- Строение прокариотической клетки

Содержание
- 3. Структуры бактериальной клетки Постоянные Временные Клеточная стенка (?) ЦПМ, мезосомы Нуклеоид Цитоплазма с рибосомами Капсула Пили
- 4. Функции клеточной стенки: защищает бактерии от механических и других повреждающих факторов; определяет форму; участвует в обмене
- 10. Клеточная стенка грамотрицательных бактерий
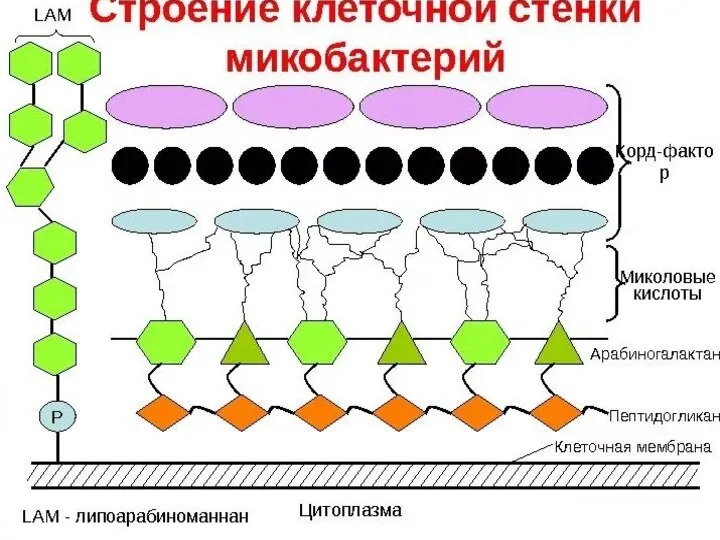
- 11. Клеточная стенка микобактерий состоит из нескольких слоев, часть из которых содержит сахара и характеризуется относительно постоянным
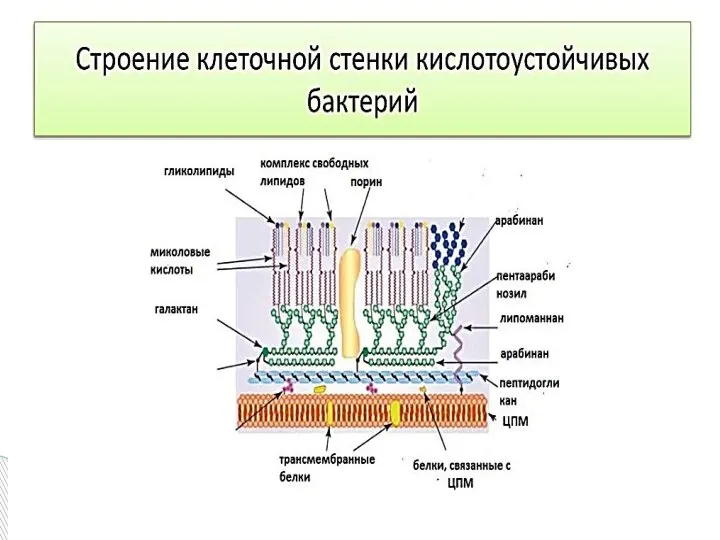
- 12. Слой гликолипидов называют " микозидами " и иногда сравнивают с микрокапсулой. Микозиды структурно и функционально напоминают
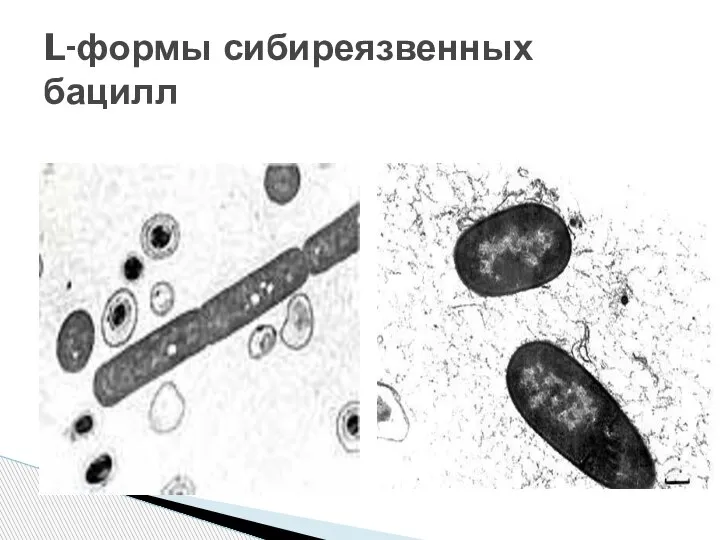
- 19. L-формы сибиреязвенных бацилл
- 20. L-формы листерий
- 22. Периплазматическое пространство - это обособленный компартмент клеток грамотрицательных бактерий. Представляет собой объём, заключённый между плазматической и
- 23. Периплазма представляет собой гелеобразный матрикс, состоящий из белков и особых олигосахаридов, которые называют осморегулируемые периплазматические глюканы.
- 26. ЦПМ бактериальной клетки имеет типичное трехслойное строение; ее основу составляет двойной слой фосфолипидов, покрытый с двух
- 27. Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ. Имеют форму клубков, петель и пузырьков (везикулярное
- 29. Клетки фототрофных бактерий содержат внутрицитоплазматические мембраны, увеличивающие общую площадь мембраны и содержащие фотосинтетический аппарат. Они имеют
- 30. Схемы строения мембран фототрофных бактерий
- 32. Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий. Часть цитоплазмы, которая имеет гомогенную коллоидную консистенцию и содержит
