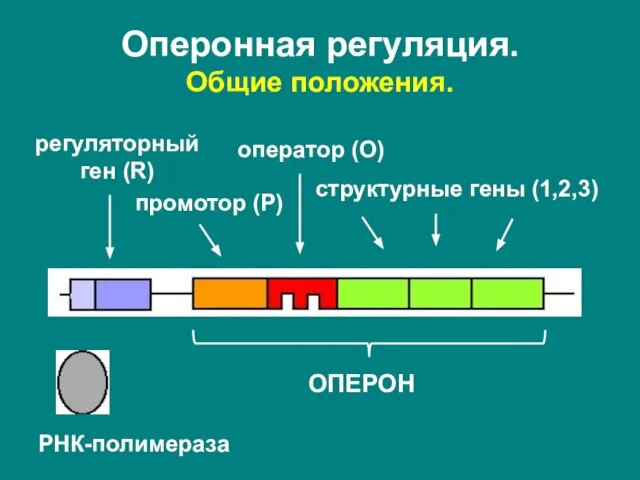
- 14. регуляторный ген (R) ОПЕРОН промотор (P) оператор (O) структурные гены (1,2,3) РНК-полимераза Оперонная регуляция. Общие положения.
- 15. Ген-ингибитор опосредует синтез белка-репрессора, который присоединяется к оператору и блокирует транскрипцию структурных генов, создавая стерические препятствия
- 16. Типы оперонов – в зависимости от влияния: индуцируемые, индуцибельные (катаболизирующие) – включающиеся в работу, регулируют синтез
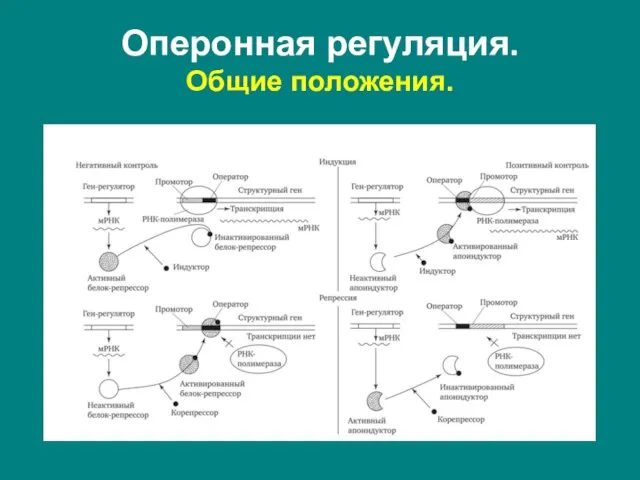
- 17. Типы регуляции оперонов: негативная (регулирующий ген – ингибитор) – эффектор связывается с белком-репрессором, что приводит к
- 18. Оперонная регуляция. Общие положения.
- 19. Lac-оперон – полицистронный оперон кишечной палочки (Escherichia coli), кодирующий гены метаболизма лактозы. Структурные гены оперона: lacZ
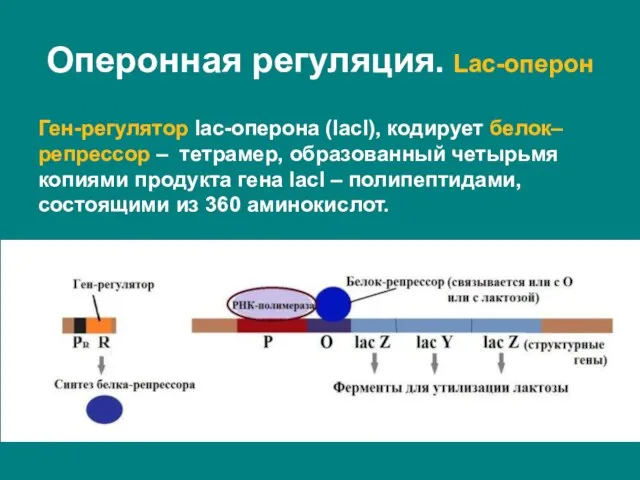
- 20. Ген-регулятор lac-оперона (lacI), кодирует белок–репрессор – тетрамер, образованный четырьмя копиями продукта гена lacI – полипептидами, состоящими
- 21. Основной способ регулирования – это негативная индукция. В безлактозной среде активный белок-репрессор связывается с оператором и
- 22. А Индуктор отсутствует R P O 1 2 3
- 23. Индуктор в среде Б R R P P O O 1 1 2 2 3 3
- 24. В Индуктор исчез из среды R P O 1 2 3
- 25. Дополнительный способ регулирования – позитивная индукция. В среде с низким содержанием глюкозы и наличием лактозы происходит
- 26. Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует превращение АТФ в циклическую
- 27. Оперонная регуляция. Lac-оперон
