- Моделирование регуляции развития меристемы побега в эмбриогенезе Arabidopsis thaliana

Содержание
- 2. Объект исследования: Arabidopsis thaliana Arabidopsis был первым растением, геном которого был полностью секвенирован. Существует программа, по
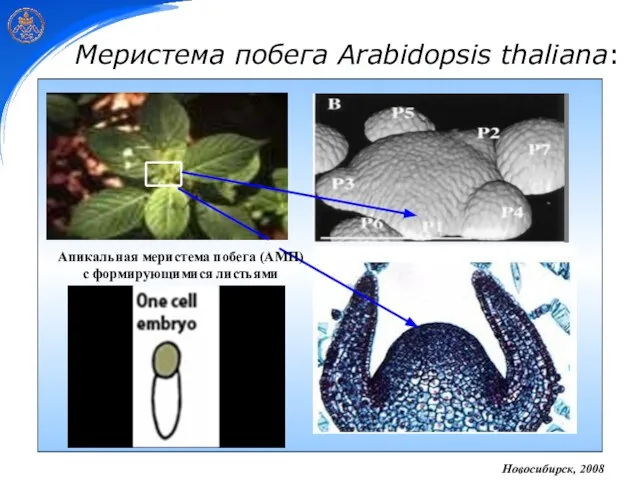
- 3. Меристема побега Arabidopsis thaliana: Новосибирск, 2008 Апикальная меристема побега (АМП) с формирующимися листьями
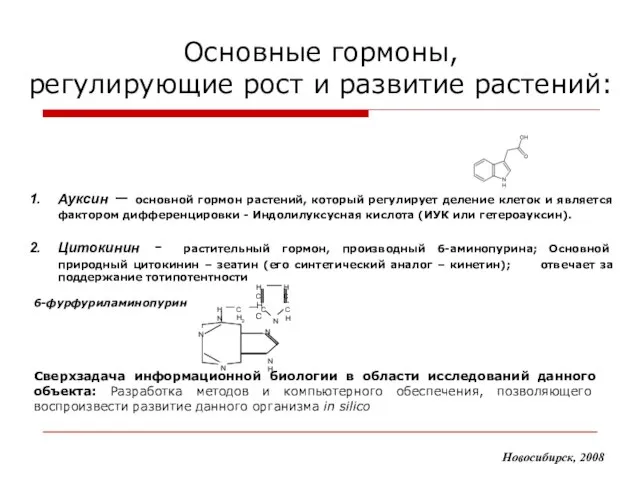
- 4. Основные гормоны, регулирующие рост и развитие растений: Новосибирск, 2008 Ауксин – основной гормон растений, который регулирует
- 5. Новосибирск, 2008 Цель: Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при развитии меристемы побега
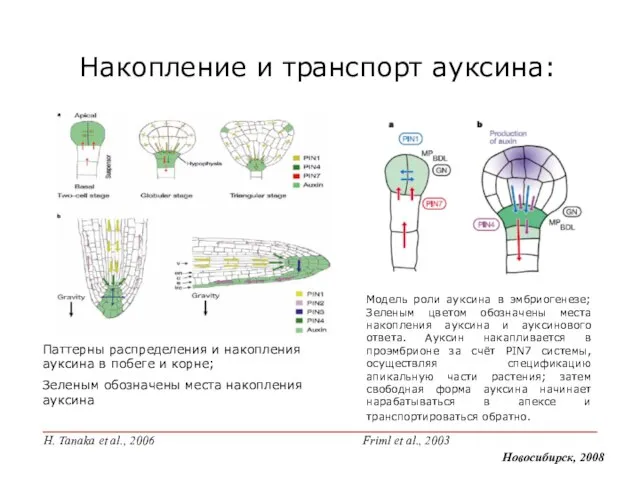
- 6. Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина и ауксинового ответа. Ауксин накапливается
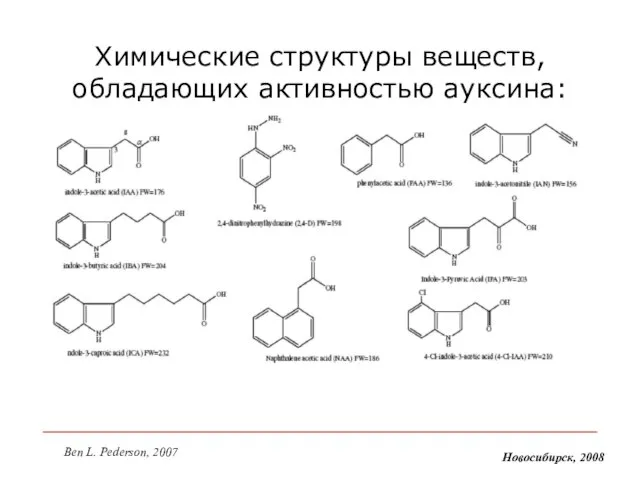
- 7. Химические структуры веществ, обладающих активностью ауксина: Новосибирск, 2008 Ben L. Pederson, 2007
- 8. Метаболизм ауксина: Новосибирск, 2008 Woodward and Bartel, 2005 Потенциальные пути биосинтеза ауксина в клетке Арабидопсиса. Пути
- 9. Конъюгация ауксина: Новосибирск, 2008 Потенциальные пути метаболизма ауксина в клетке Арабидопсиса. Woodward and Bartel, 2005