- 33. Запасные гранулы содержат временный избыток метаболитов. Наличие и количество гранул изменяется в зависимости от вида бактерий
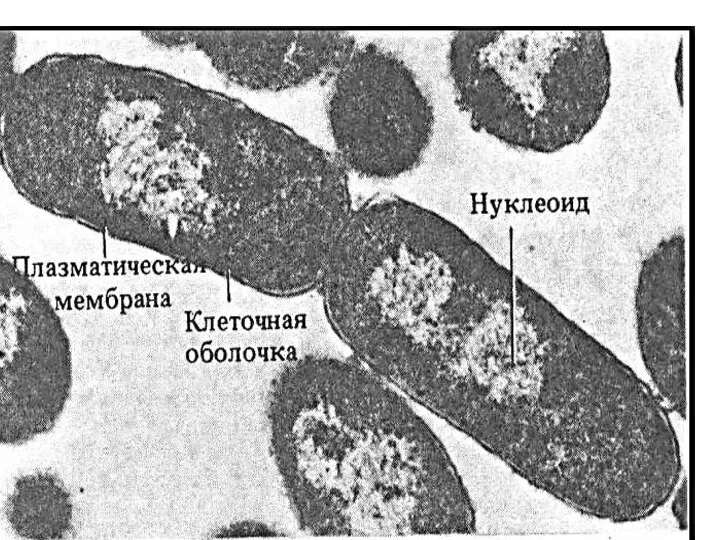
- 34. Нуклеоид – эквивалент ядра («подобный ядру»), содержит одну молекулу ДНК, представляющую собой двойную спираль, замкнутую в
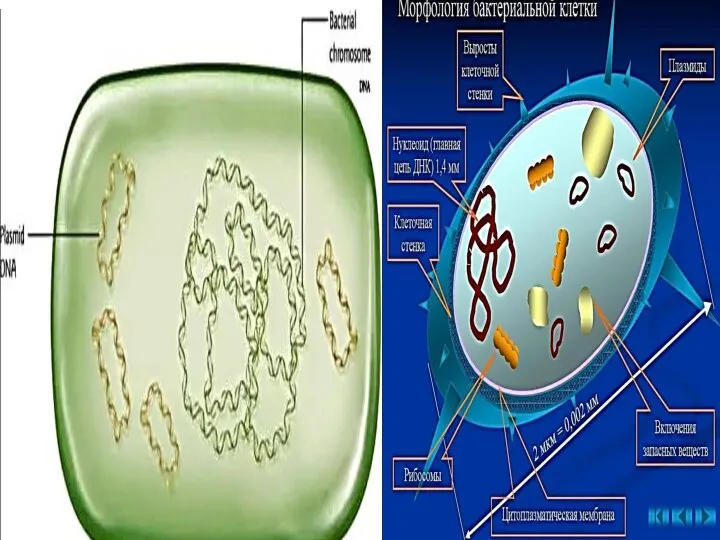
- 36. Плазмиды Плазмиды — внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой ДНК (термин «плазмиды»
- 37. Некоторые плазмиды находятся под строгим контролем. Это означает, что их репликация сопряжена с репликацией хромосомы так,
- 42. Рибосомы – рибонуклеопротеидные частицы, участвующие в биосинтезе белка. Размер составляет 20 нм; имеют округлую или слегка
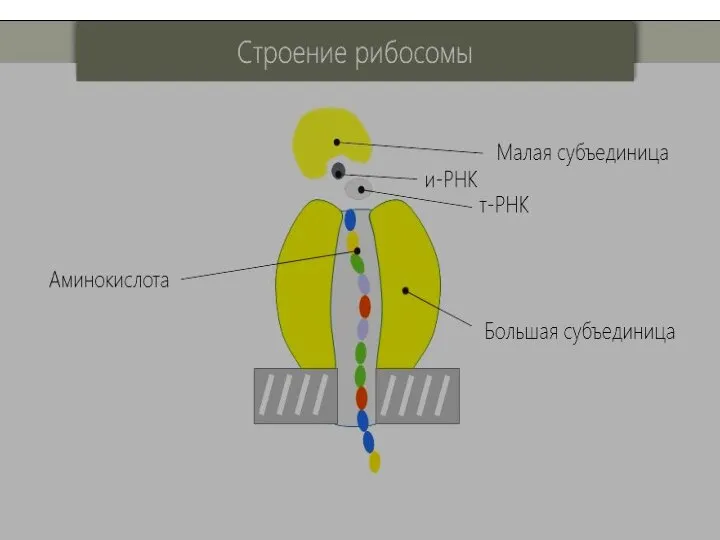
- 43. Рибосомы- это макромолекулярный комплекс, который состоит из двух субчастиц разной величины: малая субчастица (30 S); большая
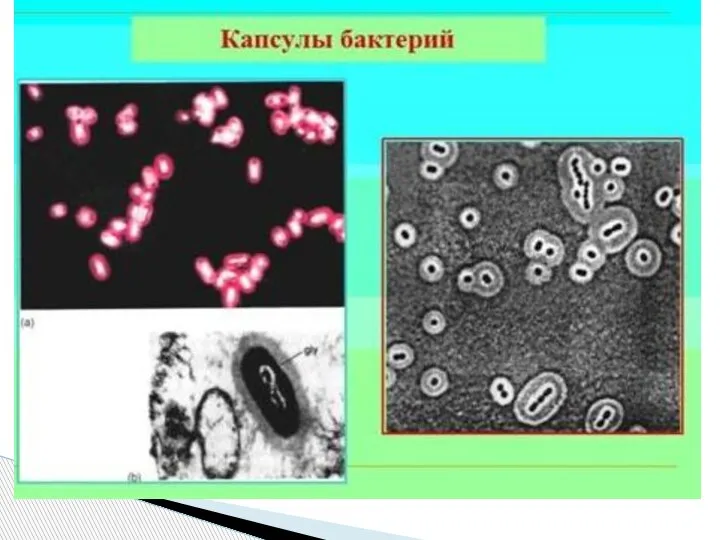
- 45. Капсула - слизистая структура толщиной более 0,2 мкм (макрокапсула) и менее 0,2 мкм (микрокапсула, выявляемая лишь
- 46. Функции капсулы различаются у непатогенных и патогенных бактерий. У непатогенных бактерий капсула чаще всего служит: -
- 47. У бактерий, вызывающих заболевания животных и человека, капсула является важным фактором вирулентности, так как она защищает
- 48. Иногда капсула мимикрирует под молекулярные структуры организма хозяина. Например, капсульный антиген К1 кишечной палочки (Escherichia coli)
- 52. От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких границ. Слизистый слой – во
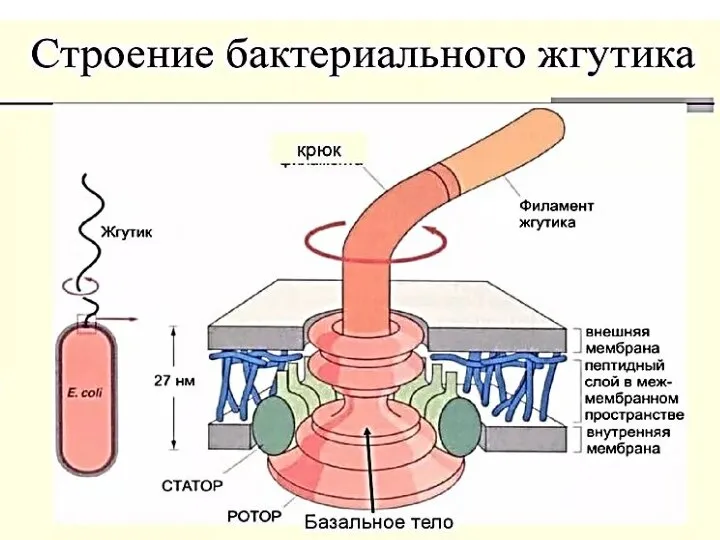
- 53. Жгутики бактерий определяют подвижность бактериальной клетки и являются Н-антигеном. Жгутики представляют собой тонкие нити, берущие начало
- 56. Базальное тело (трансмембранный мотор). Представляет собой систему колец, находящихся в плазматической мембране и клеточной стенке бактерий.