- 28. Триптофановый оперон – оперон, содержащий гены ферментов, задействованных в биосинтезе аминокислоты триптофан. Был описан в 1953
- 29. Оперонная регуляция. Trp-оперон
- 30. Основной способ регулирования – это негативная репрессия. На значительном расстоянии от оперона находится ген trpR, кодирующий
- 31. Второй способ регулирования – это аттенюация. Сразу после оператора в триптофановом опероне располагается последовательность – лидерная.
- 32. Высокая концентрация триптофана в среде. В начале первого гена оперона закодировано несколько остатков триптофана; в присутствии
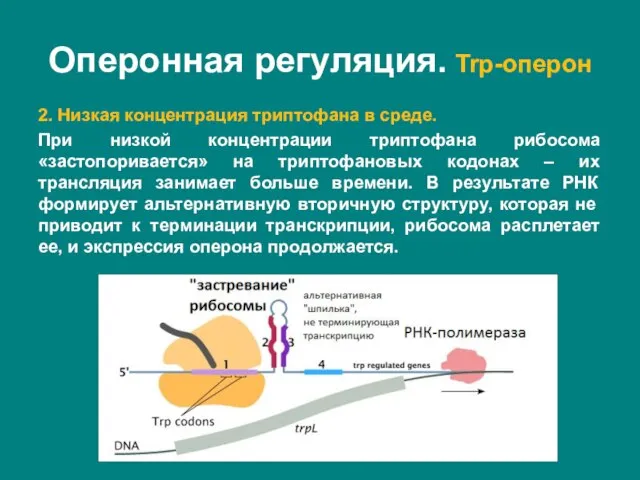
- 33. 2. Низкая концентрация триптофана в среде. При низкой концентрации триптофана рибосома «застопоривается» на триптофановых кодонах –
- 34. Эукариоты РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА.
- 35. Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции. Когда нужно экспрессировать определенные гены, хроматин
- 36. Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется цитозин в последовательности ЦГ,
- 37. Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен только у диплоидных организмов
- 38. Альтернативный сплайсинг. Это регуляция на уровне процессинга. При альтернативном сплайсинге порядок сшивки экзонов может быть различным.
- 39. Регуляция стабильности иРНК. Готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие же иРНК могут дополнительно
- 40. Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов. Регуляция экспрессии гена. Трансляционный
- 42. Скачать презентацию

