- Індивідуальний розвиток огранізму

Содержание
- 2. Етапи індивідуального розвитку організмів Індивідуальний розвиток організму від його зародження до смерті називається онтогенезом. Цей процес
- 3. Зародковий (ембріональний) період розвитку Цей період триває від моменту запліднення яйцеклітини до початку вільного самостійного життя
- 4. Дроблення та утворення бластули (на прикладі ссавців) Відразу після запліднення починається процес поділу зиготи, який називають
- 5. Типи дроблення яйцеклітин: а — повне рівномірне ізолецитальної яйцеклітини; б — повне нерівномірне телолецитальної яйцеклітини; в
- 6. Утворення гаструли Сукупність процесів, які забезпечують утворення гаструли, називають гаструляцією. При цьому із бластодерми різними способами
- 8. Стадія утворення трьох зародкових листків На наступній стадії утворення трьох зародкових листків — між екто- й
- 9. Типи утворення мезодерми
- 10. Диференціація тканин і органів під час зародкового розвитку (процес гістогенезу та органогенезу) В кожному зародковому листку
- 11. Закладання нервової трубкн: а: 1 — мезодермальний дах первинної кишки з бічними пластинками (2) розміщений між
- 12. Формування тканин та органів зародка (на прикладі ланцентника)
- 13. Сперматозоїд рухається по слизовій оболонці фалопієвих труб у пошуку яйцеклітини Проникнення сперматозоїду до яцеклітини крізь її
- 14. Повздовжній розріз крізь головку сперматозоїда. Генетичний матеріал міститься у голівці +8 днів. Вік ембріону — одна
- 15. +22 дні. Розвиток ембріону продовжуєетсья, і можна побачити утворення нервової трубки +24 дні. Один із найголовніших
- 16. +9 тижнів. Судини на голові розвинулись у вигляді літери V ; починається розвиток кісток черепу +
- 17. + 20 тижнів. Зародок вже розміром біля 20 сантиметрів. + 36 тижнів.
- 18. Насіння квіткових рослин
- 19. Явище взаємодії частин зародка, що розвивається, та його біологічне значення За нормальних умов розвитку окремі частини
- 20. Післяембріональний (постембріональний) розвиток, його типи і етапи у тварин Постембріональний (постнатальний) період може бути прямим і
- 21. У розвитку з повним перетворенням личинка різко відрізняється від дорослої тварини. Вона має личинкові (провізорні) органи,
- 22. Розвиток з неповним перетворенням спостерігається у представників прямокрилих, тарганів, бабок тощо. За неповного перетворення в комах,
- 23. Ріст і його типи Ріст — невід'ємна властивість живого. Він полягає у збільшенні кількості клітин та
- 24. Явище регенерації і його біологічне значення У дорослому організмі тривають процеси розвитку, що пов'язані з поділом
- 25. Життєвий цикл Цикл розвитку — сукупність стадій розвитку організму між певним етапом його життя й тим
- 26. Приклади складних життєвих циклів попелиць та аурелії: а — життєвий цикл попелиць: чергування крилатих і безкрилих
- 27. Чергування різних поколінь у життєвому циклі та його біологічне значення У життєвому циклі багатьох водоростей і
- 28. Способи відтворення У тваринному світі відомо три способи відтворення потомства: яйцеродіння, яйцеживородіння та живородіння. Яйцеродінням називається
- 29. ТКАНИНИ РОСЛИННОГО ОРГАНІЗМУ Тканини — стійкі комплекси клітин, подібні за походженням, будовою та пристосовані до виконання
- 30. Провідні тканини забезпечують транспорт речовин між надземними та підземними частинами рослини, а також здійснюють взаємозв'язок між
- 31. Тканини рослин Покривні тканини Провідні тканини Механічні тканини
- 32. Поперечний розріз крізь пластинку листа
- 33. Передумови утворення тканин у процесі еволюції У процесі еволюції утворення тканин у рослин було зумовлене переходом
- 34. Тканини людини та тварин В організмі людини та тварин виділяють чотири типи тканин: епітеліальну, сполучну, м'язову
- 35. Епітеліальна тканина Епітеліальна тканина вкриває поверхню тіла вистилає слизові оболонки, відокремлюючи ор ганізм від зовнішнього середовища.
- 36. Епітеліальні тканини
- 37. Сполучна тканина Сполучна тканина виконує різноманітні функції: опорну, захисну, живильну (трофічну), транспортну, запасаючу. Виділяють такі типи
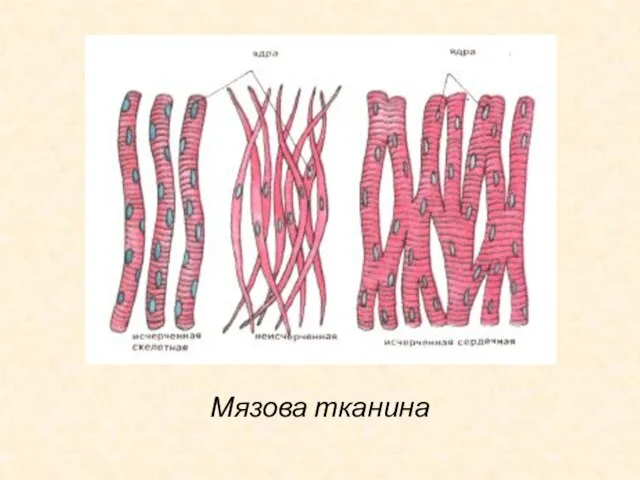
- 38. М'язова тканина М'язова тканина виконує рухову (моторну) функцію. Клітини м'язової тканини називають міоцитами. У цитоплазмі міоцитів
- 39. Мязова тканина
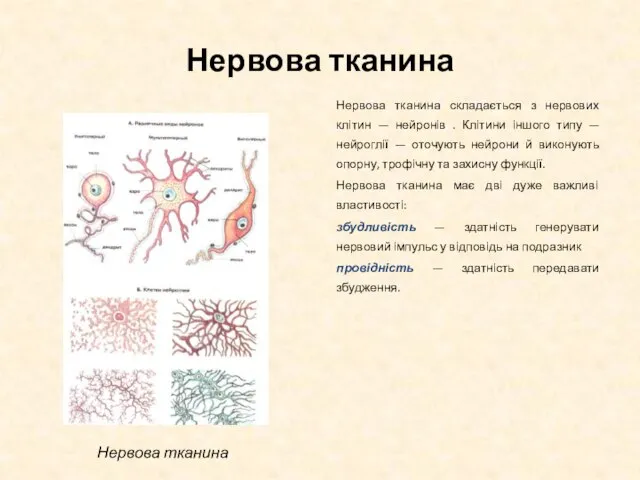
- 40. Нервова тканина Нервова тканина складається з нервових клітин — нейронів . Клітини іншого типу — нейроглії
- 42. Скачать презентацию
Слайд 2Етапи індивідуального розвитку організмів
Індивідуальний розвиток організму від його зародження до смерті називається
Етапи індивідуального розвитку організмів
Індивідуальний розвиток організму від його зародження до смерті називається

