- Биохимия нервной ткани

Содержание
- 2. План лекции Введение Функции нервной системы и нейрона Особенности нейрона как высокоспециализированного типа клетки Молекулярные основы
- 3. Нервная система обеспечивает структурно-функциональную взаимосвязь в организме регуляторную взаимосвязь между органами и тканями адекватную реакцию организма
- 4. О чём эта лекция? Эту лекцию следует рассматривать как краткое изложение основных аспектов, существенных для понимания
- 5. НЕЙРОН © 2008, В.Г.Зайцев Считается, что единичный нейрон является функциональной (но не структурной!) единицей нервной системы
- 6. Нейрон перерабатывает информацию Восприятие внешнего сигнала (стимула) – одиночного или множества стимулов Обработка сигнала (сигналов) по
- 7. Морфология нейрона Адекватная окраска нейронов разработана в 1873-1885 гг. Камилло Гольджи: фиксация дихроматом калия и импрегнация
- 8. Клеточная архитектура мозга © 2008, В.Г.Зайцев Сантьяго Рамон-и-Кахал, 1888-1891 гг.
- 9. Структурно-функциональное строение © 2008, В.Г.Зайцев
- 10. Полярен ли нейрон? Рамон-и-Кахал и Ван Гегухтен сформулировали «закон динамической поляризации» нейрона По этому закону дендриты
- 11. Ионные транспортные системы, вовлеченные в формирование мембранного потенциала © 2008, В.Г.Зайцев Ионный транспорт Активный транспорт Ионные
- 12. Нервный импульс Нервный импульс волна возбуждения, распространяющаяся по нервному волокну Мембранный потенциал разность потенциалов между внутренней
- 13. Физиологический мембранный потенциал © 2008, В.Г.Зайцев В возбудимых клетках называется потенциалом покоя. Содержание ионов
- 14. Стадии формирования импульса Активация хемозависимых Na-каналов нейромедиатором Открытие потенциал-зависимых Na-каналов в момент достижения порогового потенциала Активация
- 15. Модуляция чувствительности к стимулу © 2008, В.Г.Зайцев
- 16. Нейроны и кальций © 2008, В.Г.Зайцев Потенциал-зависимые кальциевые каналы активируются деполяризацией мембраны Хемозависимые кальциевые каналы активируются
- 17. Нейротоксины © 2008, В.Г.Зайцев Тетродотоксин (рыба фугу, калифорнийский тритон и др.) Сакситоксин (моллюски в водах Аляски)
- 18. Нейротоксины © 2008, В.Г.Зайцев
- 19. Проведение нервного импульса © 2008, В.Г.Зайцев Электротоническое (непрерывное) Немиелинизированные аксоны Медленное (не более 3 м/с) На
- 20. Электротоническое проведение © 2008, В.Г.Зайцев
- 21. Сальтаторное проведение © 2008, В.Г.Зайцев
- 22. Миелиновые волокна © 2008, В.Г.Зайцев
- 23. Химические особенности миелина © 2008, В.Г.Зайцев Липиды Очень высокое содержание (до 75-85% от массы мембраны) Уникальное
- 24. Белок А1 миелиновой оболочки © 2008, В.Г.Зайцев
- 25. Нарушения миелинизации нейронов © 2008, В.Г.Зайцев Рассеянный склероз Периферическая демиелинизирующая нейропатия (синдром Ландри – Гуиллян –
- 26. Рассеянный склероз (Multiple Sclerosis) © 2008, В.Г.Зайцев Хроническое заболевание головного и спинного мозга Наиболее распространенное демиелинизирующее
- 27. Рассеянный склероз © 2008, В.Г.Зайцев Основная причина – демиелинизация с последующим нарушением функций В экспериментах установлено,
- 28. *** Другие дефекты миелиновой оболочки © 2008, В.Г.Зайцев
- 29. Передача сигнала от клетки к клетке © 2008, В.Г.Зайцев «…окончания ветвей древовидного образования одного нейрона не
- 30. Синапсы © 2008, В.Г.Зайцев Электрические и химические Простые и специализированные Возбуждающие и тормозные С непосредственными и
- 31. Электрические синапсы © 2008, В.Г.Зайцев Встречаются редко Роль в ЦНС у млекопитающих неясна Обладают однонаправленным действием
- 32. Синапсы между аксоном (А) и дендритом (Д) © 2008, В.Г.Зайцев
- 33. Синапсы между аксонами и дендритами © 2008, В.Г.Зайцев
- 34. Локализация синапсов разного типа © 2008, В.Г.Зайцев
- 35. Последовательность событий в синапсе © 2008, В.Г.Зайцев Синтез нейромедиатора (в теле нейрона или в окончании аксона)
- 36. Последовательность событий в синапсе © 2008, В.Г.Зайцев
- 37. Аксональный транспорт © 2008, В.Г.Зайцев
- 38. Аксональный транспорт © 2008, В.Г.Зайцев
- 39. Жизненный цикл синаптических пузырьков © 2008, В.Г.Зайцев
- 40. Постсинаптические рецепторы © 2008, В.Г.Зайцев Ионотропные быстрые единый комплекс с ионным каналом работают на открытие каналов
- 41. Постсинаптические рецепторы © 2008, В.Г.Зайцев
- 42. Непептидные нейромедиаторы © 2008, В.Г.Зайцев
- 43. Инактивация катехоламинов © 2008, В.Г.Зайцев
- 44. Инактивация норадреналина © 2008, В.Г.Зайцев
- 45. Инактивация дофамина в регуляции синаптической активности © 2008, В.Г.Зайцев
- 46. Связь генотипа с поведением © 2008, В.Г.Зайцев
- 47. Связь генотипа с поведением © 2008, В.Г.Зайцев
- 48. Связь генотипа с поведением © 2008, В.Г.Зайцев
- 49. Латеральное торможение © 2008, В.Г.Зайцев
- 50. NO и долговременная потенциация © 2008, В.Г.Зайцев
- 51. Ретротранспорт ГАМК © 2008, В.Г.Зайцев
- 52. Действие лекарственных препаратов © 2008, В.Г.Зайцев
- 53. Энергетический обмен © 2008, В.Г.Зайцев Источник АТФ Только (!) аэробный распад глюкозы Аэробная продукция АТФ может
- 54. Углеводный обмен © 2008, В.Г.Зайцев Потребление глюкозы Гексокиназа имеет очень низкое значение Km и оченьы высокое
- 55. Обмен белков © 2008, В.Г.Зайцев Сильнокислый белок S-100 (изоформа мозга) преимущественно в клетках глии снижение содержания
- 56. Обмен аминокислот и пептидов © 2008, В.Г.Зайцев Общее содержание До 30-100 мМ, что в несколько (до
- 57. Шунт ГАМК © 2008, В.Г.Зайцев
- 58. Метаболизм ГАМК © 2008, В.Г.Зайцев
- 59. Регуляция глутаматдекарбоксилазы © 2008, В.Г.Зайцев
- 60. Обмен глутамата в нейронах © 2008, В.Г.Зайцев
- 61. Особенности обмена пептидных нейромедиаторов © 2008, В.Г.Зайцев
- 62. Обмен липидов © 2008, В.Г.Зайцев Обмен Скорость обновления очень низкая! – особенно в миелиновой мембране Особенно
- 63. Метаболизм ацетилхолина © 2008, В.Г.Зайцев
- 64. Обмен нуклеотидов © 2008, В.Г.Зайцев Скорость обмена Очень высокая скорость обмена РНК и очень низкая ДНК
- 65. Регуляция роста нейронов © 2008, В.Г.Зайцев
- 66. Фактор роста нервов © 2008, В.Г.Зайцев
- 67. Регуляция роста нейронов © 2008, В.Г.Зайцев
- 68. Нейроны и формирование условных рефлексов © 2008, В.Г.Зайцев
- 69. Астроциты и олигодендроциты © 2008, В.Г.Зайцев
- 70. Глия и функции нервной ткани © 2008, В.Г.Зайцев
- 71. Активация макрофагов клетками головного мозга © 2008, В.Г.Зайцев
- 72. Вовлеченность иммунных механизмов в повреждение нервной ткани © 2008, В.Г.Зайцев Ly – лимфоциты; Mac – макрофаги;
- 74. Скачать презентацию
Слайд 2План лекции
Введение
Функции нервной системы и нейрона
Особенности нейрона как высокоспециализированного типа клетки
Молекулярные основы
План лекции
Введение
Функции нервной системы и нейрона
Особенности нейрона как высокоспециализированного типа клетки
Молекулярные основы