Слайд 3Регуляция экспрессии (выражения) генов может осуществляться на нескольких уровнях:
генном – изменение
Регуляция экспрессии (выражения) генов может осуществляться на нескольких уровнях:
генном – изменение

Слайд 4регуляция осуществляется за счёт увеличения или уменьшения количества копий генов (реже ДНК
регуляция осуществляется за счёт увеличения или уменьшения количества копий генов (реже ДНК

Слайд 5Путем:
полиплоидизации – кратного увеличения числа хромосом,
политенизации хромосом – возникновения многократно
Путем:
полиплоидизации – кратного увеличения числа хромосом,
политенизации хромосом – возникновения многократно

Слайд 6Прокариоты
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА.
ТРАНСКРИПЦИОННЫЙ УРОВЕНЬ
Прокариоты
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА.
ТРАНСКРИПЦИОННЫЙ УРОВЕНЬ

Слайд 7Действующие в клетках прокариот регуляторные механизмы обеспечивают:
Возможность включения (дерепрессии) или выключения (репрессии)
Действующие в клетках прокариот регуляторные механизмы обеспечивают:
Возможность включения (дерепрессии) или выключения (репрессии)

Слайд 8При первом типе регуляции:
одни гены в норме репрессированы, и их дерепрессия происходит
При первом типе регуляции:
одни гены в норме репрессированы, и их дерепрессия происходит

Слайд 9* У E.coli ферменты, обеспечивающие утилизацию сахаров в качестве единственных источников углерода и азота, синтезируются
* У E.coli ферменты, обеспечивающие утилизацию сахаров в качестве единственных источников углерода и азота, синтезируются

Слайд 102. Регуляция второго типа:
Обнаружена у фагов, инфицировавших клетки бактерий – обеспечивает запуск
2. Регуляция второго типа:
Обнаружена у фагов, инфицировавших клетки бактерий – обеспечивает запуск

Слайд 11Оба типа регуляции осуществляются в отношении лишь тех генов, постоянное функционирование которых
Оба типа регуляции осуществляются в отношении лишь тех генов, постоянное функционирование которых

Слайд 12Оперонная
регуляция
Ф. Жакоб и Ж. Моно
1961 г.
Регуляция экспрессии гена. Транскрипционный уровень. Прокариоты
Оперонная
регуляция
Ф. Жакоб и Ж. Моно
1961 г.
Регуляция экспрессии гена. Транскрипционный уровень. Прокариоты

Слайд 13Оперонная регуляция – транскрипция группы структурных генов, кодирующих полипептиды, тесно связанные между
Оперонная регуляция – транскрипция группы структурных генов, кодирующих полипептиды, тесно связанные между

Слайд 14регуляторный
ген (R)
ОПЕРОН
промотор (P)
оператор (O)
структурные гены (1,2,3)
РНК-полимераза
Оперонная регуляция.
Общие положения.
регуляторный
ген (R)
ОПЕРОН
промотор (P)
оператор (O)
структурные гены (1,2,3)
РНК-полимераза
Оперонная регуляция.
Общие положения.
Слайд 15Ген-ингибитор опосредует синтез белка-репрессора, который присоединяется к оператору и блокирует транскрипцию структурных
Ген-ингибитор опосредует синтез белка-репрессора, который присоединяется к оператору и блокирует транскрипцию структурных

Слайд 16Типы оперонов – в зависимости от влияния:
индуцируемые, индуцибельные (катаболизирующие) – включающиеся в
Типы оперонов – в зависимости от влияния:
индуцируемые, индуцибельные (катаболизирующие) – включающиеся в

Слайд 17Типы регуляции оперонов:
негативная (регулирующий ген – ингибитор) – эффектор связывается с белком-репрессором,
Типы регуляции оперонов:
негативная (регулирующий ген – ингибитор) – эффектор связывается с белком-репрессором,

Слайд 18Оперонная регуляция.
Общие положения.
Оперонная регуляция.
Общие положения.
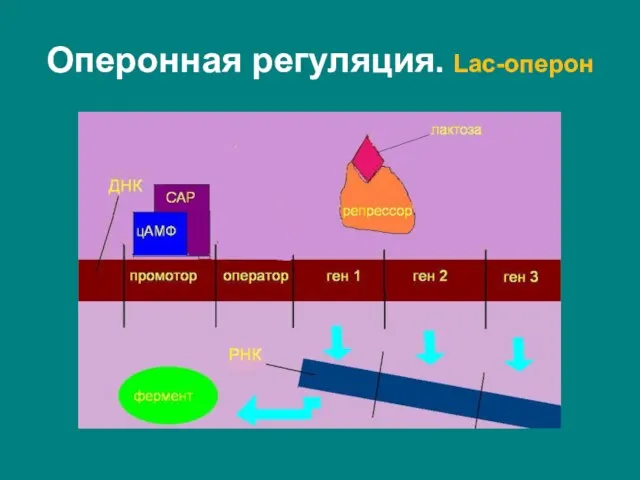
Слайд 19Lac-оперон – полицистронный оперон кишечной палочки (Escherichia coli), кодирующий гены метаболизма лактозы.
Структурные гены оперона:
lacZ
Lac-оперон – полицистронный оперон кишечной палочки (Escherichia coli), кодирующий гены метаболизма лактозы.
Структурные гены оперона:
lacZ