Слайд 3Зародковий (ембріональний) період розвитку
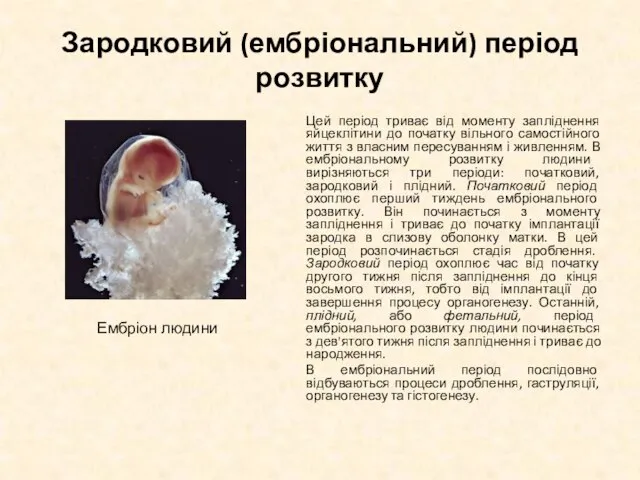
Цей період триває від моменту запліднення яйцеклітини до початку
Зародковий (ембріональний) період розвитку
Цей період триває від моменту запліднення яйцеклітини до початку
Слайд 4Дроблення та утворення бластули (на прикладі ссавців)
Відразу після запліднення починається процес
Дроблення та утворення бластули (на прикладі ссавців)
Відразу після запліднення починається процес

