- Patogennost_i_virulentnost_mikroorganizmov

Содержание
- 2. Патогенность (болезнетворность) (от греч. pathos - страдание, genos - происхождение) - видовой полидетерминантный признак возбудителя, характеризующий
- 3. Патогенность характеризуют следующие признаки Потенциальность может реализоваться при определенных условиях: наличии восприимчивого макроорганизма; определенном влиянии факторов
- 4. 3. Полидетерминантность контролируется совокупностью хромосомных генов и мобильных генетических элементов (плазмид, транспозонов, умеренных фагов). Острова патогенности
- 5. 4) могут утрачиваться и передаваться другим микроорганизмам; 5) способны к горизонтальной внутиривидовой и межвидовой передаче, поэтому
- 6. 4. Специфичность - способность вызывать типичные для данного вида возбудителя патоморфологические и патофизиологические изменения в определенных
- 7. 6. Качественная категория. Классификация микроорганизмов по степени патогенности: 1) облигатно-патогенные способны преодолевать защитные механизмы макроорганизма и
- 8. 3) непатогенные - сапрофиты (греч. sapros - гнилой, phytos - растение) питаются органическими веществами от отмерших
- 9. Вирулентность - биологическое свойство микроорганизма, характеризующее меру патогенности конкретного штамма патогенного возбудителя (степень фенотипического проявления патогенности
- 10. Характеристика вирулентности 1. Индивидуальный (штаммовый) признак микроорганизма, количественное выражение патогенности. 2. Вариабельность. Штаммы вида по этому
- 11. 3. Динамичность определяется совокупностью факторов патогенности; подвержена фенотипическому и генотипическому изменению; возможно естественное и искусственное снижение
- 12. Количественный критерий вирулентности - минимальное количество микроорганизмов, которое при определенном способе заражения животных определенного вида, веса
- 13. Показатели, характеризующие степень вирулентности: а) Dcl (Dosis certe letalis) - наименьшая доза микробных клеток, являющаяся смертельной
- 14. Каждому патогенному виду микроорганизмов присущ свойственный только ему набор конкретных материальных субстратов - факторов патогенности. В
- 15. Классификации экзотоксинов I. По молекулярной организации экзотоксины делятся на 2 группы: 1. Состоящие из двух фрагментов:
- 16. II. По степени связи с бактериальной клеткой экзотоксины делятся на 3 класса: 1. Секретируемые во внешнюю
- 17. III. По механизму действия бактериальные токсины делятся на 6 групп: 1. Мембранотоксины, повреждающие клеточные мембраны: вызывают
- 18. 3. Активирующие пути метаболизма, контролируемые вторичными мессенджерами: нарушают синтез различных клеточных белков, не вызывая непосредственной гибели
- 19. 5. Эксфолиатины S. aureus, влияющие на процесс взаимодействия клеток между собой и с межклеточными веществами, вызывают
- 20. Эндотоксины 1. Липополисахаридные комплексы КС, главным образом Грам–бактерий, освобождаются только после гибели бактерий. Существенной частью эндотоксина
- 22. Скачать презентацию
Слайд 2Патогенность (болезнетворность) (от греч. pathos - страдание, genos - происхождение) - видовой
Патогенность (болезнетворность) (от греч. pathos - страдание, genos - происхождение) - видовой

Слайд 3Патогенность характеризуют следующие признаки
Потенциальность может реализоваться при определенных условиях: наличии восприимчивого макроорганизма;
Патогенность характеризуют следующие признаки
Потенциальность может реализоваться при определенных условиях: наличии восприимчивого макроорганизма;

Слайд 43. Полидетерминантность контролируется совокупностью хромосомных генов и мобильных генетических элементов (плазмид, транспозонов,
3. Полидетерминантность контролируется совокупностью хромосомных генов и мобильных генетических элементов (плазмид, транспозонов,

Слайд 54) могут утрачиваться и передаваться другим микроорганизмам;
5) способны к горизонтальной внутиривидовой
4) могут утрачиваться и передаваться другим микроорганизмам;
5) способны к горизонтальной внутиривидовой

Слайд 64. Специфичность - способность вызывать типичные для данного вида возбудителя патоморфологические и
4. Специфичность - способность вызывать типичные для данного вида возбудителя патоморфологические и

Слайд 76. Качественная категория.
Классификация микроорганизмов по степени патогенности:
1) облигатно-патогенные способны преодолевать
6. Качественная категория.
Классификация микроорганизмов по степени патогенности:
1) облигатно-патогенные способны преодолевать

Слайд 83) непатогенные - сапрофиты (греч. sapros - гнилой, phytos - растение) питаются
3) непатогенные - сапрофиты (греч. sapros - гнилой, phytos - растение) питаются

Слайд 9Вирулентность - биологическое свойство микроорганизма, характеризующее меру патогенности конкретного штамма патогенного возбудителя
Вирулентность - биологическое свойство микроорганизма, характеризующее меру патогенности конкретного штамма патогенного возбудителя

Слайд 10Характеристика вирулентности
1. Индивидуальный (штаммовый) признак микроорганизма, количественное выражение патогенности.
2. Вариабельность.
Штаммы
Характеристика вирулентности
1. Индивидуальный (штаммовый) признак микроорганизма, количественное выражение патогенности.
2. Вариабельность.
Штаммы

Слайд 113. Динамичность определяется совокупностью факторов патогенности; подвержена фенотипическому и генотипическому изменению; возможно
3. Динамичность определяется совокупностью факторов патогенности; подвержена фенотипическому и генотипическому изменению; возможно

Слайд 12Количественный критерий вирулентности - минимальное количество микроорганизмов, которое при определенном способе заражения