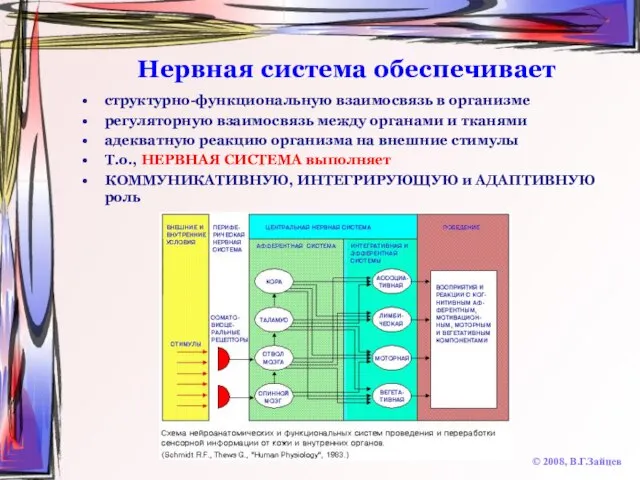
Слайд 3Нервная система обеспечивает
структурно-функциональную взаимосвязь в организме
регуляторную взаимосвязь между органами и тканями
адекватную реакцию
Нервная система обеспечивает
структурно-функциональную взаимосвязь в организме
регуляторную взаимосвязь между органами и тканями
адекватную реакцию
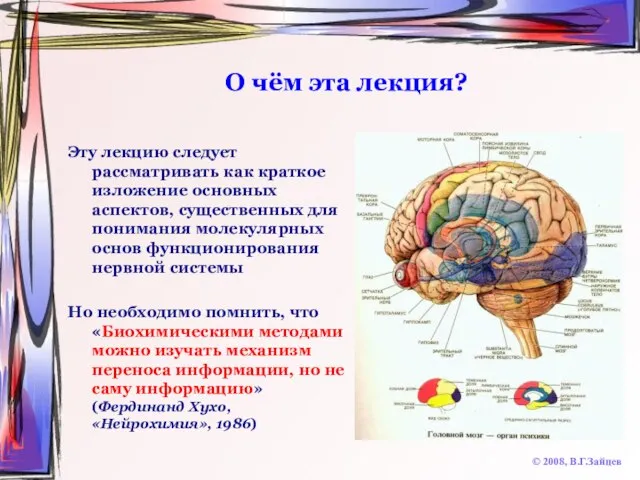
Слайд 4О чём эта лекция?
Эту лекцию следует рассматривать как краткое изложение основных аспектов,
О чём эта лекция?
Эту лекцию следует рассматривать как краткое изложение основных аспектов,
Слайд 5НЕЙРОН
© 2008, В.Г.Зайцев
Считается, что единичный нейрон является функциональной (но не структурной!) единицей
НЕЙРОН
© 2008, В.Г.Зайцев
Считается, что единичный нейрон является функциональной (но не структурной!) единицей

Слайд 6Нейрон перерабатывает информацию
Восприятие внешнего сигнала (стимула) – одиночного или множества стимулов
Обработка сигнала
Нейрон перерабатывает информацию
Восприятие внешнего сигнала (стимула) – одиночного или множества стимулов
Обработка сигнала

Слайд 7Морфология нейрона
Адекватная окраска нейронов разработана в 1873-1885 гг. Камилло Гольджи:
фиксация дихроматом калия
Морфология нейрона
Адекватная окраска нейронов разработана в 1873-1885 гг. Камилло Гольджи:
фиксация дихроматом калия

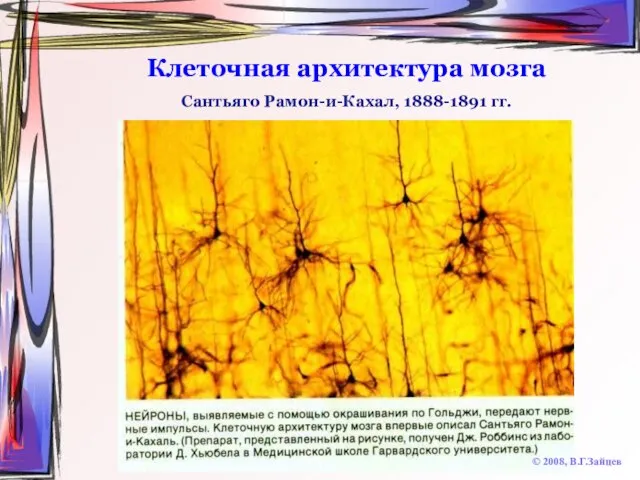
Слайд 8Клеточная архитектура мозга
© 2008, В.Г.Зайцев
Сантьяго Рамон-и-Кахал, 1888-1891 гг.
Клеточная архитектура мозга
© 2008, В.Г.Зайцев
Сантьяго Рамон-и-Кахал, 1888-1891 гг.
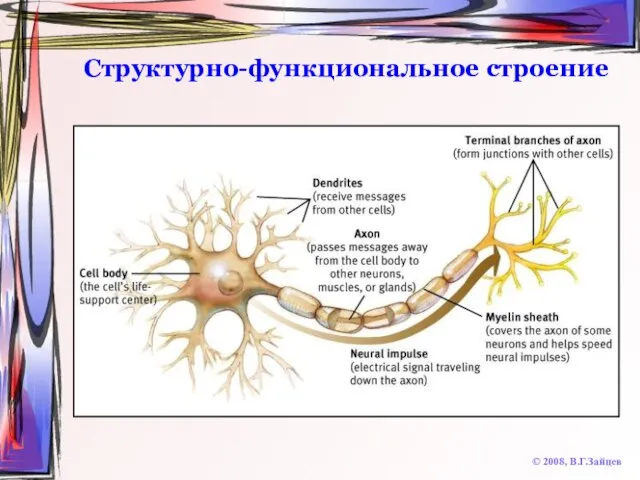
Слайд 9Структурно-функциональное строение
© 2008, В.Г.Зайцев
Структурно-функциональное строение
© 2008, В.Г.Зайцев
Слайд 10Полярен ли нейрон?
Рамон-и-Кахал и Ван Гегухтен сформулировали «закон динамической поляризации» нейрона
По этому
Полярен ли нейрон?
Рамон-и-Кахал и Ван Гегухтен сформулировали «закон динамической поляризации» нейрона
По этому

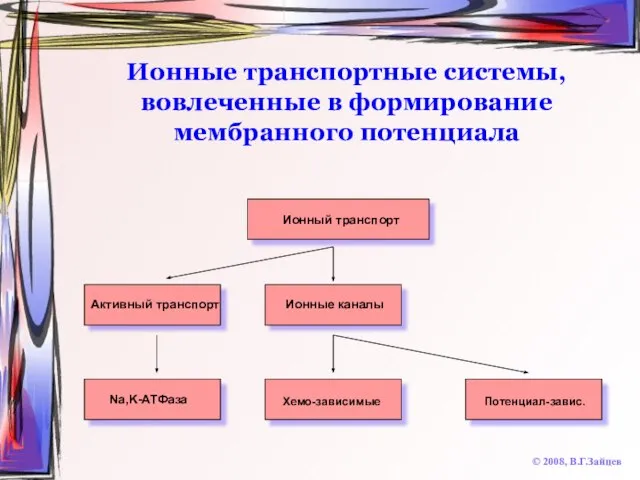
Слайд 11Ионные транспортные системы, вовлеченные в формирование мембранного потенциала
© 2008, В.Г.Зайцев
Ионный транспорт
Активный транспорт
Ионные
Ионные транспортные системы, вовлеченные в формирование мембранного потенциала
© 2008, В.Г.Зайцев
Ионный транспорт
Активный транспорт
Ионные
Слайд 12Нервный импульс
Нервный импульс
волна возбуждения, распространяющаяся по нервному волокну
Мембранный потенциал
разность потенциалов между внутренней
Нервный импульс
Нервный импульс
волна возбуждения, распространяющаяся по нервному волокну
Мембранный потенциал
разность потенциалов между внутренней

Слайд 13Физиологический мембранный потенциал
© 2008, В.Г.Зайцев
В возбудимых клетках называется потенциалом покоя.
Содержание ионов
Физиологический мембранный потенциал
© 2008, В.Г.Зайцев
В возбудимых клетках называется потенциалом покоя.
Содержание ионов