Слайд 5Типи дроблення яйцеклітин:
а — повне рівномірне ізолецитальної яйцеклітини; б — повне нерівномірне
Типи дроблення яйцеклітин:
а — повне рівномірне ізолецитальної яйцеклітини; б — повне нерівномірне

Слайд 6Утворення гаструли
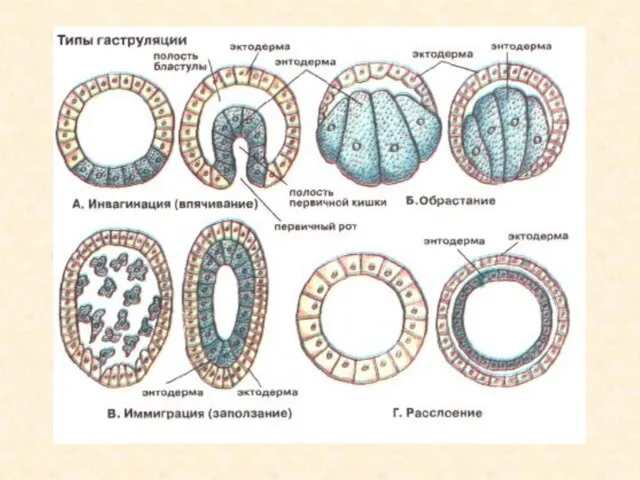
Сукупність процесів, які забезпечують утворення гаструли, називають гаструляцією. При цьому із
Утворення гаструли
Сукупність процесів, які забезпечують утворення гаструли, називають гаструляцією. При цьому із

Слайд 8Стадія утворення трьох зародкових листків
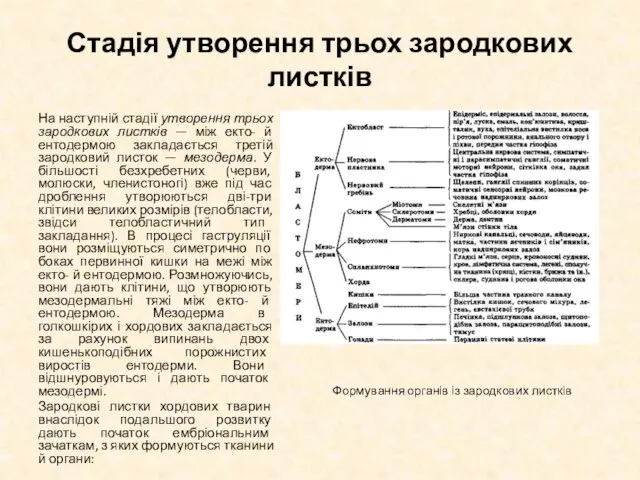
На наступній стадії утворення трьох зародкових листків —
Стадія утворення трьох зародкових листків
На наступній стадії утворення трьох зародкових листків —
Слайд 9Типи утворення мезодерми
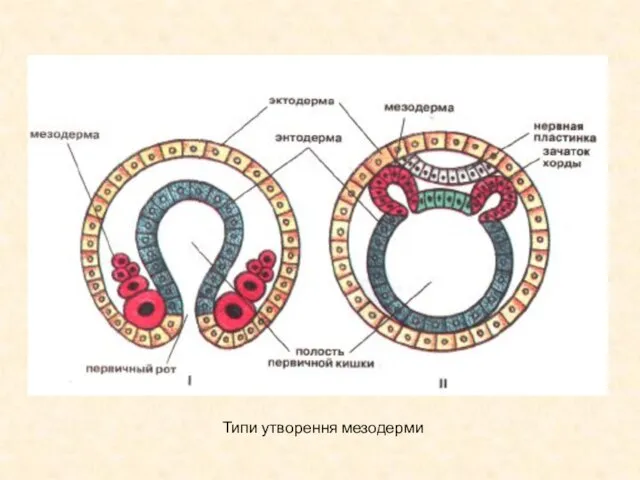
Типи утворення мезодерми
Слайд 10Диференціація тканин і органів під час зародкового розвитку (процес гістогенезу та органогенезу)
В
Диференціація тканин і органів під час зародкового розвитку (процес гістогенезу та органогенезу)
В