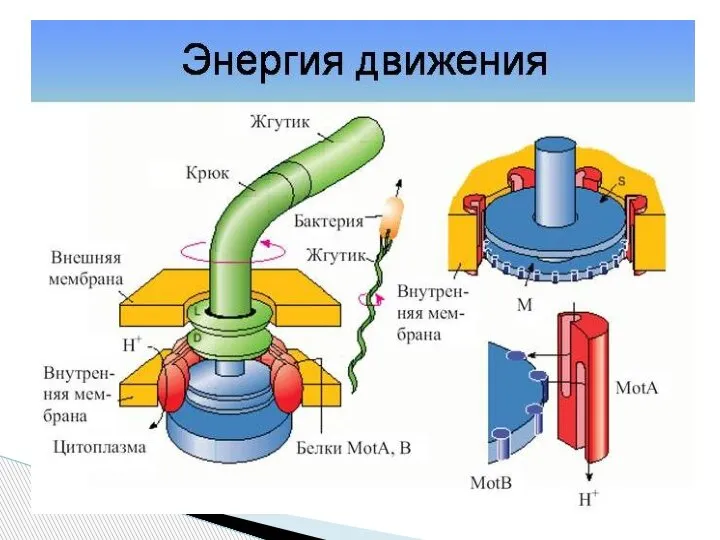
- 58. Точный механизм работы базального тела неизвестен. Большинство исследователей полагает, что базальное тело преобразует химическую энергию в
- 60. Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно со скоростью 1/3 от
- 61. Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против часовой стрелки) в один
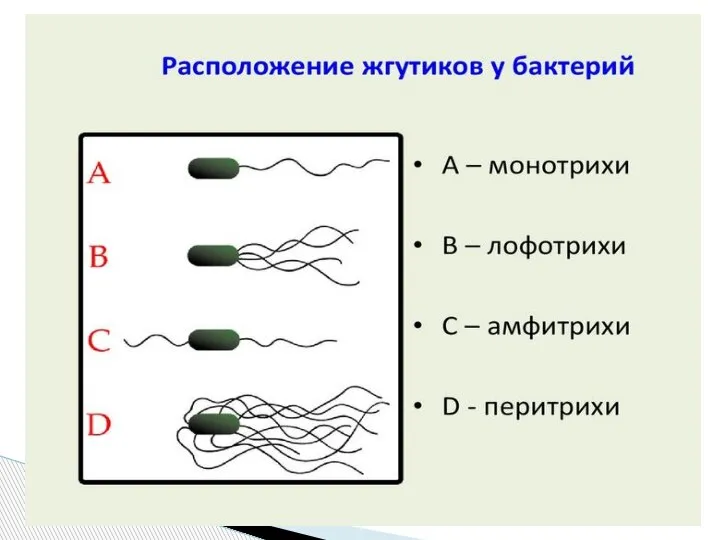
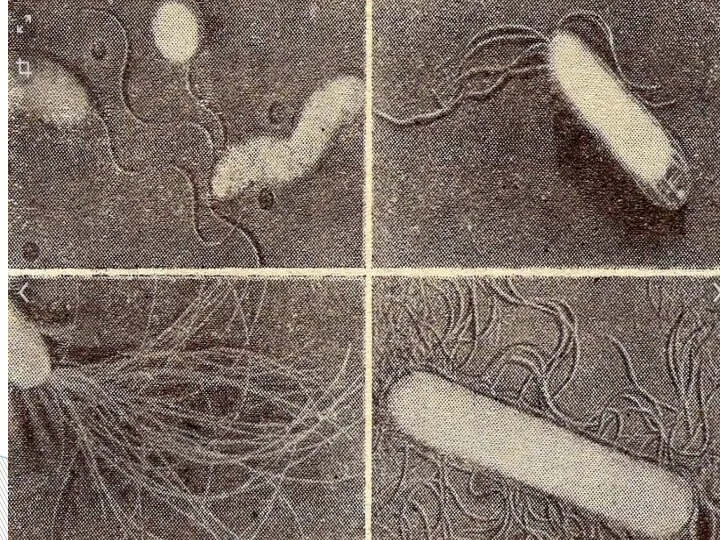
- 64. Характер движения определяется характером жгутования. Бактерии с полярно расположенными жгутиками движутся по прямой, только иногда делая
- 65. Таксисы - двигательные реакции свободно передвигающихся микроорганизмов. Таксисы происходят под влиянием: одностороннего раздражения, вызванного действием света
- 66. По характеру реагирования на раздражение различают: - положительный таксис — движения по направлению к раздражителю, -
- 67. Скользящее движение — движение отдельных бактериальных клеток или их колоний по твёрдой поверхности вдоль их длинной
- 68. Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем
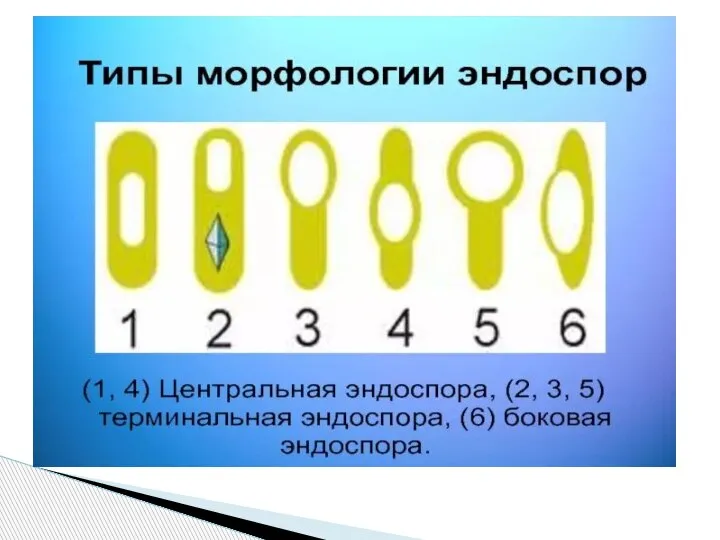
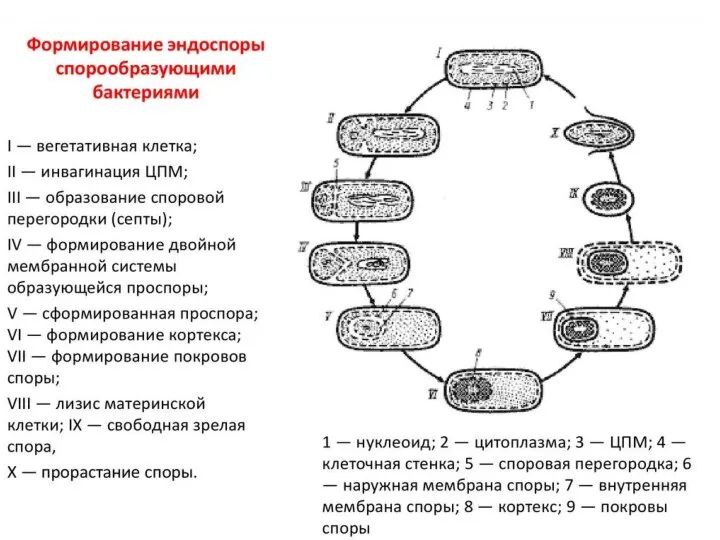
- 69. Эндоспоры – особый тип покоящихся клеток фирмакутных, т.е. бактерий с грамположительным типом оболочки, хотя некоторые образующие
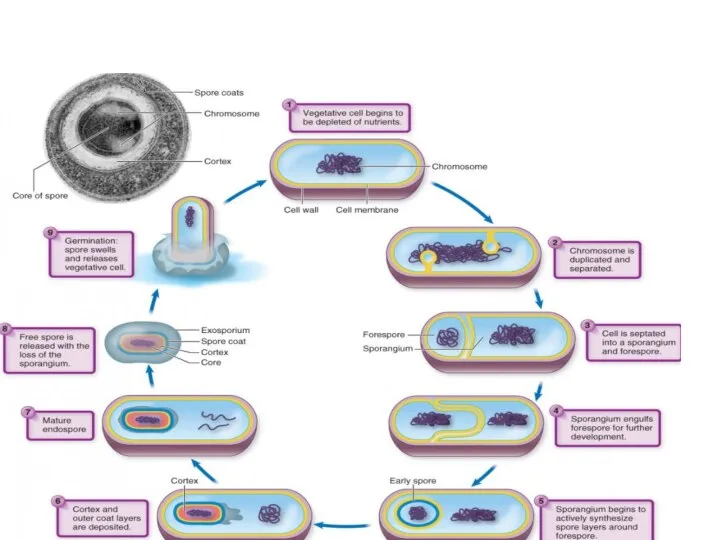
- 71. Процесс спорообразования принято подразделять на 6 последовательных стадий, смена которых определяется активированием соответствующих генов. При этом
- 72. Стадия 2 характеризуется отделением от осевого тяжа ДНК участка, соответствующего хромосоме, который движется к полюсу клетки.
- 73. Между 2-й и 3-й стадиями происходит врастание двойной мембраны или септы так, что в результате образуется
- 74. На 4-й стадии между двумя мембранами проспоры начинается образование муреинового слоя. Процесс идет в 2этапа: -