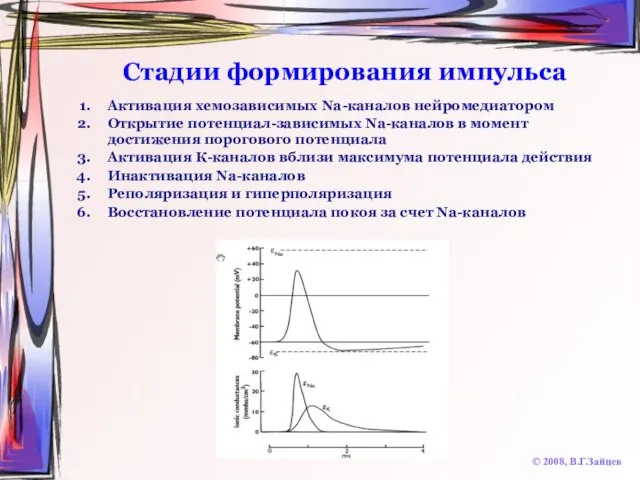
Слайд 14Стадии формирования импульса
Активация хемозависимых Na-каналов нейромедиатором
Открытие потенциал-зависимых Na-каналов в момент достижения порогового
Стадии формирования импульса
Активация хемозависимых Na-каналов нейромедиатором
Открытие потенциал-зависимых Na-каналов в момент достижения порогового
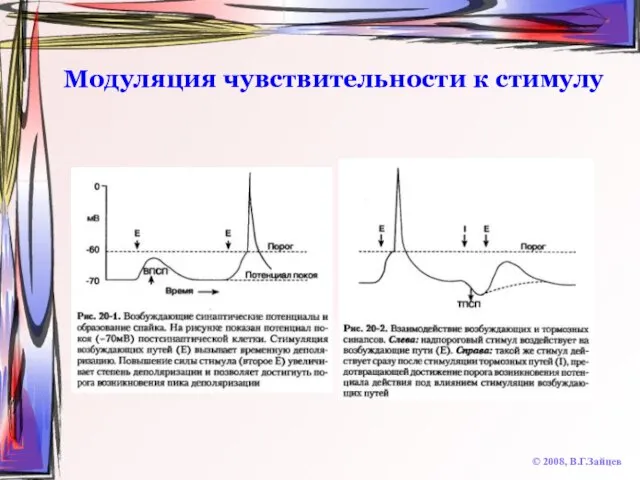
Слайд 15Модуляция чувствительности к стимулу
© 2008, В.Г.Зайцев
Модуляция чувствительности к стимулу
© 2008, В.Г.Зайцев
Слайд 16Нейроны и кальций
© 2008, В.Г.Зайцев
Потенциал-зависимые кальциевые каналы активируются деполяризацией мембраны
Хемозависимые кальциевые каналы
Нейроны и кальций
© 2008, В.Г.Зайцев
Потенциал-зависимые кальциевые каналы активируются деполяризацией мембраны
Хемозависимые кальциевые каналы

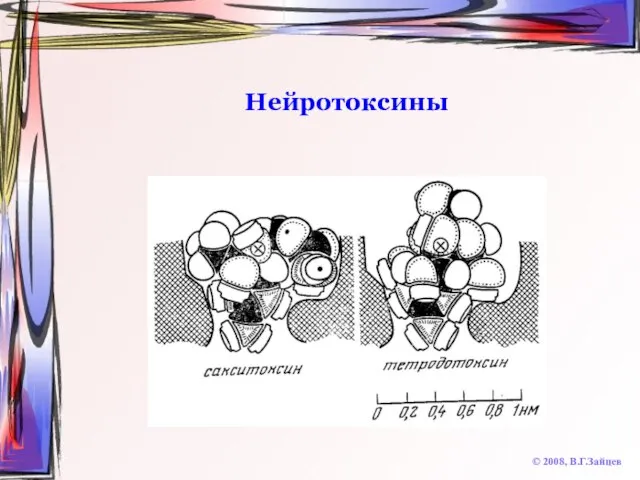
Слайд 17Нейротоксины
© 2008, В.Г.Зайцев
Тетродотоксин (рыба фугу, калифорнийский тритон и др.)
Сакситоксин (моллюски в водах
Нейротоксины
© 2008, В.Г.Зайцев
Тетродотоксин (рыба фугу, калифорнийский тритон и др.)
Сакситоксин (моллюски в водах

Слайд 18Нейротоксины
© 2008, В.Г.Зайцев
Нейротоксины
© 2008, В.Г.Зайцев
Слайд 19Проведение нервного импульса
© 2008, В.Г.Зайцев
Электротоническое (непрерывное)
Немиелинизированные аксоны
Медленное (не более 3 м/с)
На короткие
Проведение нервного импульса
© 2008, В.Г.Зайцев
Электротоническое (непрерывное)
Немиелинизированные аксоны
Медленное (не более 3 м/с)
На короткие

Слайд 20Электротоническое проведение
© 2008, В.Г.Зайцев
Электротоническое проведение
© 2008, В.Г.Зайцев

Слайд 21Сальтаторное проведение
© 2008, В.Г.Зайцев
Сальтаторное проведение
© 2008, В.Г.Зайцев

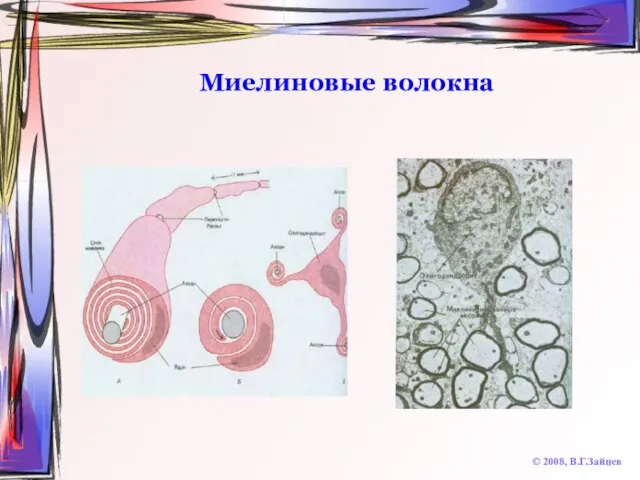
Слайд 22Миелиновые волокна
© 2008, В.Г.Зайцев
Миелиновые волокна
© 2008, В.Г.Зайцев
Слайд 23Химические особенности миелина
© 2008, В.Г.Зайцев
Липиды
Очень высокое содержание (до 75-85% от массы мембраны)
Уникальное
Химические особенности миелина
© 2008, В.Г.Зайцев
Липиды
Очень высокое содержание (до 75-85% от массы мембраны)
Уникальное

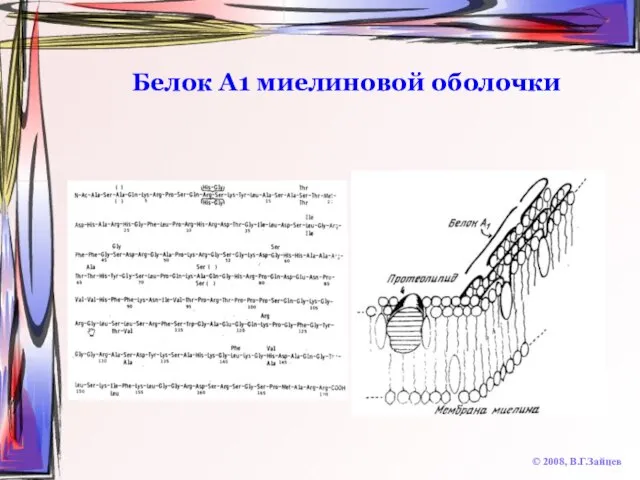
Слайд 24Белок А1 миелиновой оболочки
© 2008, В.Г.Зайцев
Белок А1 миелиновой оболочки
© 2008, В.Г.Зайцев
Слайд 25Нарушения миелинизации нейронов
© 2008, В.Г.Зайцев
Рассеянный склероз
Периферическая демиелинизирующая нейропатия (синдром Ландри – Гуиллян
Нарушения миелинизации нейронов
© 2008, В.Г.Зайцев
Рассеянный склероз
Периферическая демиелинизирующая нейропатия (синдром Ландри – Гуиллян

Слайд 26Рассеянный склероз (Multiple Sclerosis)
© 2008, В.Г.Зайцев
Хроническое заболевание головного и спинного мозга
Наиболее
Рассеянный склероз (Multiple Sclerosis)
© 2008, В.Г.Зайцев
Хроническое заболевание головного и спинного мозга
Наиболее