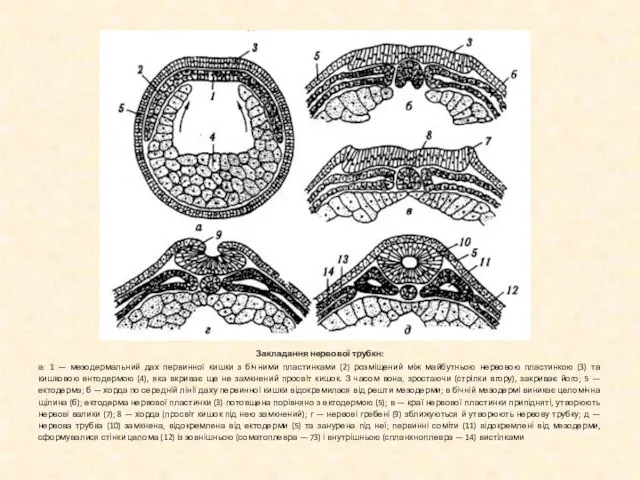
Слайд 11Закладання нервової трубкн:
а: 1 — мезодермальний дах первинної кишки з бічними пластинками
Закладання нервової трубкн:
а: 1 — мезодермальний дах первинної кишки з бічними пластинками
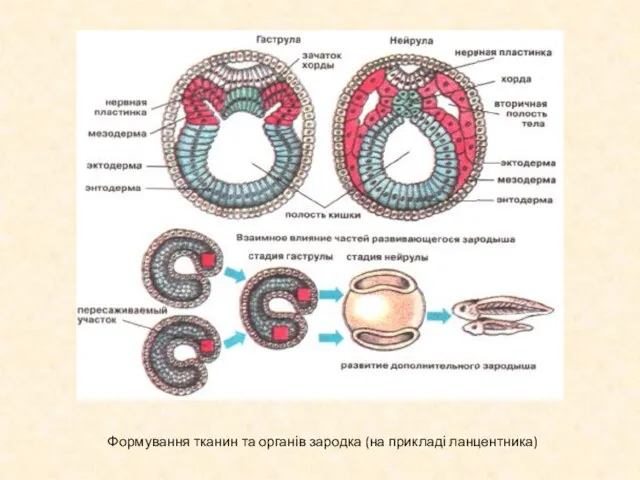
Слайд 12Формування тканин та органів зародка (на прикладі ланцентника)
Формування тканин та органів зародка (на прикладі ланцентника)
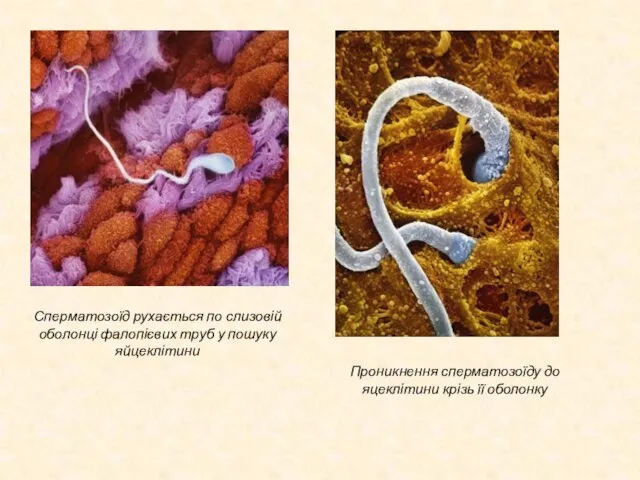
Слайд 13Сперматозоїд рухається по слизовій оболонці фалопієвих труб у пошуку яйцеклітини
Проникнення сперматозоїду до
Сперматозоїд рухається по слизовій оболонці фалопієвих труб у пошуку яйцеклітини
Проникнення сперматозоїду до
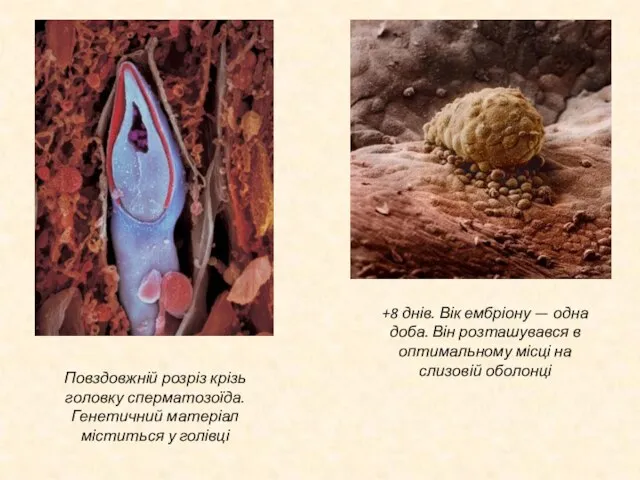
Слайд 14Повздовжній розріз крізь головку сперматозоїда. Генетичний матеріал міститься у голівці
+8 днів. Вік
Повздовжній розріз крізь головку сперматозоїда. Генетичний матеріал міститься у голівці
+8 днів. Вік