- 75. На 5-й стадии спорообразования у бацилл начинают формироваться покровы споры. На срезах они сначала имеют вид
- 76. На 6-й стадии спорообразования заканчивается формирование всех характерных структур споры, она приобретает термоустойчивость. Иногда выделяют еще
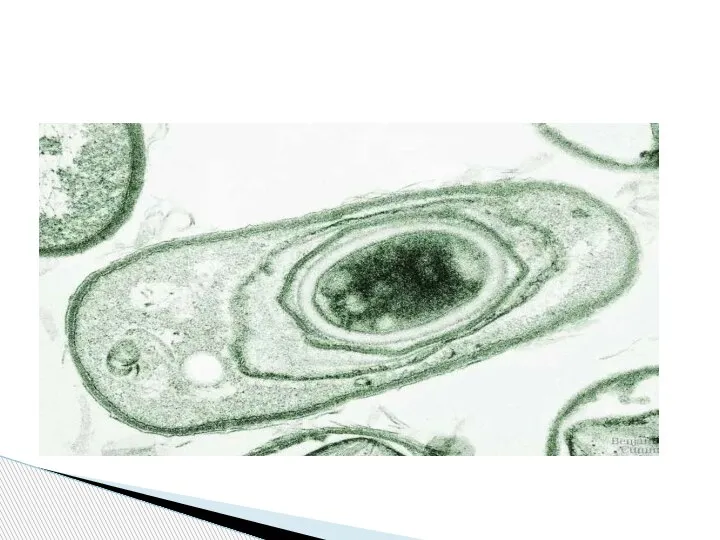
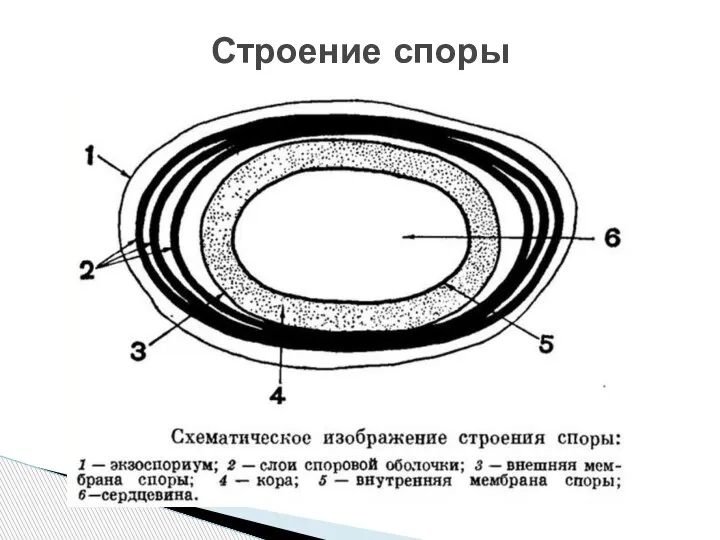
- 77. Строение зрелой споры у разных видов соответствует одной и той же схеме. Сердцевина споры содержит белки
- 78. Кортекс и зародышевая стенка кроме муреина содержат ряд литических ферментов, которые активируются при прорастании или при
- 79. Экзоспориум образует как бы свободный мешок, внутри которого лежит спора, контактируя с ним только в некоторых
- 80. У многих видов рода Clostridium споры снабжены различными придатками и выростами. По классификации, предложенной В.И. Дудой,
- 83. Строение споры
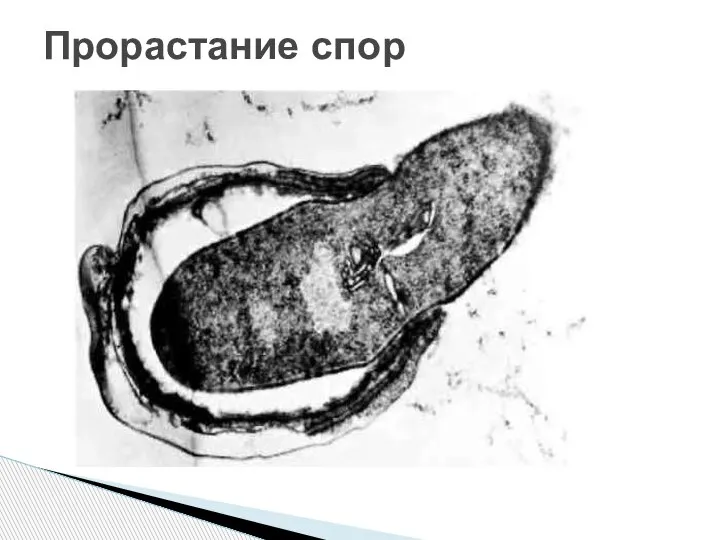
- 85. Процесс прорастания споры принято подразделять на три стадии – активацию, инициацию и вырастание. Активация означает готовность
- 86. Прорастание спор
- 93. Скачать презентацию
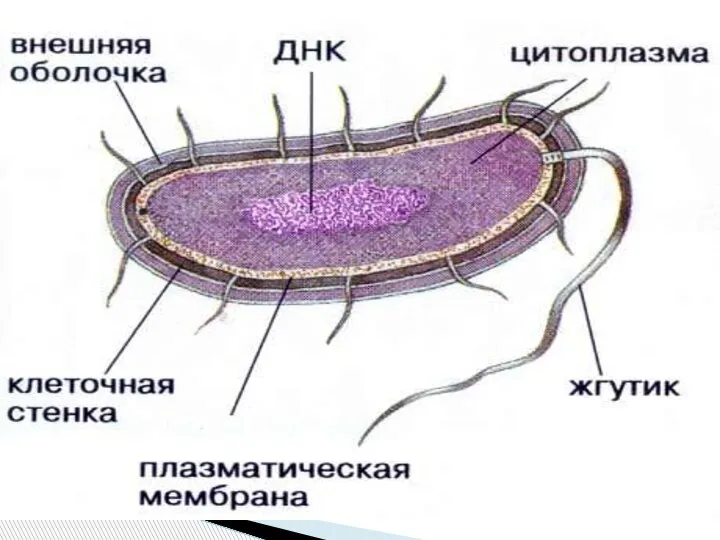
Слайд 3Структуры бактериальной клетки
Постоянные
Временные
Клеточная
стенка (?)
ЦПМ,
мезосомы
Нуклеоид
Цитоплазма
с рибосомами
Капсула
Пили
Эндоспора
Жгутики
Плазмиды
Структуры бактериальной клетки
Постоянные
Временные
Клеточная
стенка (?)
ЦПМ,
мезосомы
Нуклеоид
Цитоплазма
с рибосомами
Капсула
Пили
Эндоспора
Жгутики
Плазмиды
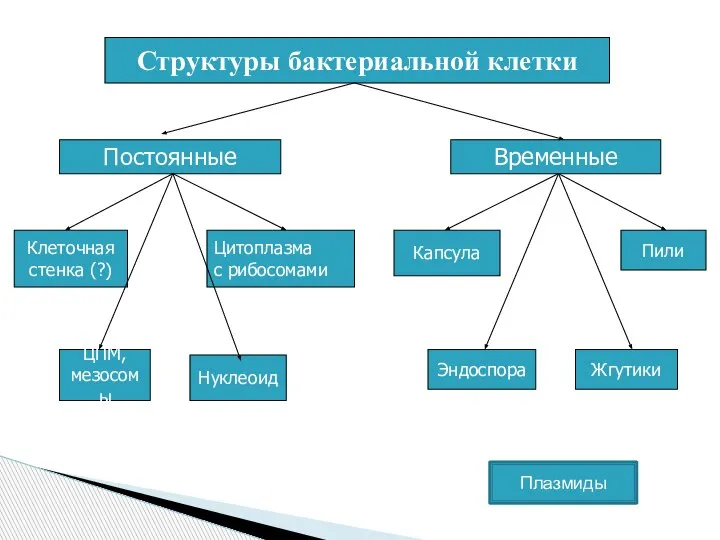
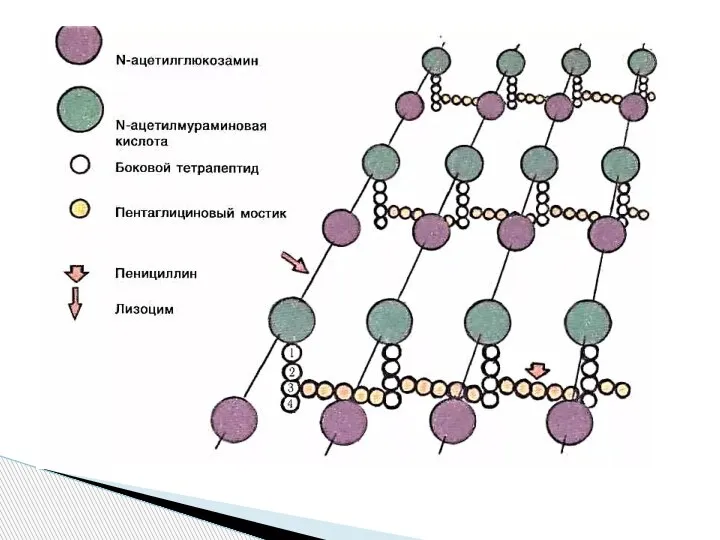
Слайд 4Функции клеточной стенки:
защищает бактерии от механических и других повреждающих факторов;
определяет форму;
участвует в
Функции клеточной стенки:
защищает бактерии от механических и других повреждающих факторов;
определяет форму;
участвует в