Слайд 27Рассеянный склероз
© 2008, В.Г.Зайцев
Основная причина – демиелинизация с последующим нарушением функций
В экспериментах
Рассеянный склероз
© 2008, В.Г.Зайцев
Основная причина – демиелинизация с последующим нарушением функций
В экспериментах

Слайд 28*** Другие дефекты миелиновой оболочки
© 2008, В.Г.Зайцев
*** Другие дефекты миелиновой оболочки
© 2008, В.Г.Зайцев

Слайд 29Передача сигнала от клетки к клетке
© 2008, В.Г.Зайцев
«…окончания ветвей древовидного образования одного
Передача сигнала от клетки к клетке
© 2008, В.Г.Зайцев
«…окончания ветвей древовидного образования одного

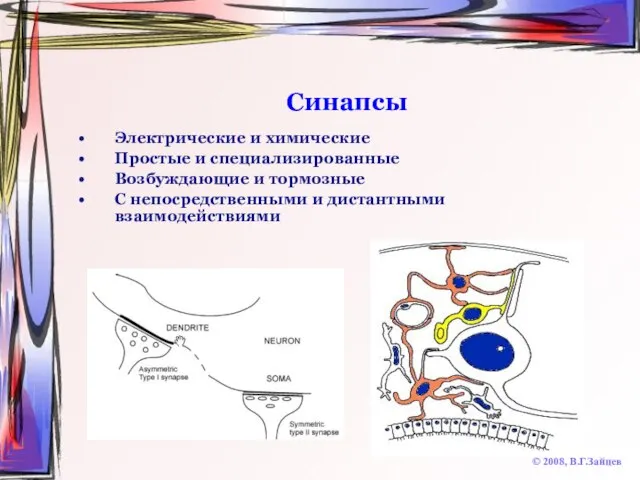
Слайд 30Синапсы
© 2008, В.Г.Зайцев
Электрические и химические
Простые и специализированные
Возбуждающие и тормозные
С непосредственными и дистантными
Синапсы
© 2008, В.Г.Зайцев
Электрические и химические
Простые и специализированные
Возбуждающие и тормозные
С непосредственными и дистантными
Слайд 31Электрические синапсы
© 2008, В.Г.Зайцев
Встречаются редко
Роль в ЦНС у млекопитающих неясна
Обладают однонаправленным действием
Имеют
Электрические синапсы
© 2008, В.Г.Зайцев
Встречаются редко
Роль в ЦНС у млекопитающих неясна
Обладают однонаправленным действием
Имеют

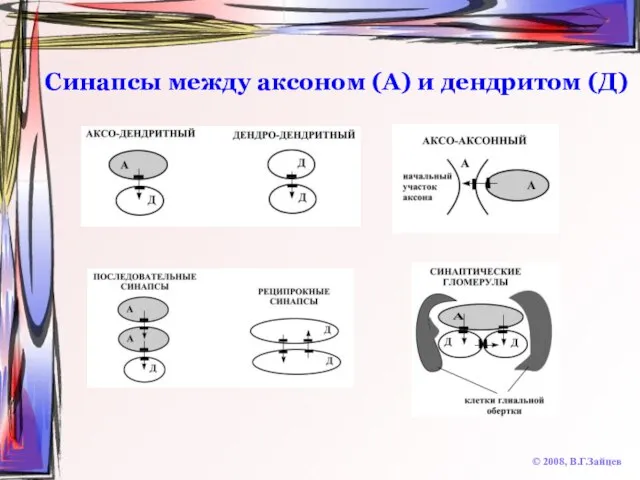
Слайд 32Синапсы между аксоном (А) и дендритом (Д)
© 2008, В.Г.Зайцев
Синапсы между аксоном (А) и дендритом (Д)
© 2008, В.Г.Зайцев
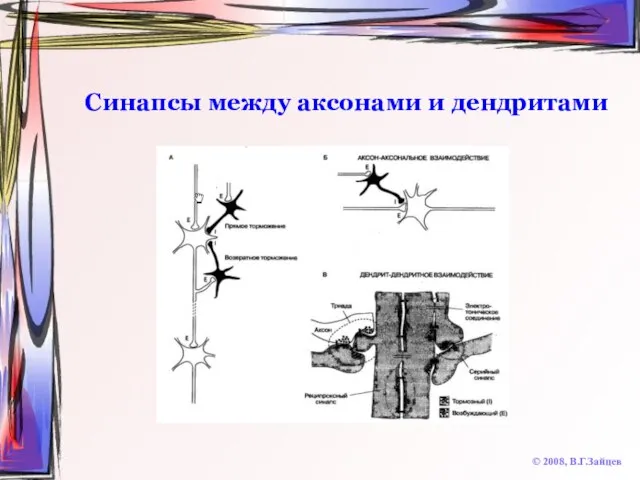
Слайд 33Синапсы между аксонами и дендритами
© 2008, В.Г.Зайцев
Синапсы между аксонами и дендритами
© 2008, В.Г.Зайцев
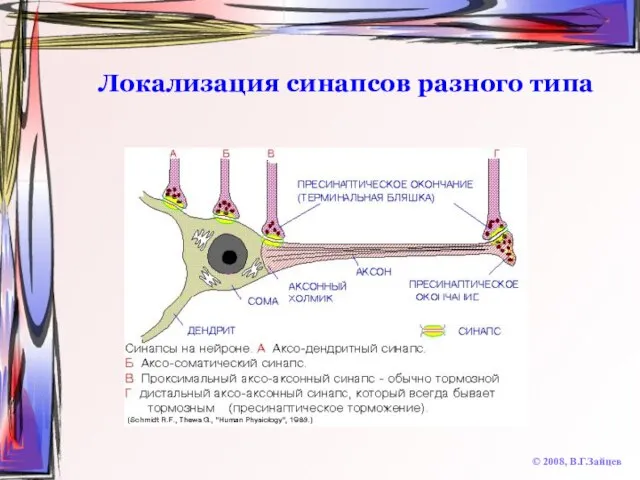
Слайд 34Локализация синапсов разного типа
© 2008, В.Г.Зайцев
Локализация синапсов разного типа
© 2008, В.Г.Зайцев
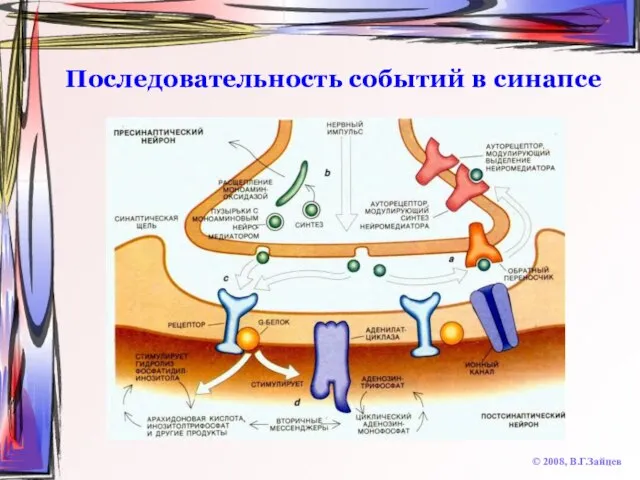
Слайд 35Последовательность событий в синапсе
© 2008, В.Г.Зайцев
Синтез нейромедиатора (в теле нейрона или в
Последовательность событий в синапсе
© 2008, В.Г.Зайцев
Синтез нейромедиатора (в теле нейрона или в

Слайд 36Последовательность событий в синапсе
© 2008, В.Г.Зайцев
Последовательность событий в синапсе
© 2008, В.Г.Зайцев
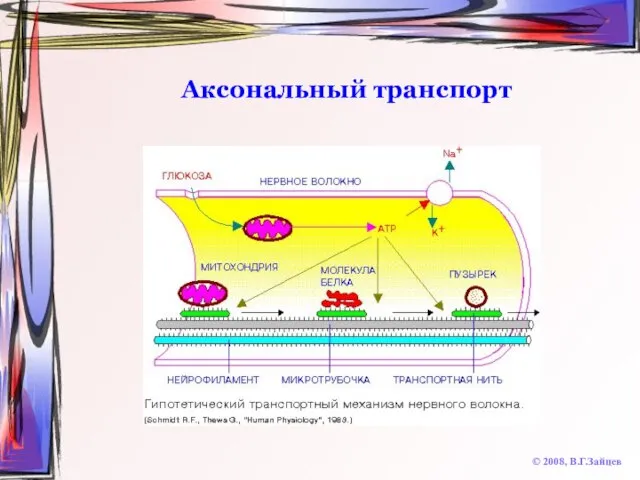
Слайд 37Аксональный транспорт
© 2008, В.Г.Зайцев
Аксональный транспорт
© 2008, В.Г.Зайцев