Слайд 15+22 дні. Розвиток ембріону продовжуєетсья, і можна побачити утворення нервової трубки
+24 дні.
+22 дні. Розвиток ембріону продовжуєетсья, і можна побачити утворення нервової трубки
+24 дні.
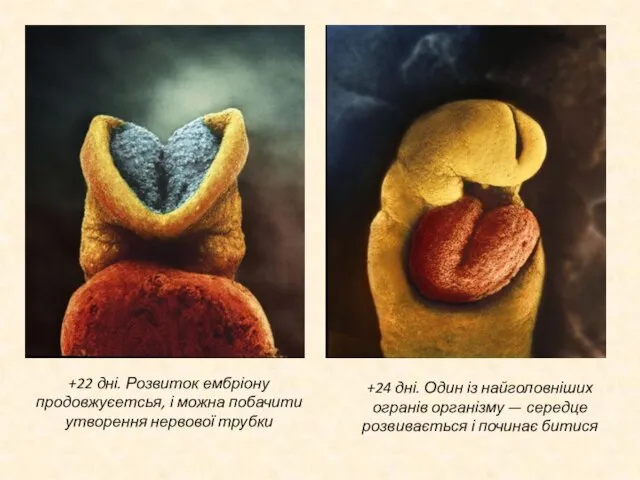
Слайд 16+9 тижнів. Судини на голові розвинулись у вигляді літери V ; починається
+9 тижнів. Судини на голові розвинулись у вигляді літери V ; починається
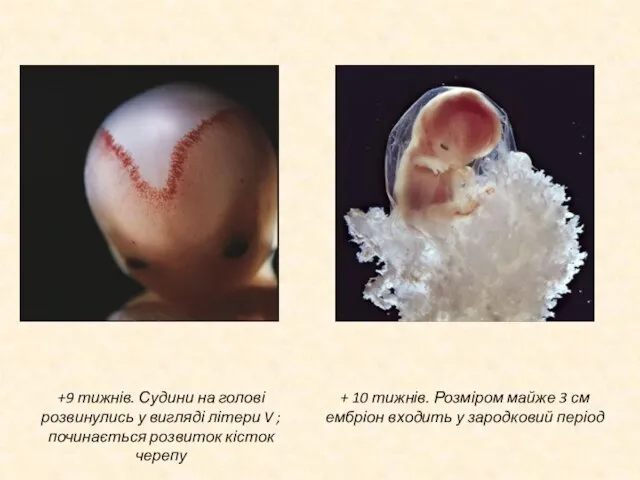
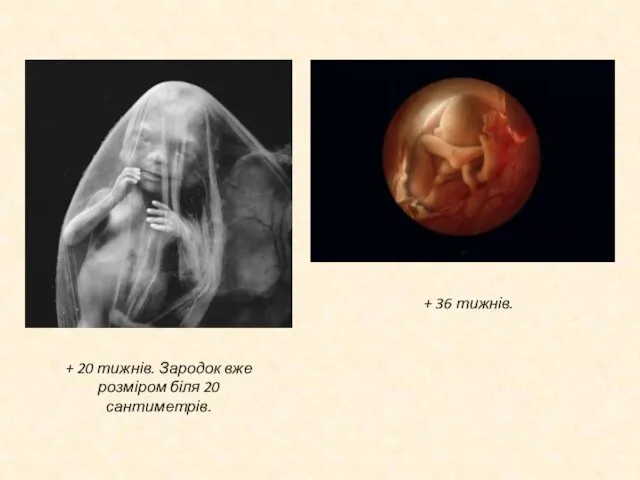
Слайд 17+ 20 тижнів. Зародок вже розміром біля 20 сантиметрів.
+ 36 тижнів.
+ 20 тижнів. Зародок вже розміром біля 20 сантиметрів.
+ 36 тижнів.
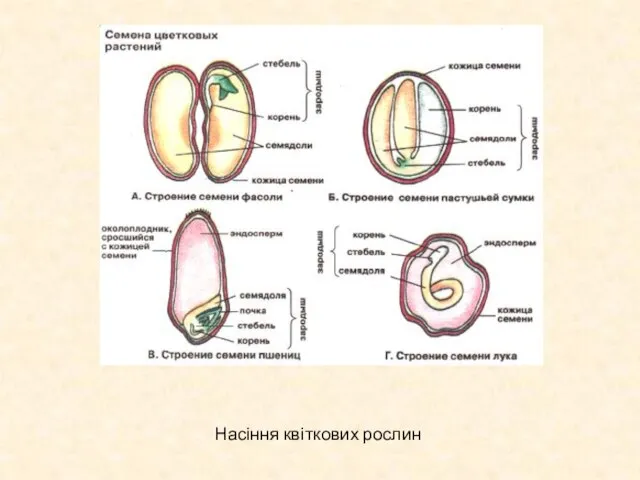
Слайд 18Насіння квіткових рослин
Насіння квіткових рослин
Слайд 19Явище взаємодії частин зародка, що розвивається, та його біологічне значення
За нормальних умов
Явище взаємодії частин зародка, що розвивається, та його біологічне значення
За нормальних умов