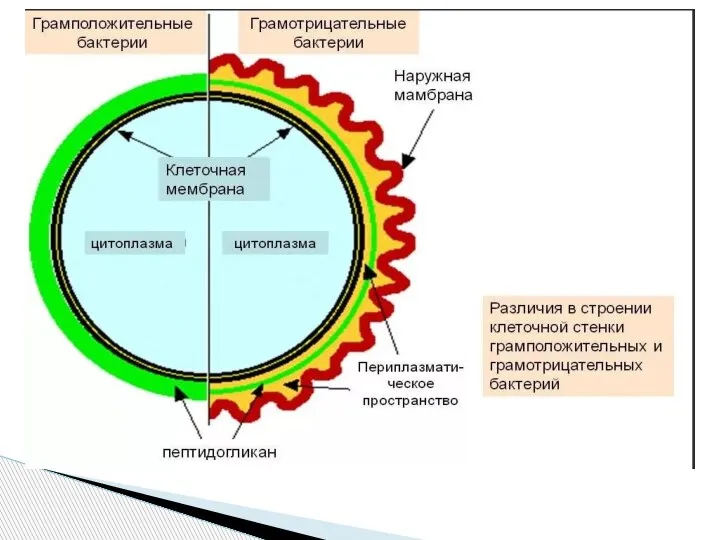

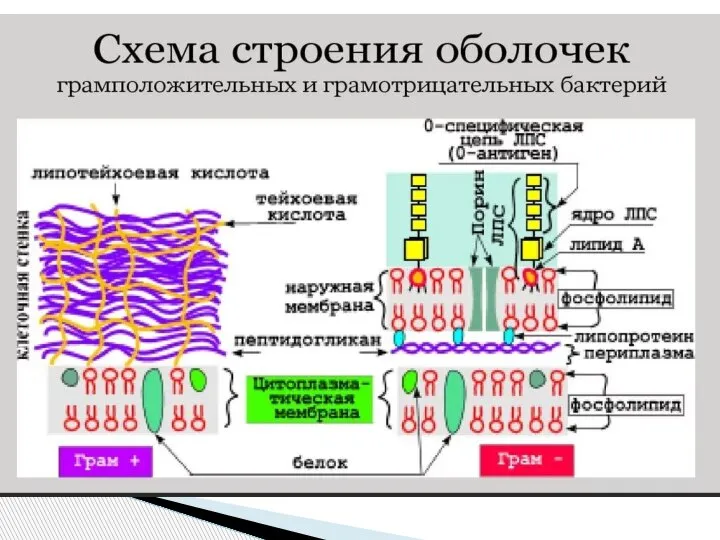
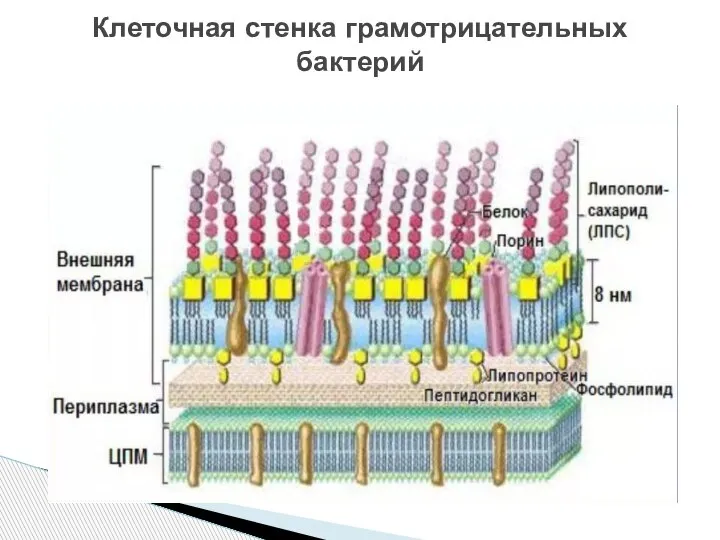
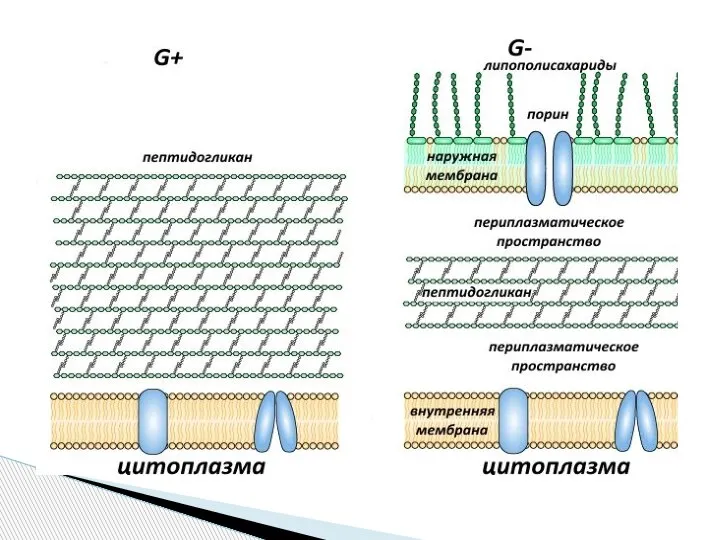
Слайд 10Клеточная стенка грамотрицательных бактерий
Клеточная стенка грамотрицательных бактерий
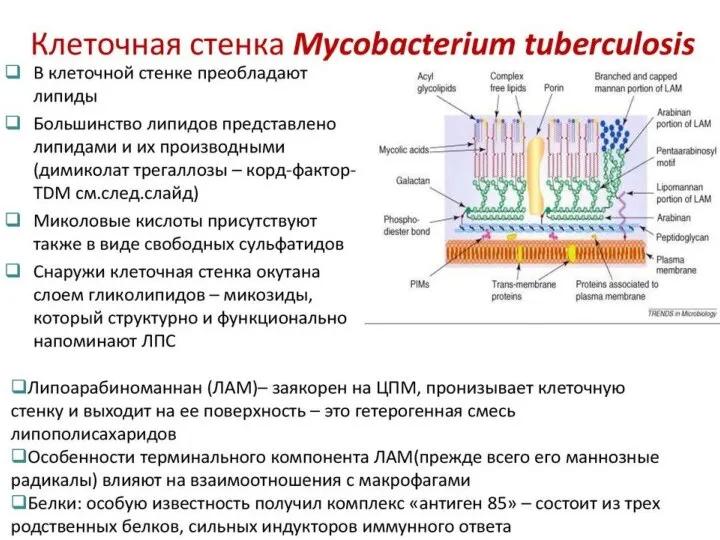
Слайд 11Клеточная стенка микобактерий состоит из нескольких слоев, часть из которых содержит сахара
Клеточная стенка микобактерий состоит из нескольких слоев, часть из которых содержит сахара