Слайд 38Аксональный транспорт
© 2008, В.Г.Зайцев
Аксональный транспорт
© 2008, В.Г.Зайцев
Слайд 39Жизненный цикл синаптических пузырьков
© 2008, В.Г.Зайцев
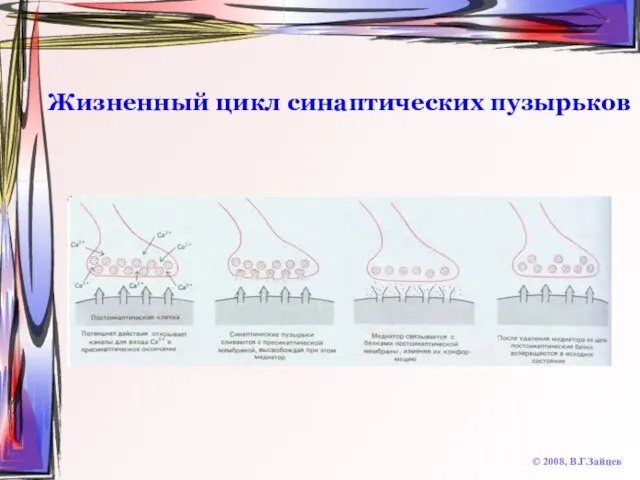
Жизненный цикл синаптических пузырьков
© 2008, В.Г.Зайцев
Слайд 40Постсинаптические рецепторы
© 2008, В.Г.Зайцев
Ионотропные
быстрые
единый комплекс с ионным каналом
работают на открытие каналов
никотиновые холинорецепторы,
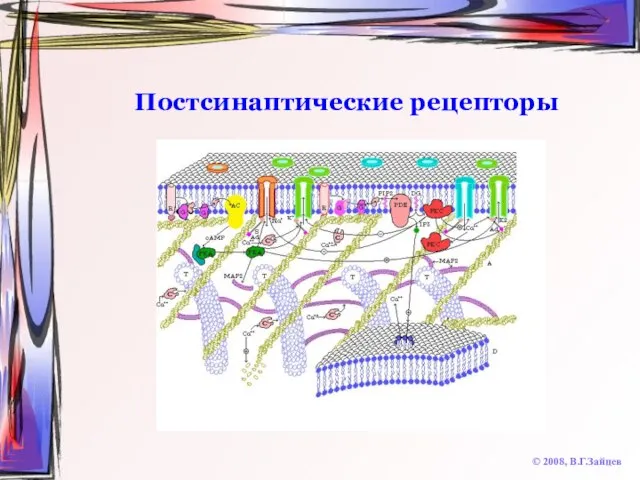
Постсинаптические рецепторы
© 2008, В.Г.Зайцев
Ионотропные
быстрые
единый комплекс с ионным каналом
работают на открытие каналов
никотиновые холинорецепторы,

Слайд 41Постсинаптические рецепторы
© 2008, В.Г.Зайцев
Постсинаптические рецепторы
© 2008, В.Г.Зайцев
Слайд 42Непептидные нейромедиаторы
© 2008, В.Г.Зайцев
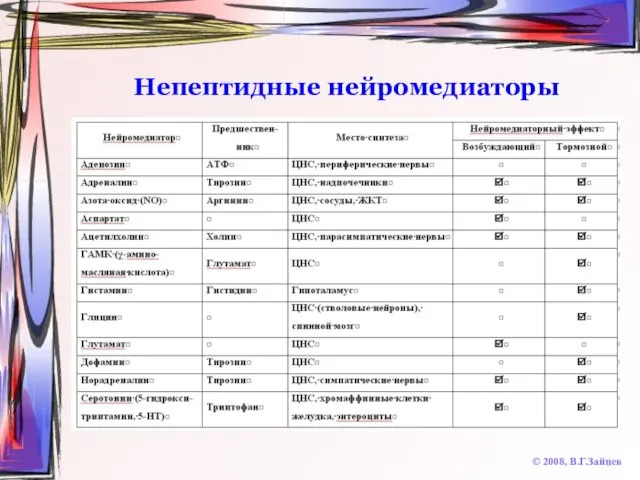
Непептидные нейромедиаторы
© 2008, В.Г.Зайцев
Слайд 43Инактивация катехоламинов
© 2008, В.Г.Зайцев
Инактивация катехоламинов
© 2008, В.Г.Зайцев