Слайд 20Післяембріональний (постембріональний) розвиток, його типи і етапи у тварин
Постембріональний (постнатальний) період може
Післяембріональний (постембріональний) розвиток, його типи і етапи у тварин
Постембріональний (постнатальний) період може

Слайд 21У розвитку з повним перетворенням личинка різко відрізняється від дорослої тварини. Вона
У розвитку з повним перетворенням личинка різко відрізняється від дорослої тварини. Вона

Слайд 22Розвиток з неповним перетворенням спостерігається у представників прямокрилих, тарганів, бабок тощо. За
Розвиток з неповним перетворенням спостерігається у представників прямокрилих, тарганів, бабок тощо. За

Слайд 23Ріст і його типи
Ріст — невід'ємна властивість живого. Він полягає у
Ріст і його типи
Ріст — невід'ємна властивість живого. Він полягає у

Слайд 24Явище регенерації і його біологічне значення
У дорослому організмі тривають процеси розвитку, що
Явище регенерації і його біологічне значення
У дорослому організмі тривають процеси розвитку, що

Слайд 25Життєвий цикл
Цикл розвитку — сукупність стадій розвитку організму між певним етапом його
Життєвий цикл
Цикл розвитку — сукупність стадій розвитку організму між певним етапом його

Слайд 26Приклади складних життєвих циклів попелиць та аурелії:
а — життєвий цикл попелиць:
Приклади складних життєвих циклів попелиць та аурелії:
а — життєвий цикл попелиць:

Слайд 27Чергування різних поколінь у життєвому циклі та його біологічне значення
У життєвому циклі
Чергування різних поколінь у життєвому циклі та його біологічне значення
У життєвому циклі

Слайд 28Способи відтворення
У тваринному світі відомо три способи відтворення потомства: яйцеродіння, яйцеживородіння та
Способи відтворення
У тваринному світі відомо три способи відтворення потомства: яйцеродіння, яйцеживородіння та

Слайд 29ТКАНИНИ РОСЛИННОГО ОРГАНІЗМУ
Тканини — стійкі комплекси клітин, подібні за походженням, будовою та
ТКАНИНИ РОСЛИННОГО ОРГАНІЗМУ
Тканини — стійкі комплекси клітин, подібні за походженням, будовою та

Слайд 30Провідні тканини забезпечують транспорт речовин між надземними та підземними частинами рослини, а
Провідні тканини забезпечують транспорт речовин між надземними та підземними частинами рослини, а

