- Макроэволюция

Содержание
- 2. Макроэволюция: разметка участка Крайние точки, оконтуривающие круг макроэволюционных процессов: Араморфоз - идиоадаптация Дивергенция – параллелизм -
- 3. Макроэволюция - разметка участка Изучение палеонтологических находок и современных форм позволяет в настоящее время считать достаточно
- 4. Макроэволюция - разметка участка Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы, (позволяя расширить старую
- 5. Макроэволюция - разметка участка А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому прогрессу, ароморфозами. Согласно
- 6. Макроэволюция - разметка участка Верхняя и нижняя челюсти и зубы различных змей 1 – питон, 2,7
- 7. Макроэволюция - разметка участка Миксоспоридии – пример катагенеза Генетический анализ показал, что миксоспоридии, всегда считавшиеся «хорошим»
- 8. Макроэволюция - разметка участка Дивергенция (от средневекового лат. divergo — отклоняюсь) Росомаха питается преимущес-твенно падалью. Широкие
- 9. Макроэволюция - разметка участка Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а конвергенция – это
- 10. Макроэволюция - разметка участка Параллелизм – конвергенция гомологичных органов Поза «стояния столбиком» и характерное для грызунов
- 11. Макроэволюция - разметка участка Термин параллелизм употребляется также не для характеристи-ки изменения органов, а для указания
- 12. Макроэволюция - разметка участка Полифилия – объединение в один таксон разных по происхождению групп. Поли- много,
- 13. Макроэволюция – современные сложности Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия и полифилия на
- 14. Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по числу аминокислотных замен (Каа)
- 15. Макроэволюция – современные сложности К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов анализа филогении практически
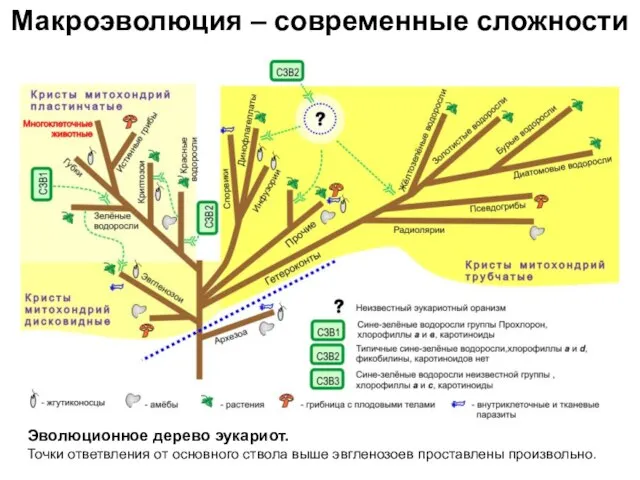
- 16. Макроэволюция – современные сложности Эволюционное дерево эукариот. Точки ответвления от основного ствола выше эвгленозоев проставлены произвольно.
- 17. Горизонтальный перенос генов – к чему это приводит Прокариоты представлены двумя царствами – архебактерии и эубактерии
- 18. Биологический прогресс: противоречие между сложностью и успехом А.Н. Северцов снимает его, разделяя биологический и морфо-физиологический прогресс
- 19. Основная философская проблема макроэволюции - направленность Ранние эволюционисты объясняли развитие природы через конечные причины, стремлением к
- 21. Скачать презентацию
Слайд 2Макроэволюция: разметка участка
Крайние точки, оконтуривающие круг макроэволюционных процессов:
Араморфоз - идиоадаптация
Дивергенция – параллелизм
Макроэволюция: разметка участка
Крайние точки, оконтуривающие круг макроэволюционных процессов:
Араморфоз - идиоадаптация
Дивергенция – параллелизм

Слайд 3Макроэволюция - разметка участка
Изучение палеонтологических находок и современных форм позволяет в настоящее
Макроэволюция - разметка участка
Изучение палеонтологических находок и современных форм позволяет в настоящее

Слайд 4Макроэволюция - разметка участка
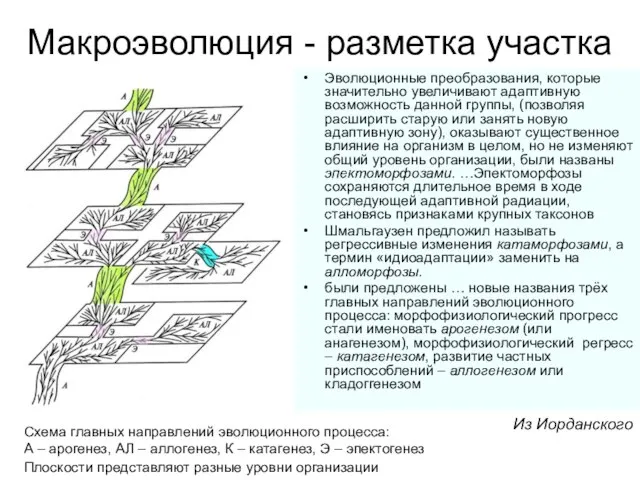
Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы,
Макроэволюция - разметка участка
Эволюционные преобразования, которые значительно увеличивают адаптивную возможность данной группы,
Слайд 5Макроэволюция - разметка участка
А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому
Макроэволюция - разметка участка
А. Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому

Слайд 6Макроэволюция - разметка участка
Верхняя и нижняя челюсти и зубы различных змей
1 –
Макроэволюция - разметка участка
Верхняя и нижняя челюсти и зубы различных змей
1 –

Слайд 7Макроэволюция - разметка участка
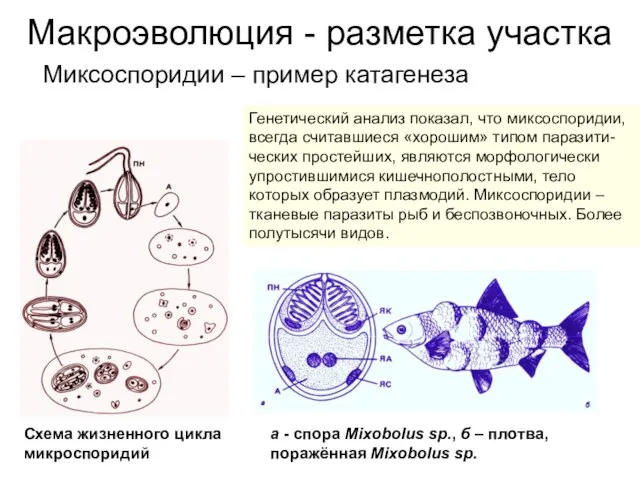
Миксоспоридии – пример катагенеза
Генетический анализ показал, что миксоспоридии, всегда
Макроэволюция - разметка участка
Миксоспоридии – пример катагенеза
Генетический анализ показал, что миксоспоридии, всегда
Слайд 8Макроэволюция - разметка участка
Дивергенция (от средневекового лат. divergo — отклоняюсь)
Росомаха питается преимущес-твенно
Макроэволюция - разметка участка
Дивергенция (от средневекового лат. divergo — отклоняюсь)
Росомаха питается преимущес-твенно

Слайд 9Макроэволюция - разметка участка
Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а
Макроэволюция - разметка участка
Конвергенция не есть полная противо-положность дивергенции. Дивергируют таксоны, а

Слайд 10Макроэволюция - разметка участка
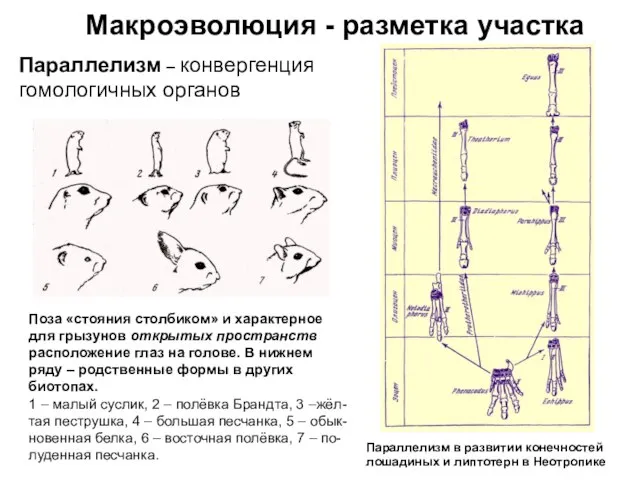
Параллелизм – конвергенция гомологичных органов
Поза «стояния столбиком» и характерное
Макроэволюция - разметка участка
Параллелизм – конвергенция гомологичных органов
Поза «стояния столбиком» и характерное
Слайд 11Макроэволюция - разметка участка
Термин параллелизм употребляется также не для характеристи-ки изменения органов,
Макроэволюция - разметка участка
Термин параллелизм употребляется также не для характеристи-ки изменения органов,

Слайд 12Макроэволюция - разметка участка
Полифилия – объединение в один таксон разных по происхождению
Макроэволюция - разметка участка
Полифилия – объединение в один таксон разных по происхождению

Слайд 13Макроэволюция – современные сложности
Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия
Макроэволюция – современные сложности
Нейтралистская теория эволюции – нейтральные признаки только дивергируют. Парафилия

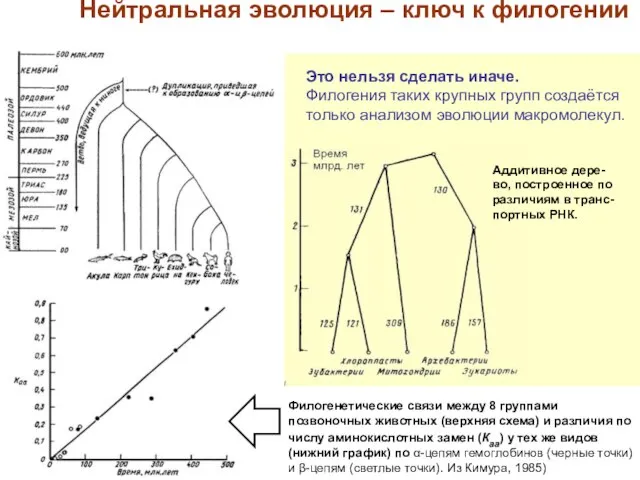
Слайд 14Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по
Филогенетические связи между 8 группами позвоночных животных (верхняя схема) и различия по
Слайд 15Макроэволюция – современные сложности
К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов
Макроэволюция – современные сложности
К настоящему времени возможности морфологического, эмбриологического и палеонтологического методов