Слайд 44Инактивация норадреналина
© 2008, В.Г.Зайцев
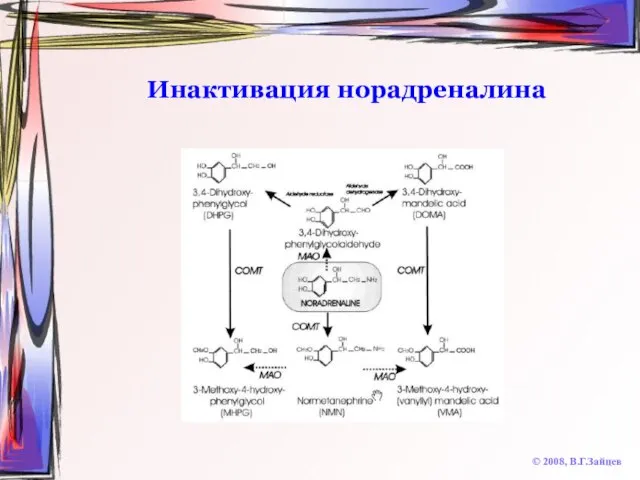
Инактивация норадреналина
© 2008, В.Г.Зайцев
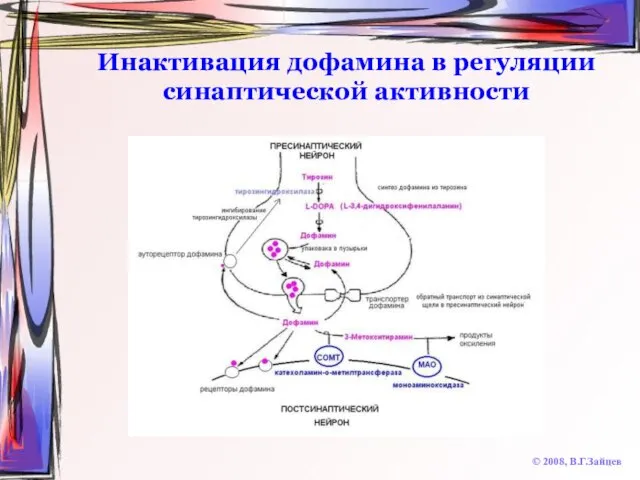
Слайд 45Инактивация дофамина в регуляции синаптической активности
© 2008, В.Г.Зайцев
Инактивация дофамина в регуляции синаптической активности
© 2008, В.Г.Зайцев
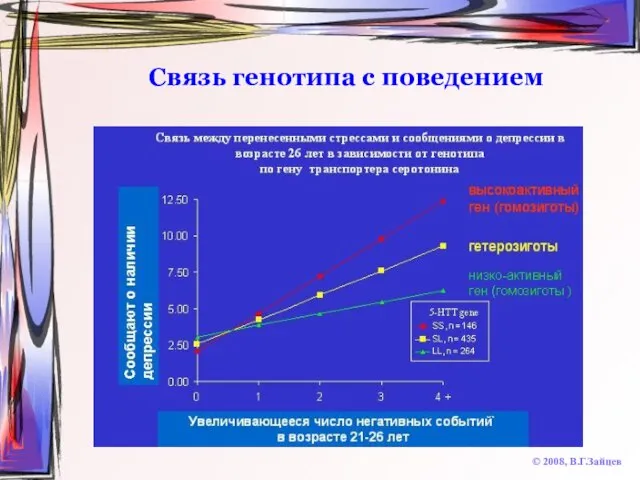
Слайд 46Связь генотипа с поведением
© 2008, В.Г.Зайцев
Связь генотипа с поведением
© 2008, В.Г.Зайцев
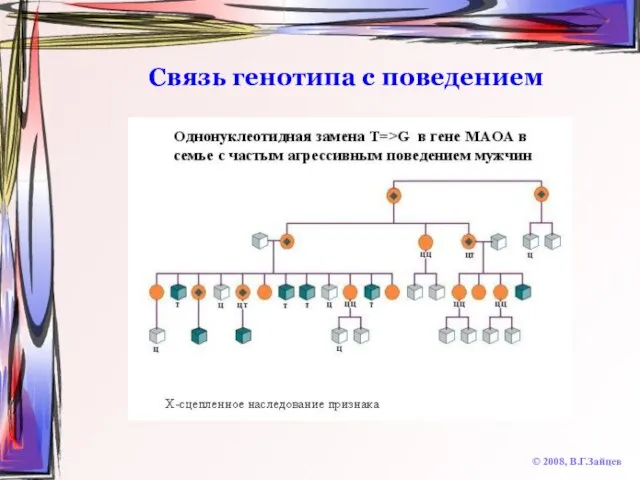
Слайд 47Связь генотипа с поведением
© 2008, В.Г.Зайцев
Связь генотипа с поведением
© 2008, В.Г.Зайцев
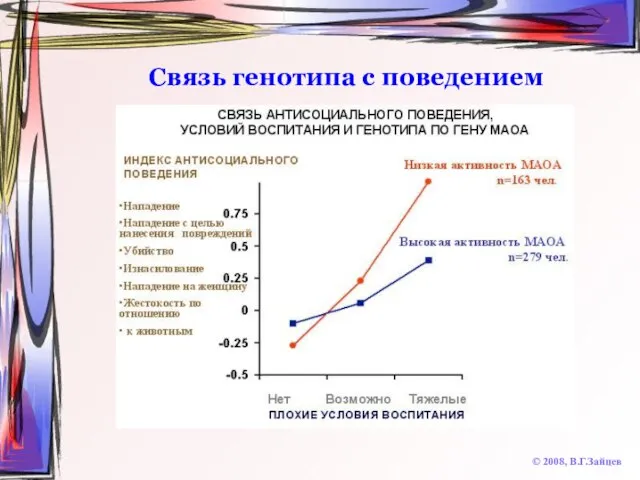
Слайд 48Связь генотипа с поведением
© 2008, В.Г.Зайцев
Связь генотипа с поведением
© 2008, В.Г.Зайцев
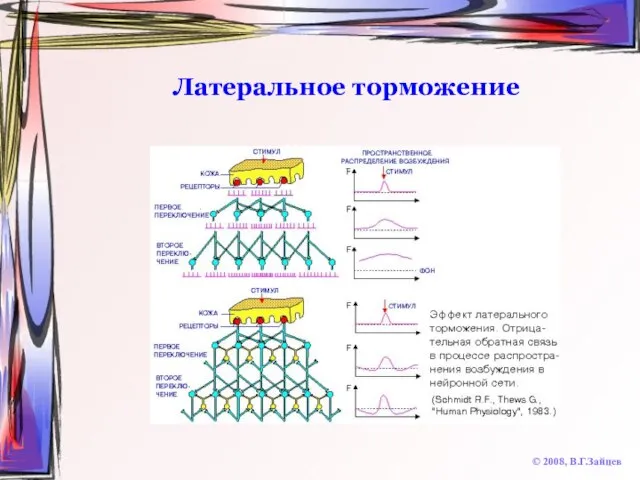
Слайд 49Латеральное торможение
© 2008, В.Г.Зайцев
Латеральное торможение
© 2008, В.Г.Зайцев
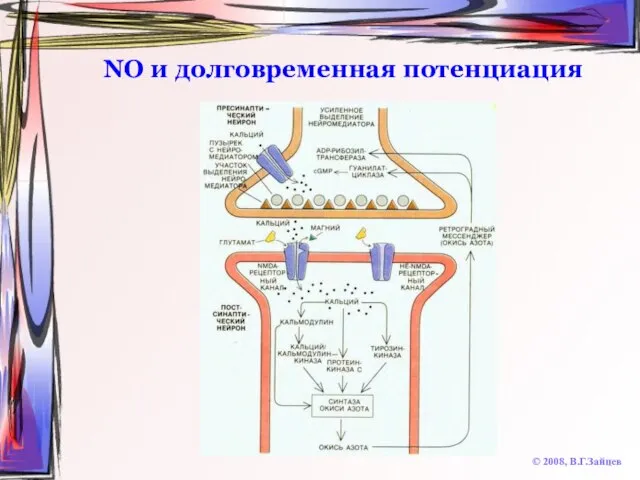
Слайд 50NO и долговременная потенциация
© 2008, В.Г.Зайцев
NO и долговременная потенциация
© 2008, В.Г.Зайцев
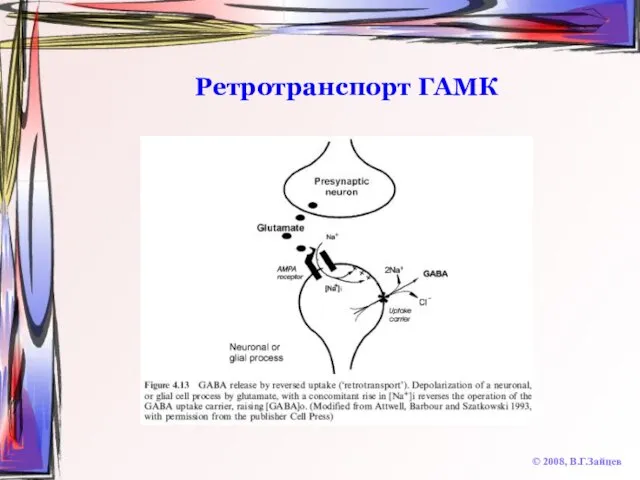
Слайд 51Ретротранспорт ГАМК
© 2008, В.Г.Зайцев
Ретротранспорт ГАМК
© 2008, В.Г.Зайцев
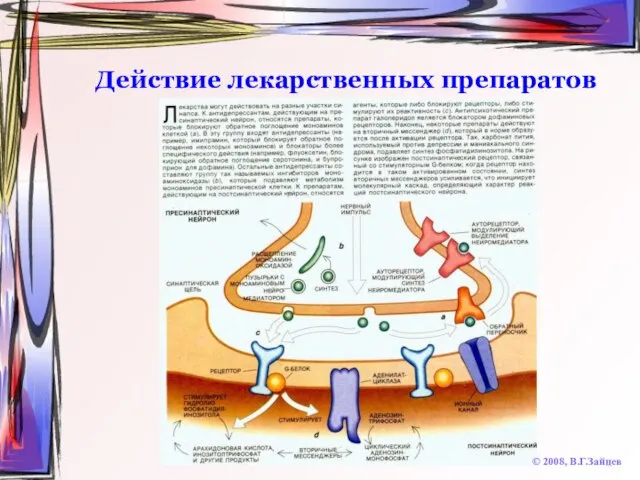
Слайд 52Действие лекарственных препаратов
© 2008, В.Г.Зайцев
Действие лекарственных препаратов
© 2008, В.Г.Зайцев
Слайд 53Энергетический обмен
© 2008, В.Г.Зайцев
Источник АТФ
Только (!) аэробный распад глюкозы
Аэробная продукция АТФ может
Энергетический обмен
© 2008, В.Г.Зайцев
Источник АТФ
Только (!) аэробный распад глюкозы
Аэробная продукция АТФ может

Слайд 54Углеводный обмен
© 2008, В.Г.Зайцев
Потребление глюкозы
Гексокиназа имеет очень низкое значение Km и оченьы
Углеводный обмен
© 2008, В.Г.Зайцев
Потребление глюкозы
Гексокиназа имеет очень низкое значение Km и оченьы

Слайд 55Обмен белков
© 2008, В.Г.Зайцев
Сильнокислый белок S-100 (изоформа мозга)
преимущественно в клетках глии
снижение содержания
Обмен белков
© 2008, В.Г.Зайцев
Сильнокислый белок S-100 (изоформа мозга)
преимущественно в клетках глии
снижение содержания

Слайд 56Обмен аминокислот и пептидов
© 2008, В.Г.Зайцев
Общее содержание
До 30-100 мМ, что в несколько
Обмен аминокислот и пептидов
© 2008, В.Г.Зайцев
Общее содержание
До 30-100 мМ, что в несколько