Слайд 12Слой гликолипидов называют " микозидами " и иногда сравнивают с микрокапсулой. Микозиды
Слой гликолипидов называют " микозидами " и иногда сравнивают с микрокапсулой. Микозиды

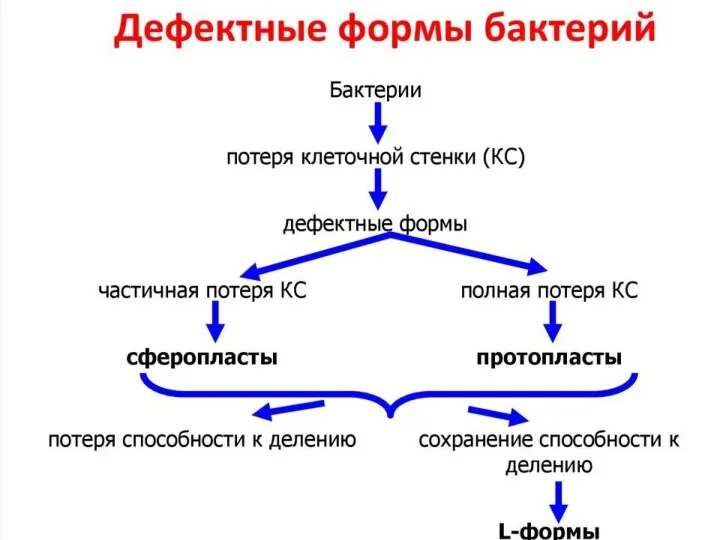


Слайд 19L-формы сибиреязвенных бацилл
L-формы сибиреязвенных бацилл
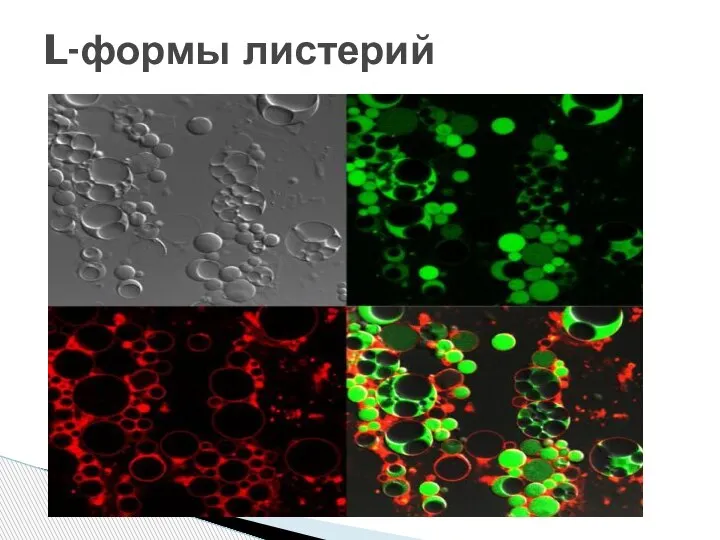
Слайд 20L-формы листерий
L-формы листерий

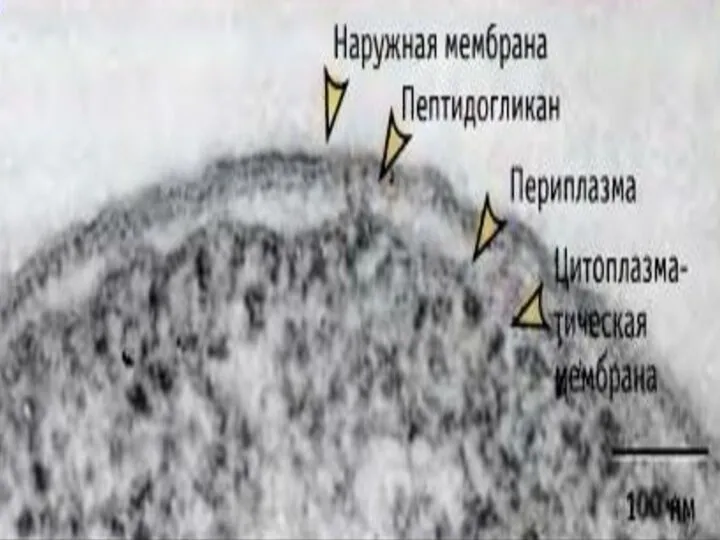
Слайд 22Периплазматическое пространство - это обособленный компартмент клеток грамотрицательных бактерий.
Представляет собой объём, заключённый
Периплазматическое пространство - это обособленный компартмент клеток грамотрицательных бактерий.
Представляет собой объём, заключённый

Слайд 23Периплазма представляет собой гелеобразный матрикс, состоящий из белков и особых олигосахаридов, которые
Периплазма представляет собой гелеобразный матрикс, состоящий из белков и особых олигосахаридов, которые

Слайд 26ЦПМ бактериальной клетки имеет типичное трехслойное строение; ее основу составляет двойной слой
ЦПМ бактериальной клетки имеет типичное трехслойное строение; ее основу составляет двойной слой

Слайд 27Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ.
Имеют форму клубков,
Мезосомы представляют собой мембранные структуры, образуемые при закручивании ЦПМ.
Имеют форму клубков,