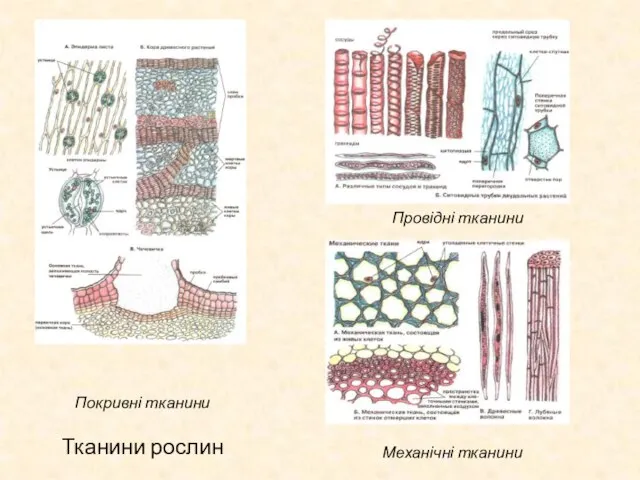
Слайд 31Тканини рослин
Покривні тканини
Провідні тканини
Механічні тканини
Тканини рослин
Покривні тканини
Провідні тканини
Механічні тканини
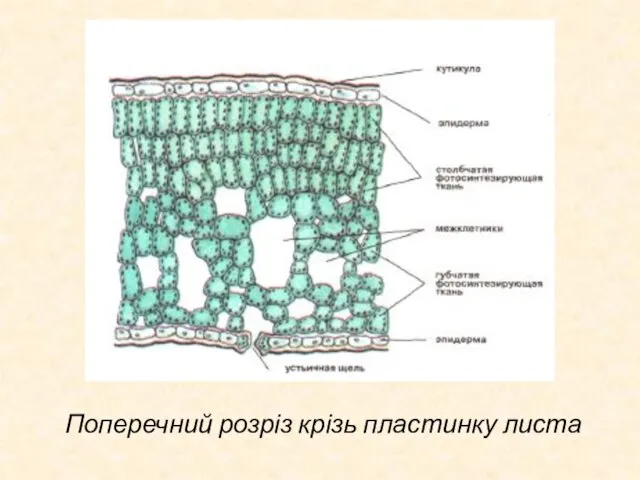
Слайд 32Поперечний розріз крізь пластинку листа
Поперечний розріз крізь пластинку листа
Слайд 33Передумови утворення тканин у процесі еволюції
У процесі еволюції утворення тканин у рослин
Передумови утворення тканин у процесі еволюції
У процесі еволюції утворення тканин у рослин

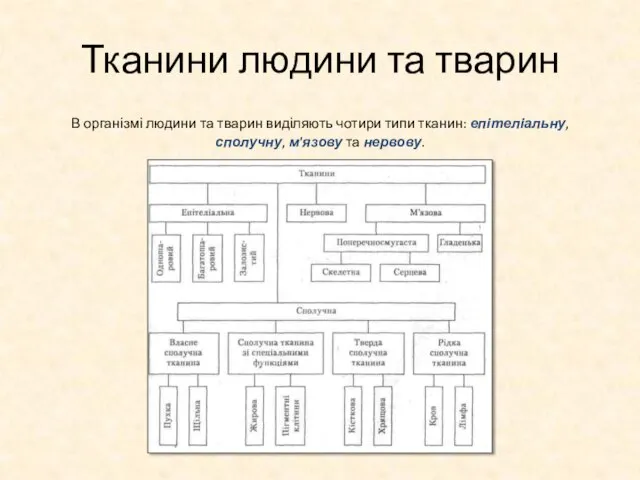
Слайд 34Тканини людини та тварин
В організмі людини та тварин виділяють чотири типи тканин:
Тканини людини та тварин
В організмі людини та тварин виділяють чотири типи тканин:
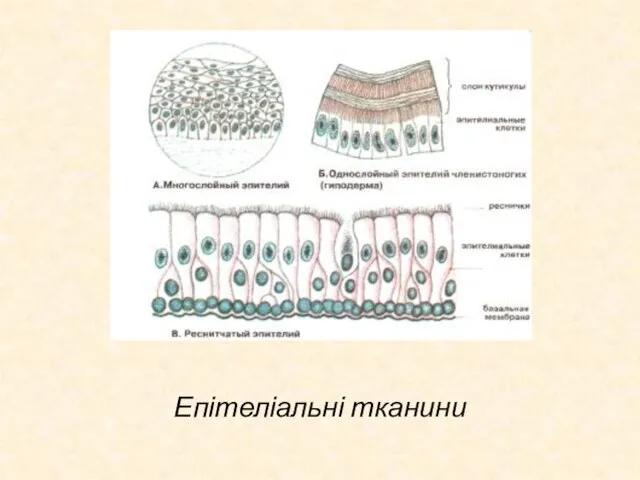
Слайд 35Епітеліальна тканина
Епітеліальна тканина вкриває поверхню тіла вистилає слизові оболонки, відокремлюючи ор ганізм
Епітеліальна тканина
Епітеліальна тканина вкриває поверхню тіла вистилає слизові оболонки, відокремлюючи ор ганізм