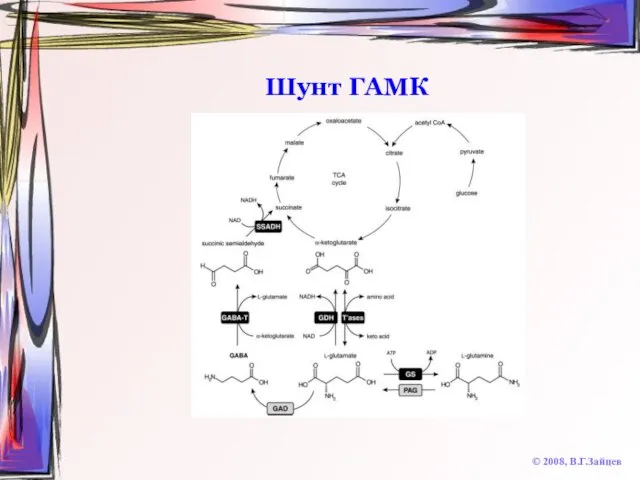
Слайд 57Шунт ГАМК
© 2008, В.Г.Зайцев
Шунт ГАМК
© 2008, В.Г.Зайцев
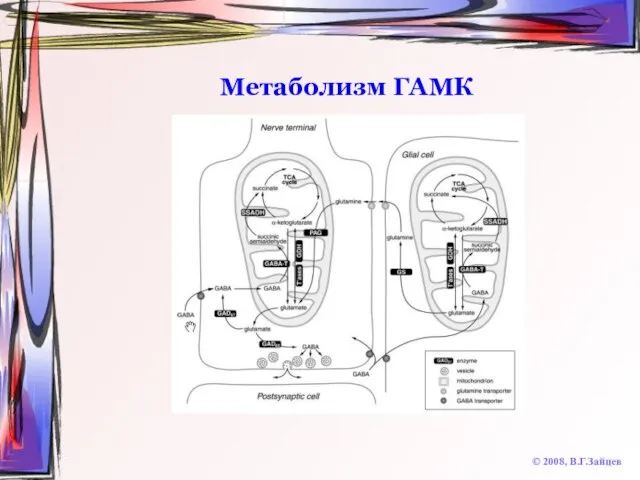
Слайд 58Метаболизм ГАМК
© 2008, В.Г.Зайцев
Метаболизм ГАМК
© 2008, В.Г.Зайцев
Слайд 59Регуляция глутаматдекарбоксилазы
© 2008, В.Г.Зайцев
Регуляция глутаматдекарбоксилазы
© 2008, В.Г.Зайцев

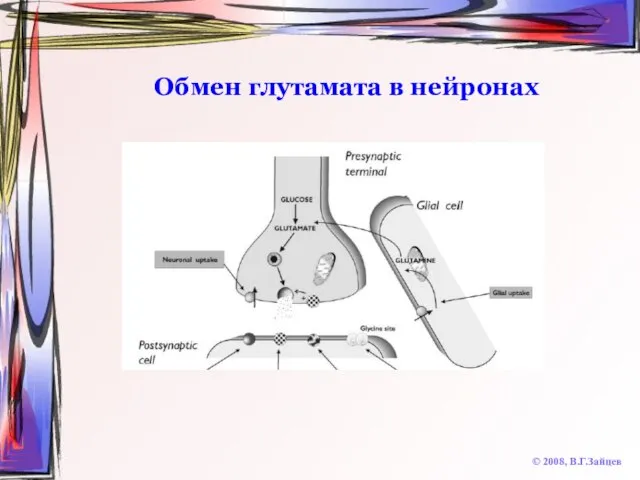
Слайд 60Обмен глутамата в нейронах
© 2008, В.Г.Зайцев
Обмен глутамата в нейронах
© 2008, В.Г.Зайцев
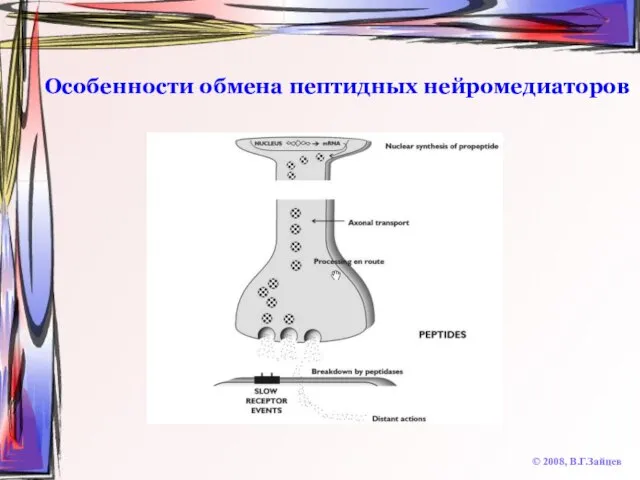
Слайд 61Особенности обмена пептидных нейромедиаторов
© 2008, В.Г.Зайцев
Особенности обмена пептидных нейромедиаторов
© 2008, В.Г.Зайцев
Слайд 62Обмен липидов
© 2008, В.Г.Зайцев
Обмен
Скорость обновления очень низкая! – особенно в миелиновой мембране
Особенно
Обмен липидов
© 2008, В.Г.Зайцев
Обмен
Скорость обновления очень низкая! – особенно в миелиновой мембране
Особенно

Слайд 63Метаболизм ацетилхолина
© 2008, В.Г.Зайцев
Метаболизм ацетилхолина
© 2008, В.Г.Зайцев

Слайд 64Обмен нуклеотидов
© 2008, В.Г.Зайцев
Скорость обмена
Очень высокая скорость обмена РНК и очень низкая
Обмен нуклеотидов
© 2008, В.Г.Зайцев
Скорость обмена
Очень высокая скорость обмена РНК и очень низкая