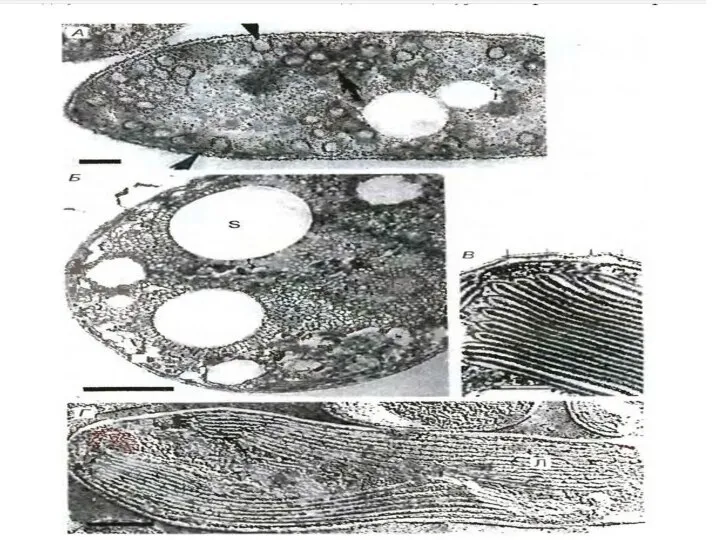
Слайд 29Клетки фототрофных бактерий содержат внутрицитоплазматические мембраны, увеличивающие общую площадь мембраны и содержащие
Клетки фототрофных бактерий содержат внутрицитоплазматические мембраны, увеличивающие общую площадь мембраны и содержащие

Слайд 30Схемы строения мембран фототрофных бактерий
Схемы строения мембран фототрофных бактерий

Слайд 32Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий.
Часть цитоплазмы, которая имеет гомогенную
Содержимое клетки, окруженное ЦПМ, составляет цитоплазму бактерий.
Часть цитоплазмы, которая имеет гомогенную

Слайд 33Запасные гранулы содержат временный избыток метаболитов.
Наличие и количество гранул изменяется в
Запасные гранулы содержат временный избыток метаболитов.
Наличие и количество гранул изменяется в

Слайд 34Нуклеоид – эквивалент ядра («подобный ядру»), содержит одну молекулу ДНК, представляющую собой
Нуклеоид – эквивалент ядра («подобный ядру»), содержит одну молекулу ДНК, представляющую собой

Слайд 36Плазмиды
Плазмиды — внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой
Плазмиды
Плазмиды — внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой

Слайд 37Некоторые плазмиды находятся под строгим контролем. Это означает, что их репликация сопряжена
Некоторые плазмиды находятся под строгим контролем. Это означает, что их репликация сопряжена




Слайд 42Рибосомы – рибонуклеопротеидные частицы, участвующие в биосинтезе белка.
Размер составляет 20 нм; имеют
Рибосомы – рибонуклеопротеидные частицы, участвующие в биосинтезе белка.
Размер составляет 20 нм; имеют

Слайд 43Рибосомы- это макромолекулярный комплекс, который состоит из двух субчастиц разной величины:
малая субчастица
Рибосомы- это макромолекулярный комплекс, который состоит из двух субчастиц разной величины:
малая субчастица

Слайд 45Капсула - слизистая структура толщиной более 0,2 мкм (макрокапсула) и менее 0,2
Капсула - слизистая структура толщиной более 0,2 мкм (макрокапсула) и менее 0,2

Слайд 46Функции капсулы различаются у непатогенных и патогенных бактерий.
У непатогенных бактерий капсула чаще
Функции капсулы различаются у непатогенных и патогенных бактерий.
У непатогенных бактерий капсула чаще

Слайд 47У бактерий, вызывающих заболевания животных и человека, капсула является важным фактором вирулентности,
У бактерий, вызывающих заболевания животных и человека, капсула является важным фактором вирулентности,

Слайд 48Иногда капсула мимикрирует под молекулярные структуры организма хозяина. Например, капсульный антиген К1
Иногда капсула мимикрирует под молекулярные структуры организма хозяина. Например, капсульный антиген К1

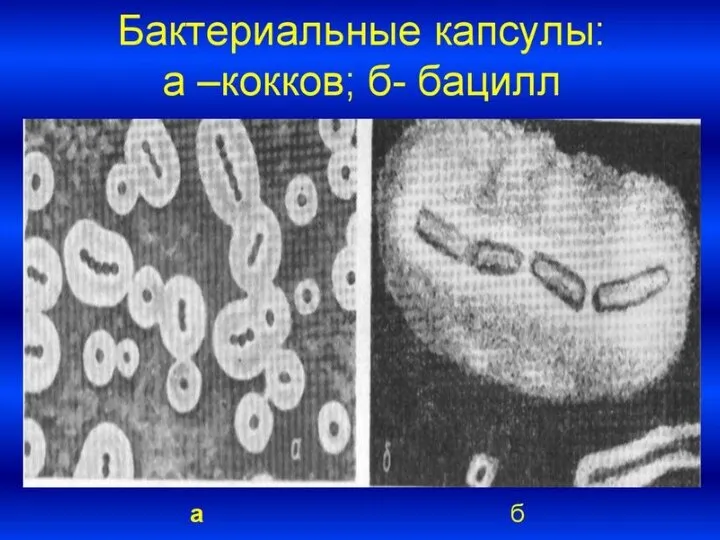
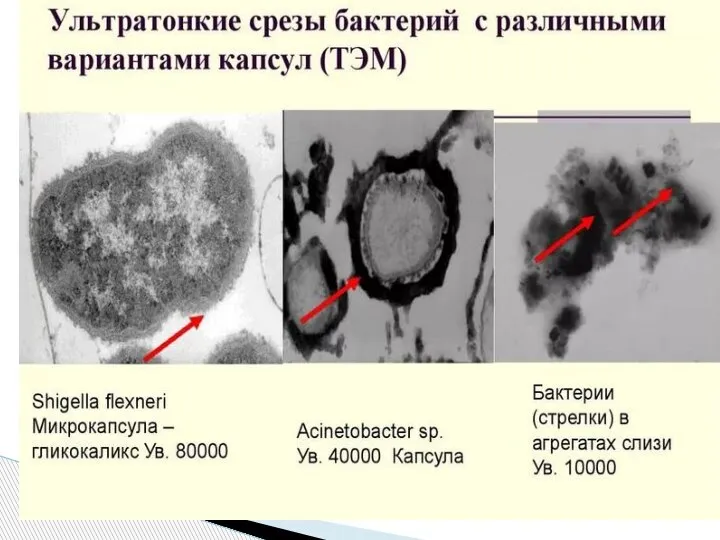
Слайд 52От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких границ.
От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких границ.

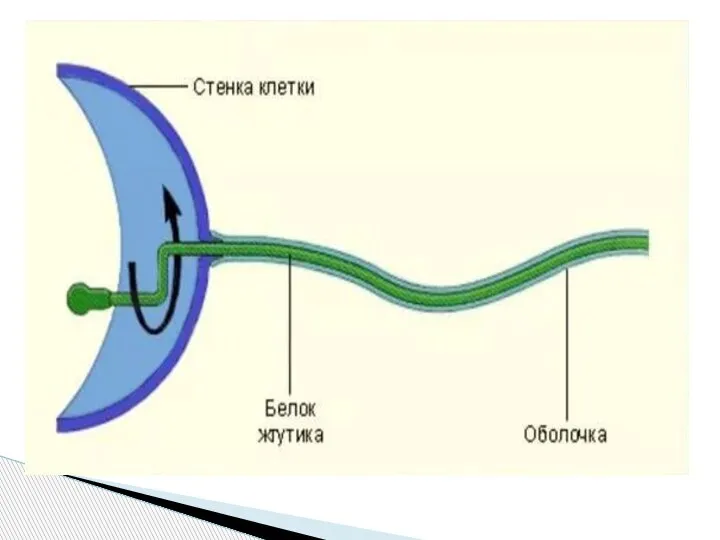
Слайд 53Жгутики бактерий определяют подвижность бактериальной клетки и являются Н-антигеном.
Жгутики представляют собой
Жгутики бактерий определяют подвижность бактериальной клетки и являются Н-антигеном.
Жгутики представляют собой