Слайд 13Показатели, характеризующие степень вирулентности:
а) Dcl (Dosis certe letalis) - наименьшая доза
Показатели, характеризующие степень вирулентности:
а) Dcl (Dosis certe letalis) - наименьшая доза

Слайд 14Каждому патогенному виду микроорганизмов присущ свойственный только ему набор конкретных материальных субстратов
Каждому патогенному виду микроорганизмов присущ свойственный только ему набор конкретных материальных субстратов

Слайд 15Классификации экзотоксинов
I. По молекулярной организации экзотоксины делятся на 2 группы:
1. Состоящие
Классификации экзотоксинов
I. По молекулярной организации экзотоксины делятся на 2 группы:
1. Состоящие

Слайд 16II. По степени связи с бактериальной клеткой экзотоксины делятся на 3 класса:
II. По степени связи с бактериальной клеткой экзотоксины делятся на 3 класса:

Слайд 17III. По механизму действия бактериальные токсины делятся на 6 групп:
1. Мембранотоксины,
III. По механизму действия бактериальные токсины делятся на 6 групп:
1. Мембранотоксины,

Слайд 183. Активирующие пути метаболизма, контролируемые вторичными мессенджерами: нарушают синтез различных клеточных белков,
3. Активирующие пути метаболизма, контролируемые вторичными мессенджерами: нарушают синтез различных клеточных белков,

Слайд 195. Эксфолиатины S. aureus, влияющие на процесс взаимодействия клеток между собой и
5. Эксфолиатины S. aureus, влияющие на процесс взаимодействия клеток между собой и

Слайд 20Эндотоксины
1. Липополисахаридные комплексы КС, главным образом Грам–бактерий, освобождаются только после гибели бактерий.
Эндотоксины
1. Липополисахаридные комплексы КС, главным образом Грам–бактерий, освобождаются только после гибели бактерий.

 Болезни, передаваемые половым путем. Меры профилактики
Болезни, передаваемые половым путем. Меры профилактики Гранулематоз Вегенера - системный васкулит
Гранулематоз Вегенера - системный васкулит Всероссийская олимпиада по детской хирургии. Викторина
Всероссийская олимпиада по детской хирургии. Викторина Флоссы. Зубные нити
Флоссы. Зубные нити Gnano: ощути вкус нанотехнологий. - презентация_
Gnano: ощути вкус нанотехнологий. - презентация_ vvedene_v_pediatriyu
vvedene_v_pediatriyu Патология сердечно-сосудистой системы у детей
Патология сердечно-сосудистой системы у детей Патогенные микобактерии
Патогенные микобактерии Мы здоровью скажем ДА
Мы здоровью скажем ДА ჰემორაგიული ცხელება
ჰემორაგიული ცხელება Адаптация и стресс
Адаптация и стресс Эпилепсия
Эпилепсия Здоров'я
Здоров'я Коленный сустав
Коленный сустав Респираторно-синцитиальный вирус (RSV)
Респираторно-синцитиальный вирус (RSV) Рассеянный склероз и беременность
Рассеянный склероз и беременность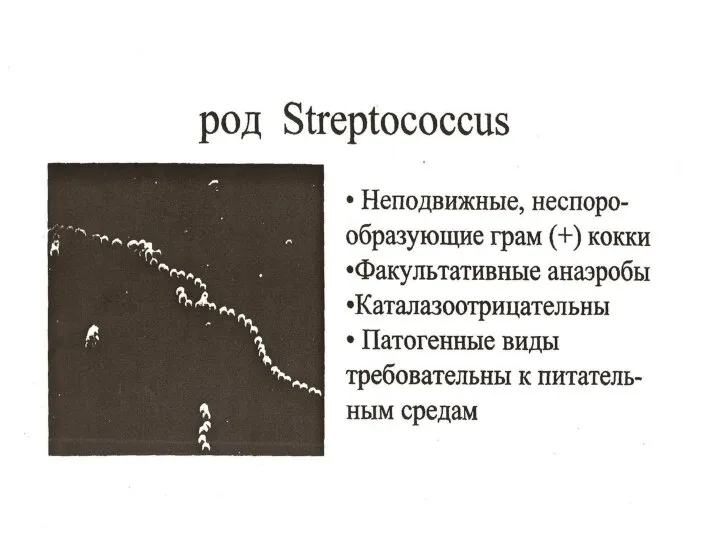 Морфология и тинкториальные свойства Streptococcus
Морфология и тинкториальные свойства Streptococcus Боль и её виды. Механизм боли
Боль и её виды. Механизм боли Паразитарные заболевания кожи человека
Паразитарные заболевания кожи человека Артикуляциялық кескін
Артикуляциялық кескін Зондовые процедуры
Зондовые процедуры Поливитаминные комплексы для людей пожилого возраста
Поливитаминные комплексы для людей пожилого возраста Инновационные разработки ФГБУ НИИПХ Росрезерва в области усовершенствования условий хранения материальных ценностей
Инновационные разработки ФГБУ НИИПХ Росрезерва в области усовершенствования условий хранения материальных ценностей Самоучитель по остеопорозу для населения. (Часть 1)
Самоучитель по остеопорозу для населения. (Часть 1) Цитостатики. Азатиоприн и 6-меркаптопурин
Цитостатики. Азатиоприн и 6-меркаптопурин Гепатоцеллюлярная карцинома
Гепатоцеллюлярная карцинома День борьбы с артритом
День борьбы с артритом Оказание первой помощи детям при несчастных случаях, травмах, отравлениях и состояниях, угрожающих жизни и здоровью
Оказание первой помощи детям при несчастных случаях, травмах, отравлениях и состояниях, угрожающих жизни и здоровью