Слайд 20Ген-регулятор lac-оперона (lacI), кодирует белок–репрессор – тетрамер, образованный четырьмя копиями продукта гена
Ген-регулятор lac-оперона (lacI), кодирует белок–репрессор – тетрамер, образованный четырьмя копиями продукта гена
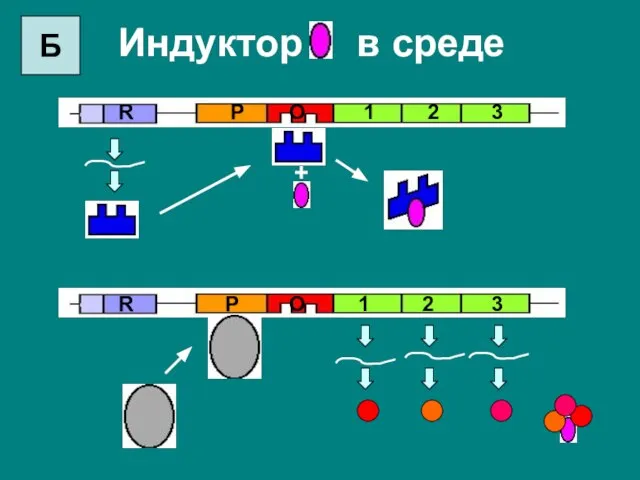
Слайд 21Основной способ регулирования – это негативная индукция.
В безлактозной среде активный белок-репрессор связывается
Основной способ регулирования – это негативная индукция.
В безлактозной среде активный белок-репрессор связывается

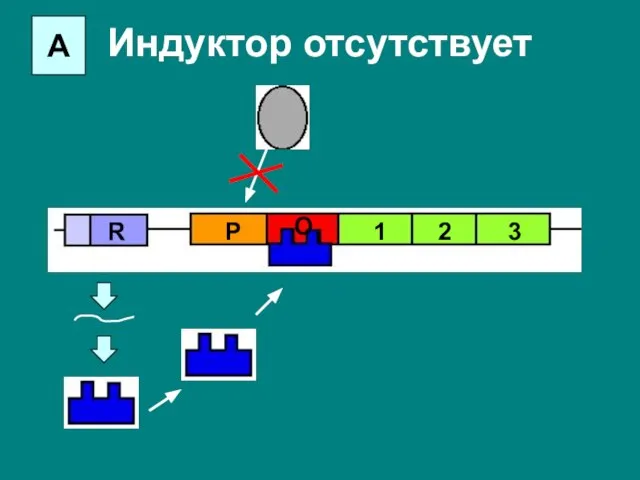
Слайд 22А
Индуктор отсутствует
R
P
O
1
2
3
А
Индуктор отсутствует
R
P
O
1
2
3
Слайд 23Индуктор в среде
Б
R
R
P
P
O
O
1
1
2
2
3
3
+
Индуктор в среде
Б
R
R
P
P
O
O
1
1
2
2
3
3
+
Слайд 24В
Индуктор исчез из среды
R
P
O
1
2
3
В
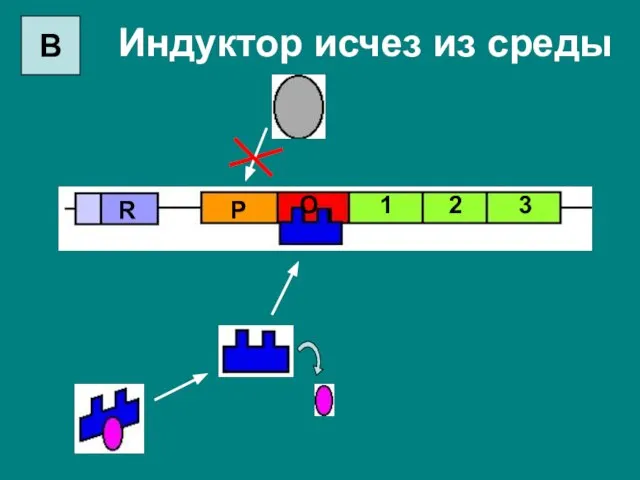
Индуктор исчез из среды
R
P
O
1
2
3
Слайд 25Дополнительный способ регулирования – позитивная индукция.
В среде с низким содержанием глюкозы и
Дополнительный способ регулирования – позитивная индукция.
В среде с низким содержанием глюкозы и