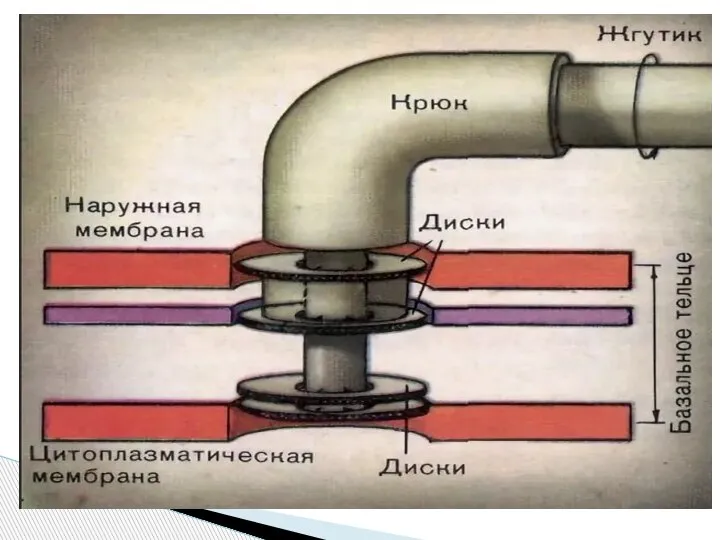
Слайд 56Базальное тело (трансмембранный мотор). Представляет собой систему колец, находящихся в плазматической мембране
Базальное тело (трансмембранный мотор). Представляет собой систему колец, находящихся в плазматической мембране

Слайд 58Точный механизм работы базального тела неизвестен.
Большинство исследователей полагает, что базальное тело
Точный механизм работы базального тела неизвестен.
Большинство исследователей полагает, что базальное тело

Слайд 60Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно
Вращение мотора вызывает пассивное вращение филамента. Более массивная клетка начинает вращаться примерно

Слайд 61Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против
Бактерии-перитрихи собирают при движении все свои жгутики (каждый из которых вращается против

Слайд 64Характер движения определяется характером жгутования.
Бактерии с полярно расположенными жгутиками движутся по
Характер движения определяется характером жгутования.
Бактерии с полярно расположенными жгутиками движутся по

Слайд 65Таксисы - двигательные реакции свободно передвигающихся микроорганизмов. Таксисы происходят под влиянием:
одностороннего раздражения,
Таксисы - двигательные реакции свободно передвигающихся микроорганизмов. Таксисы происходят под влиянием:
одностороннего раздражения,

Слайд 66По характеру реагирования на раздражение различают:
- положительный таксис — движения по
По характеру реагирования на раздражение различают:
- положительный таксис — движения по

Слайд 67Скользящее движение — движение отдельных бактериальных клеток или их колоний по твёрдой
Скользящее движение — движение отдельных бактериальных клеток или их колоний по твёрдой

Слайд 68Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х

Слайд 69Эндоспоры – особый тип покоящихся клеток фирмакутных, т.е. бактерий с грамположительным типом
Эндоспоры – особый тип покоящихся клеток фирмакутных, т.е. бактерий с грамположительным типом

Слайд 71Процесс спорообразования принято подразделять на 6 последовательных стадий, смена которых определяется активированием
Процесс спорообразования принято подразделять на 6 последовательных стадий, смена которых определяется активированием

Слайд 72Стадия 2 характеризуется отделением от осевого тяжа ДНК участка, соответствующего хромосоме, который
Стадия 2 характеризуется отделением от осевого тяжа ДНК участка, соответствующего хромосоме, который

Слайд 73Между 2-й и 3-й стадиями происходит врастание двойной мембраны или септы так,

Слайд 74На 4-й стадии между двумя мембранами проспоры начинается образование муреинового слоя. Процесс
На 4-й стадии между двумя мембранами проспоры начинается образование муреинового слоя. Процесс

Слайд 75На 5-й стадии спорообразования у бацилл начинают формироваться покровы споры. На срезах
На 5-й стадии спорообразования у бацилл начинают формироваться покровы споры. На срезах

Слайд 76На 6-й стадии спорообразования заканчивается формирование всех характерных структур споры, она приобретает
На 6-й стадии спорообразования заканчивается формирование всех характерных структур споры, она приобретает

Слайд 77Строение зрелой споры у разных видов соответствует одной и той же схеме.
Строение зрелой споры у разных видов соответствует одной и той же схеме.

Слайд 78Кортекс и зародышевая стенка кроме муреина содержат ряд литических ферментов, которые активируются
Кортекс и зародышевая стенка кроме муреина содержат ряд литических ферментов, которые активируются

Слайд 79Экзоспориум образует как бы свободный мешок, внутри которого лежит спора, контактируя с
Экзоспориум образует как бы свободный мешок, внутри которого лежит спора, контактируя с

Слайд 80У многих видов рода Clostridium споры снабжены различными придатками и выростами. По
У многих видов рода Clostridium споры снабжены различными придатками и выростами. По

Слайд 83Строение споры
Строение споры
Слайд 85Процесс прорастания споры принято подразделять на три стадии – активацию, инициацию и
Процесс прорастания споры принято подразделять на три стадии – активацию, инициацию и

Слайд 86Прорастание спор
Прорастание спор





 Змея. Виды змей
Змея. Виды змей Формы иммуного ответа
Формы иммуного ответа Siberian Tiger
Siberian Tiger Современное состояние ушастой совы (Asio Otus), гнездящейся в урбанизированном ландшафте
Современное состояние ушастой совы (Asio Otus), гнездящейся в урбанизированном ландшафте Цепи питания
Цепи питания Хозяйственно-биологические особенности лошадей
Хозяйственно-биологические особенности лошадей Экология на садовом участке. Подготовка рассады
Экология на садовом участке. Подготовка рассады Видообразование. Борьба за существование и естественный отбор – движущие силы эволюции
Видообразование. Борьба за существование и естественный отбор – движущие силы эволюции Видообразование. Результаты микроэволюции
Видообразование. Результаты микроэволюции Полив комнатных растений
Полив комнатных растений Коралловые полипы
Коралловые полипы Биологические закономерности в русских сказках
Биологические закономерности в русских сказках Парамфистомоз. Дикроцелиоз. Эуритрематоз
Парамфистомоз. Дикроцелиоз. Эуритрематоз Естественный отбор. Основоположники современной (синтетической) теории эволюции
Естественный отбор. Основоположники современной (синтетической) теории эволюции Класс Листостебельные мхи
Класс Листостебельные мхи День моржа
День моржа Complex analysis of metabolic status, intracellular pH, viscosity and cytoskeleton of human
Complex analysis of metabolic status, intracellular pH, viscosity and cytoskeleton of human Приготовление микропрепарата
Приготовление микропрепарата Презентация на тему Птичьи разговоры
Презентация на тему Птичьи разговоры  Многообразие живого и наука систематика
Многообразие живого и наука систематика транскрипция и трансляция
транскрипция и трансляция Исторические концепции происхождения жизни
Исторические концепции происхождения жизни Хвойные растения
Хвойные растения Вегетативное размножение растений
Вегетативное размножение растений Углеводы. Важнейшие моносахариды
Углеводы. Важнейшие моносахариды Предпосылки возникновения учения Ч. Дарвина
Предпосылки возникновения учения Ч. Дарвина Виды РНК. тРНК
Виды РНК. тРНК Кто такие домашние животные?
Кто такие домашние животные?