Слайд 36Епітеліальні тканини
Епітеліальні тканини
Слайд 37Сполучна тканина
Сполучна тканина виконує різноманітні функції: опорну, захисну, живильну (трофічну), транспортну, запасаючу.
Сполучна тканина
Сполучна тканина виконує різноманітні функції: опорну, захисну, живильну (трофічну), транспортну, запасаючу.

Слайд 38М'язова тканина
М'язова тканина виконує рухову (моторну) функцію. Клітини м'язової тканини називають міоцитами.
М'язова тканина
М'язова тканина виконує рухову (моторну) функцію. Клітини м'язової тканини називають міоцитами.

Слайд 39Мязова тканина
Мязова тканина
Слайд 40Нервова тканина
Нервова тканина складається з нервових клітин — нейронів . Клітини іншого
Нервова тканина
Нервова тканина складається з нервових клітин — нейронів . Клітини іншого
 Посмотрите, это я
Посмотрите, это я Обобщение по теме: «Однородные члены предложения»
Обобщение по теме: «Однородные члены предложения» Научное обеспечение развития стратегии пищевой и перерабатывающей промышленности
Научное обеспечение развития стратегии пищевой и перерабатывающей промышленности Трудовое право. Формы регулирования труда работников. Система и особенности источников трудового права
Трудовое право. Формы регулирования труда работников. Система и особенности источников трудового права ИСТОРИЯ ЧАСОВ
ИСТОРИЯ ЧАСОВ Что такое кротость?
Что такое кротость? Генрих Герц
Генрих Герц Эрмитаж. Новая концепция интерфейса платформы «1С-Битрикс 9.5»
Эрмитаж. Новая концепция интерфейса платформы «1С-Битрикс 9.5» Дифференциальная диагностика нейроинфекций
Дифференциальная диагностика нейроинфекций Современная терапия артериальной гипертензии
Современная терапия артериальной гипертензии  Опишите картинку
Опишите картинку Географические координаты Долгота и широта
Географические координаты Долгота и широта Комплексное решение для автоматизации предприятий сферы ЖКХ
Комплексное решение для автоматизации предприятий сферы ЖКХ 3 сыйныфта татар теле дәресе
3 сыйныфта татар теле дәресе Совершенствование воспитательной системы в школах Пензенской области
Совершенствование воспитательной системы в школах Пензенской области Природни забележителности и резервати в Предбалкана
Природни забележителности и резервати в Предбалкана Видеофильм о Балтийске предоставлен авторомБекиш Евгением Александровичем
Видеофильм о Балтийске предоставлен авторомБекиш Евгением Александровичем Юрий Зиссер, генеральный директор УП «Надежные программы» (TUT.BY, hoster.by), зампредседателя ОО «Белинфоком»
Юрий Зиссер, генеральный директор УП «Надежные программы» (TUT.BY, hoster.by), зампредседателя ОО «Белинфоком» Изменение поведения. Мотивация. Мотивационное интервью
Изменение поведения. Мотивация. Мотивационное интервью Казахстан в годы войны против фашистских захватчиков (1941-1945)
Казахстан в годы войны против фашистских захватчиков (1941-1945) Презентация на тему Свободное падение тел
Презентация на тему Свободное падение тел  Одарённость как педагогический и психологический феномен
Одарённость как педагогический и психологический феномен Амфибии
Амфибии Превращение энергии (3 класс)
Превращение энергии (3 класс) A working day
A working day Деловая игра: «Роль электронных образовательных ресурсов в реализации мыслительного подхода»
Деловая игра: «Роль электронных образовательных ресурсов в реализации мыслительного подхода» Британия и ее колонии
Британия и ее колонии Режим дня и его влияние на здоровье ребёнка и качество его обучения Хорошими людьми становятся больше от упражнения, чем от при
Режим дня и его влияние на здоровье ребёнка и качество его обучения Хорошими людьми становятся больше от упражнения, чем от при