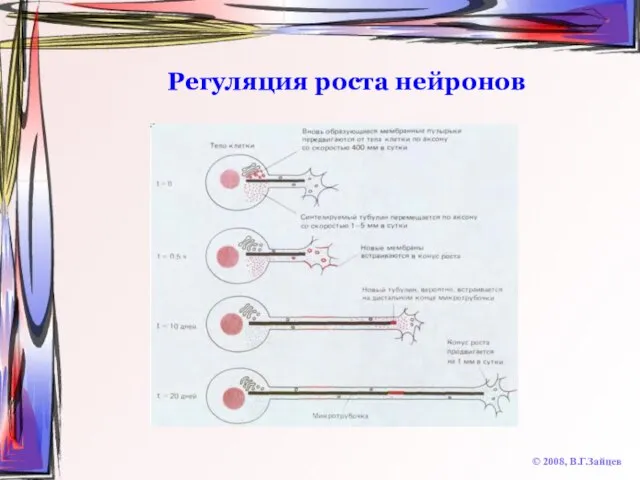
Слайд 65Регуляция роста нейронов
© 2008, В.Г.Зайцев
Регуляция роста нейронов
© 2008, В.Г.Зайцев
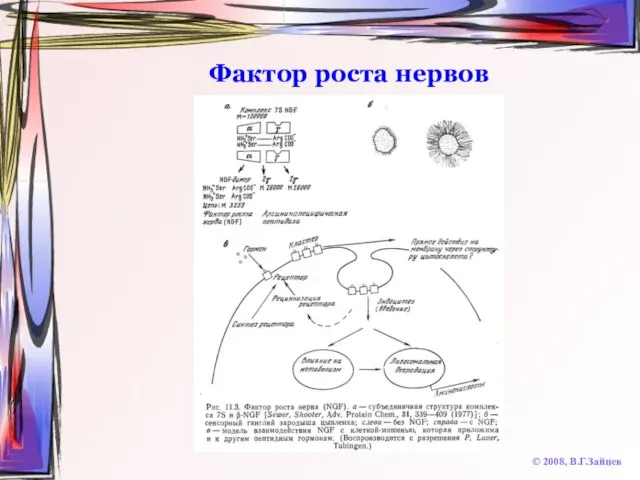
Слайд 66Фактор роста нервов
© 2008, В.Г.Зайцев
Фактор роста нервов
© 2008, В.Г.Зайцев
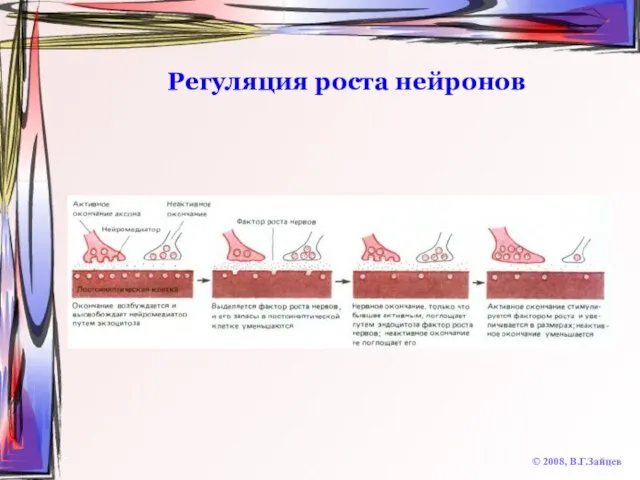
Слайд 67Регуляция роста нейронов
© 2008, В.Г.Зайцев
Регуляция роста нейронов
© 2008, В.Г.Зайцев
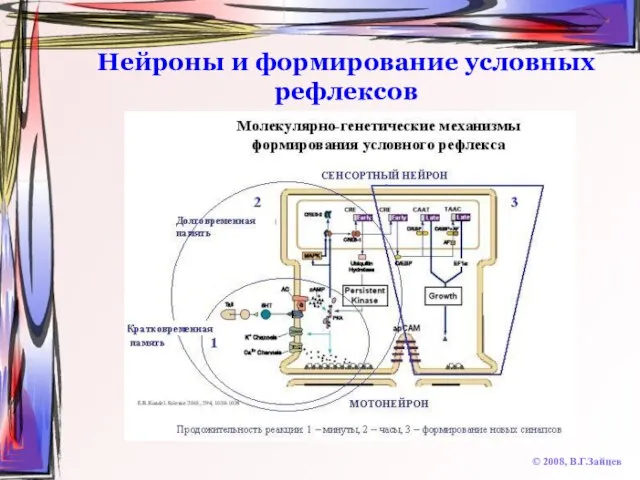
Слайд 68Нейроны и формирование условных рефлексов
© 2008, В.Г.Зайцев
Нейроны и формирование условных рефлексов
© 2008, В.Г.Зайцев
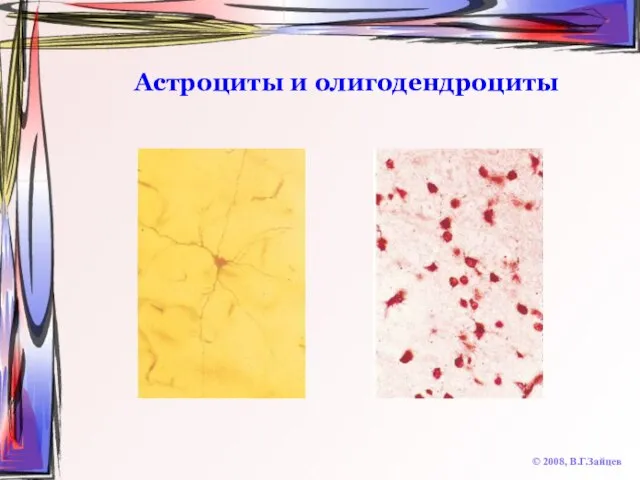
Слайд 69Астроциты и олигодендроциты
© 2008, В.Г.Зайцев
Астроциты и олигодендроциты
© 2008, В.Г.Зайцев
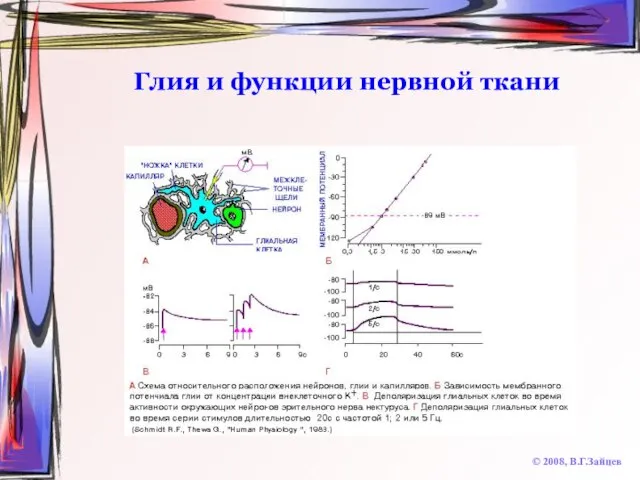
Слайд 70Глия и функции нервной ткани
© 2008, В.Г.Зайцев
Глия и функции нервной ткани
© 2008, В.Г.Зайцев
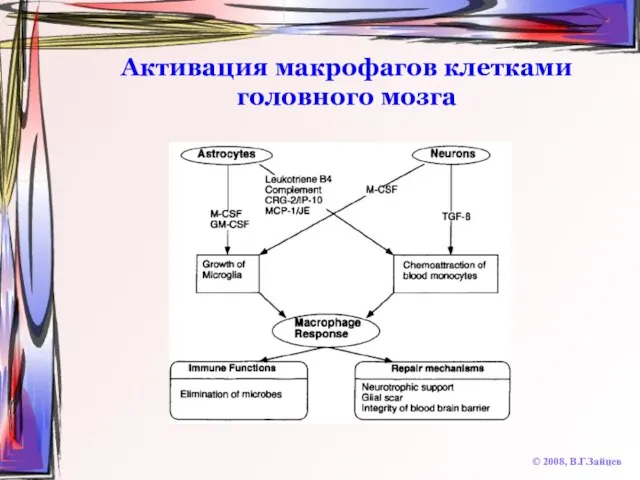
Слайд 71Активация макрофагов клетками головного мозга
© 2008, В.Г.Зайцев
Активация макрофагов клетками головного мозга
© 2008, В.Г.Зайцев
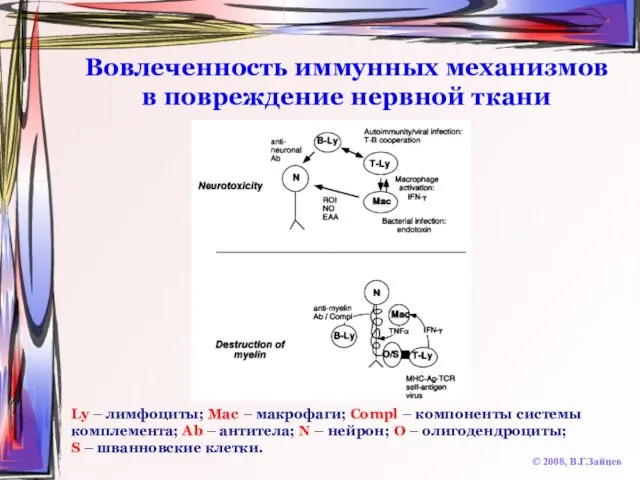
Слайд 72Вовлеченность иммунных механизмов в повреждение нервной ткани
© 2008, В.Г.Зайцев
Ly – лимфоциты; Mac
Вовлеченность иммунных механизмов в повреждение нервной ткани
© 2008, В.Г.Зайцев
Ly – лимфоциты; Mac
 Презентация на тему Пожар в лесу
Презентация на тему Пожар в лесу Огорсад иль садород?
Огорсад иль садород? Числа от 1 до 10
Числа от 1 до 10 Деятельностный подход на уроках математики
Деятельностный подход на уроках математики Transformation of a Drawing
Transformation of a Drawing Староакульшетская школа
Староакульшетская школа Шахматные фигуры и начальная позиция (урок № 7)
Шахматные фигуры и начальная позиция (урок № 7) Информация, ее виды и свойства
Информация, ее виды и свойства Какую роль играет вкус,зрение,обоняние в жизнедеятельности человека
Какую роль играет вкус,зрение,обоняние в жизнедеятельности человека Лабынкырский чёрт
Лабынкырский чёрт "Основы религиозных культур и светской этики"
"Основы религиозных культур и светской этики" Презентация на тему Корненожки. Амеба обыкновенная
Презентация на тему Корненожки. Амеба обыкновенная Керамзитовый завод Козульки
Керамзитовый завод Козульки Планерка обработчиков
Планерка обработчиков 20121009_duhovnost
20121009_duhovnost Части речи 3 класс
Части речи 3 класс Стратегический менеджмент в условиях спонтанных изменений
Стратегический менеджмент в условиях спонтанных изменений Протоколы маршрутизации
Протоколы маршрутизации  Презентация на тему Многогранники и их различия
Презентация на тему Многогранники и их различия Эквадор
Эквадор Золотое правило НРАВСТВЕННОСТИ
Золотое правило НРАВСТВЕННОСТИ Липецкий государственный технический университет Кафедра прикладной механики
Липецкий государственный технический университет Кафедра прикладной механики ЗАГАДКА МЁБИУСА
ЗАГАДКА МЁБИУСА Russia is my motherland
Russia is my motherland Многообразие органических веществ
Многообразие органических веществ Классификация химических реакций (11 класс)
Классификация химических реакций (11 класс) Фредерик Уинслоу Тейлор. Ду́глас Мак-Гре́гор Теория Х
Фредерик Уинслоу Тейлор. Ду́глас Мак-Гре́гор Теория Х Педагогика лек6 -2022
Педагогика лек6 -2022