Слайд 26Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует
Когда концентрация глюкозы в клетке снижается, происходит активация фермента аденилатциклазы, которая катализирует

Слайд 27Оперонная регуляция. Lac-оперон
Оперонная регуляция. Lac-оперон
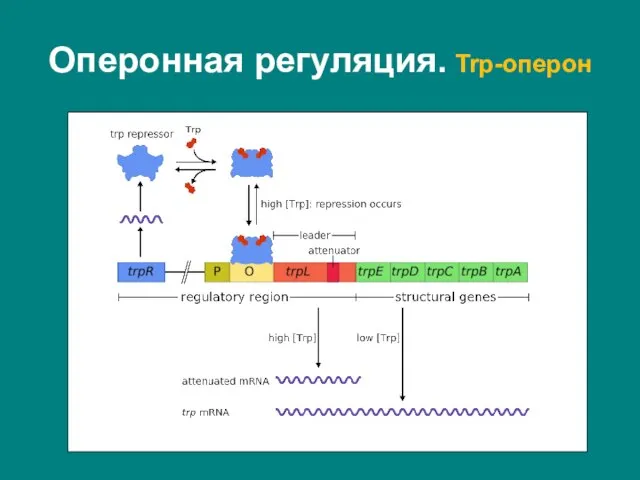
Слайд 28Триптофановый оперон – оперон, содержащий гены ферментов, задействованных в биосинтезе аминокислоты триптофан.
Был
Триптофановый оперон – оперон, содержащий гены ферментов, задействованных в биосинтезе аминокислоты триптофан.
Был

Слайд 29Оперонная регуляция. Trp-оперон
Оперонная регуляция. Trp-оперон
Слайд 30Основной способ регулирования – это негативная репрессия.
На значительном расстоянии от оперона находится
Основной способ регулирования – это негативная репрессия.
На значительном расстоянии от оперона находится

Слайд 31Второй способ регулирования – это аттенюация.
Сразу после оператора в триптофановом опероне
Второй способ регулирования – это аттенюация.
Сразу после оператора в триптофановом опероне