Слайд 16Макроэволюция – современные сложности
Эволюционное дерево эукариот.
Точки ответвления от основного ствола выше
Макроэволюция – современные сложности
Эволюционное дерево эукариот. Точки ответвления от основного ствола выше
Слайд 17Горизонтальный перенос генов – к чему это приводит
Прокариоты представлены двумя царствами –
Горизонтальный перенос генов – к чему это приводит
Прокариоты представлены двумя царствами –

Слайд 18Биологический прогресс: противоречие между сложностью и успехом
А.Н. Северцов снимает его, разделяя биологический
Биологический прогресс: противоречие между сложностью и успехом
А.Н. Северцов снимает его, разделяя биологический

Слайд 19Основная философская проблема макроэволюции - направленность
Ранние эволюционисты объясняли развитие природы через конечные
Основная философская проблема макроэволюции - направленность
Ранние эволюционисты объясняли развитие природы через конечные

 Действие норм права во времени, в пространстве и по кругу лиц
Действие норм права во времени, в пространстве и по кругу лиц Решение показательных неравенств
Решение показательных неравенств Международные переговоры. Специфика переговорного процесса в различных условиях
Международные переговоры. Специфика переговорного процесса в различных условиях Бизнес-план: производство духов в Хабаровске
Бизнес-план: производство духов в Хабаровске Система и отрасли права
Система и отрасли права Презентация на тему Иван Калита внук Александра Невского
Презентация на тему Иван Калита внук Александра Невского  Институт будущего
Институт будущего Школьный компьютер в 21 веке Проектная работа учеников 5 класса ГОУ ЦО № 1474, Учитель Головкин Ю. В.
Школьный компьютер в 21 веке Проектная работа учеников 5 класса ГОУ ЦО № 1474, Учитель Головкин Ю. В. Intelli Corder
Intelli Corder Экспертный отдел продаж
Экспертный отдел продаж Уставный капитал 480 млн. рублей ОАО «Камский Индустриальный парк «Мастер» основано 29 июля 2004 года в городе Набережные Челны на базе
Уставный капитал 480 млн. рублей ОАО «Камский Индустриальный парк «Мастер» основано 29 июля 2004 года в городе Набережные Челны на базе Центр дополнительного профессионального образования ГБОУВО РК КИПУ
Центр дополнительного профессионального образования ГБОУВО РК КИПУ Что такое оценка?
Что такое оценка? Пропиточные лаки, эмали и компаунды
Пропиточные лаки, эмали и компаунды Семантические особенности фразеологизмов-зоонимов с компонентами лошадь, корова в якутском и бурятском языках
Семантические особенности фразеологизмов-зоонимов с компонентами лошадь, корова в якутском и бурятском языках ШКОЛЬНЫЙ ЭТАПКОНКУРСА« УЧИТЕЛЬ ГОДА-2010»
ШКОЛЬНЫЙ ЭТАПКОНКУРСА« УЧИТЕЛЬ ГОДА-2010» Особенности формирования экономики знаний в современных условиях
Особенности формирования экономики знаний в современных условиях Ситуация на рынке жилья Санкт-Петербурга в 2008 году. Прогноз на 2009.
Ситуация на рынке жилья Санкт-Петербурга в 2008 году. Прогноз на 2009. Живопись в начале xx века
Живопись в начале xx века Государственное регулирование охраны труда
Государственное регулирование охраны труда Синус, косинус, тангенс суммы и разности аргументов (10 класс)
Синус, косинус, тангенс суммы и разности аргументов (10 класс) История Олимпийских игр. Олимпиады Древней Греции (776 до н.э. – 394 н.э.)
История Олимпийских игр. Олимпиады Древней Греции (776 до н.э. – 394 н.э.) Bogunly gurçuklar
Bogunly gurçuklar Иван Алексеевич Бунин
Иван Алексеевич Бунин Государственное учреждение «Управление по обеспечению рационального использования и качества топливно-энергетических ресурсов
Государственное учреждение «Управление по обеспечению рационального использования и качества топливно-энергетических ресурсов  Открытие Декады филологического образования в МБОУ Гайдаровская СОШ
Открытие Декады филологического образования в МБОУ Гайдаровская СОШ Composers: Romantic period (1820 -1910)
Composers: Romantic period (1820 -1910) Степень числа с натуральным показателем
Степень числа с натуральным показателем