Слайд 32Высокая концентрация триптофана в среде.
В начале первого гена оперона закодировано несколько остатков
Высокая концентрация триптофана в среде.
В начале первого гена оперона закодировано несколько остатков
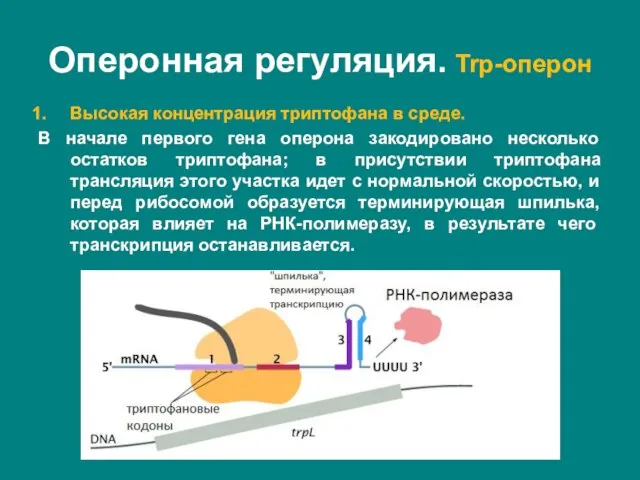
Слайд 332. Низкая концентрация триптофана в среде.
При низкой концентрации триптофана рибосома «застопоривается» на
2. Низкая концентрация триптофана в среде.
При низкой концентрации триптофана рибосома «застопоривается» на
Слайд 34Эукариоты
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА.
Эукариоты
РЕГУЛЯЦИЯ ЭКСПРЕССИИ ГЕНА И СТЕПЕНИ ПРОЯВЛЕНИЯ ПРИЗНАКА.
Слайд 35Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции. Когда нужно
Конденсация и деконденсация хроматина. Это наиболее универсальный метод регуляции транскрипции. Когда нужно

Слайд 36Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется
Метилирование и деметилирование ДНК. Метилирование ДНК происходит в регуляторных областях гена. Метилируется

Слайд 37Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен
Геномный импринтинг. Это малоизученный способ регуляции экспрессии генов у эукариот. Он возможен

Слайд 38Альтернативный сплайсинг. Это регуляция на уровне процессинга. При альтернативном сплайсинге порядок сшивки
Альтернативный сплайсинг. Это регуляция на уровне процессинга. При альтернативном сплайсинге порядок сшивки

Слайд 39Регуляция стабильности иРНК. Готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие
Регуляция стабильности иРНК. Готовые иРНК не «допускаются» к рибосомам или разрушаются. Другие

Слайд 40Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов.
Регуляция
Риборегуляторы. Были обнаружены РНК, выполняющие регуляторные функции путем ослабления работы отдельных генов.
Регуляция

 Последствия деятельности человека в экосистемах
Последствия деятельности человека в экосистемах Порушення під час вагітності. Шкідливі звички і вагітність
Порушення під час вагітності. Шкідливі звички і вагітність Знакомство с внешним строением растения
Знакомство с внешним строением растения Отряды насекомых
Отряды насекомых Генетический анализ
Генетический анализ Дикие животные
Дикие животные Великаны и гиганты. Определение деревьев по побегам
Великаны и гиганты. Определение деревьев по побегам Среды обитания организмов. Водная среда обитания
Среды обитания организмов. Водная среда обитания 5 класс увеличительные приборы
5 класс увеличительные приборы Мухомор красный
Мухомор красный Размножение организмов
Размножение организмов Грибное лукошко
Грибное лукошко Рыбы, гады и насекомые
Рыбы, гады и насекомые Инструменты для груминга
Инструменты для груминга Отделы и области тела животных
Отделы и области тела животных Теория внешнего вмешательства в происхождение человека
Теория внешнего вмешательства в происхождение человека Клетка и её органоиды
Клетка и её органоиды Эукариотическая клетка. Цитоплазма (2)
Эукариотическая клетка. Цитоплазма (2) Отряд рукокрылые
Отряд рукокрылые Гаметогенез и дробление
Гаметогенез и дробление Презентация на тему Влияние человека на животных
Презентация на тему Влияние человека на животных  Филин обыкновенный
Филин обыкновенный ООО Биота Россия. Биологические пестициды
ООО Биота Россия. Биологические пестициды Внутренние органы птиц
Внутренние органы птиц Коровы
Коровы Биоразнообразие. Повторение. 7 класс
Биоразнообразие. Повторение. 7 класс Биосфера, как область взаимодействия общества и природы
Биосфера, как область взаимодействия общества и природы Гистофизиология скелетной мышечной ткани
Гистофизиология скелетной мышечной ткани