- 10. Структурная модель генной сети метаболизма ауксина*: Новосибирск, 2008 *-Ananko et al., 2005
- 11. *цитоплазма *ядро *клеточная мембрана *пероксисома *ЭПР *хлоропласт *митохондрия Новосибирск, 2008 Структурная модель генной сети метаболизма ауксина:
- 12. Новосибирск, 2008 Структурная модель генной сети метаболизма ауксина:
- 13. Новосибирск, 2008 Редуцированная структурная модель генной сети метаболизма ауксина:
- 14. MGSgenerator: блок автоматической конвертации формата ГС в формат моделей: Новосибирск, 2008
- 15. MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем: Новосибирск, 2008
- 16. Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега: В клетках побега Арабидопсиса, свободная
- 17. Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке в зависимости от начальной концентрации:
- 18. Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях (trp2,trp3): Расчёты модели качественно совпадают
- 19. Эмбриональное развитие меристемы побега Arabidopsis Thaliana Область моделирования Новосибирск, 2008
- 20. Разработка пространственно – распределённой модели: Основные принципы: Клетки автомата могут обмениваться химическими сигналами. Было выбрана 3
- 21. Типы клеток автомата: Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS и находятся в верхней
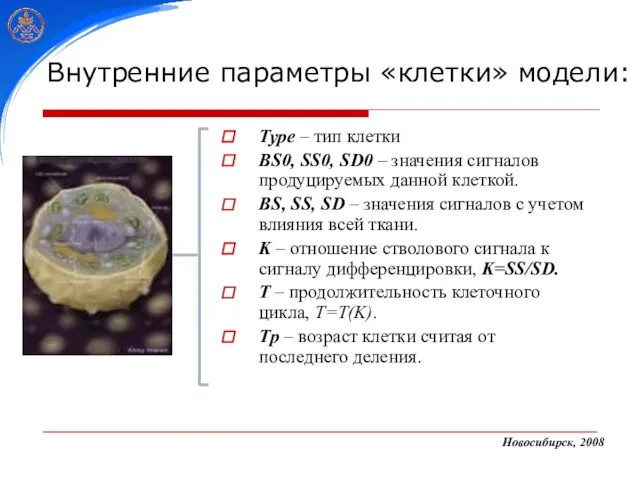
- 22. Type – тип клетки BS0, SS0, SD0 – значения сигналов продуцируемых данной клеткой. BS, SS, SD
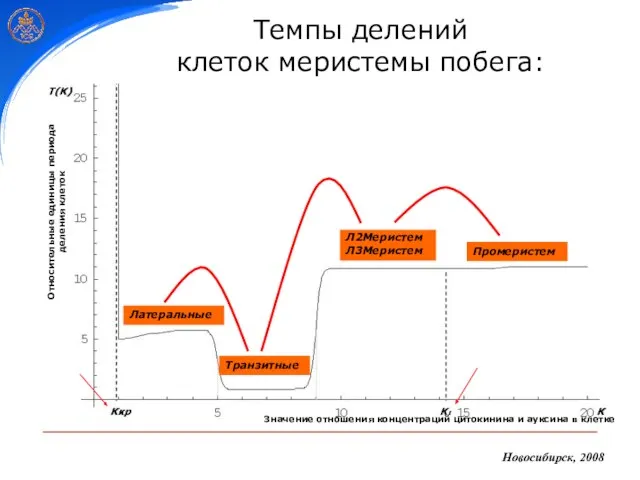
- 23. Новосибирск, 2008 Темпы делений клеток меристемы побега: Промеристем Л2Меристем Л3Меристем Транзитные Латеральные
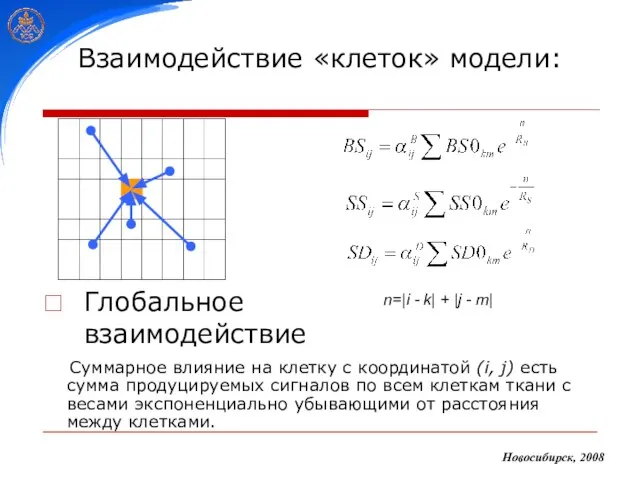
- 24. Взаимодействие «клеток» модели: Глобальное взаимодействие Новосибирск, 2008 Суммарное влияние на клетку с координатой (i, j) есть
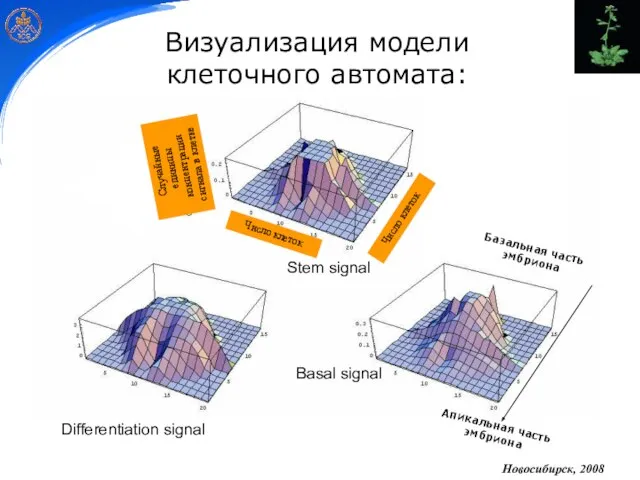
- 25. Stem signal Differentiation signal Basal signal Визуализация модели клеточного автомата: Новосибирск, 2008
- 26. Результаты моделирования. Нормальное развитие зародыша: Новосибирск, 2008
- 27. Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and Floral Meristem Cell Proliferation and Fate.
- 28. Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки): В развивающейся АМП Нормальное развитие
- 29. Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки): Нормальное развитие Мутация II типа
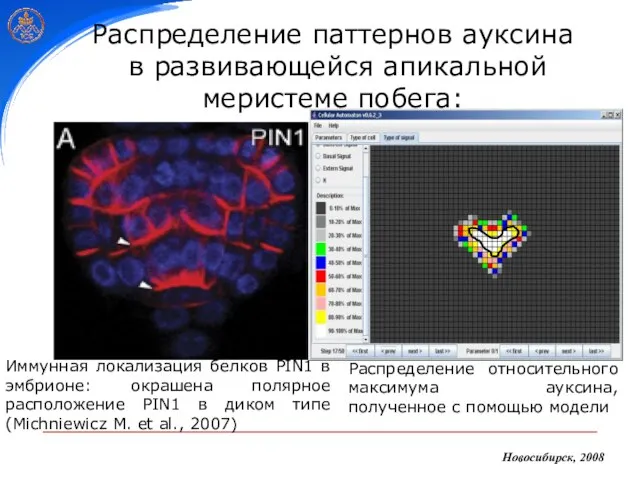
- 30. Распределение паттернов ауксина в развивающейся апикальной меристеме побега: Иммунная локализация белков PIN1 в эмбрионе: окрашена полярное
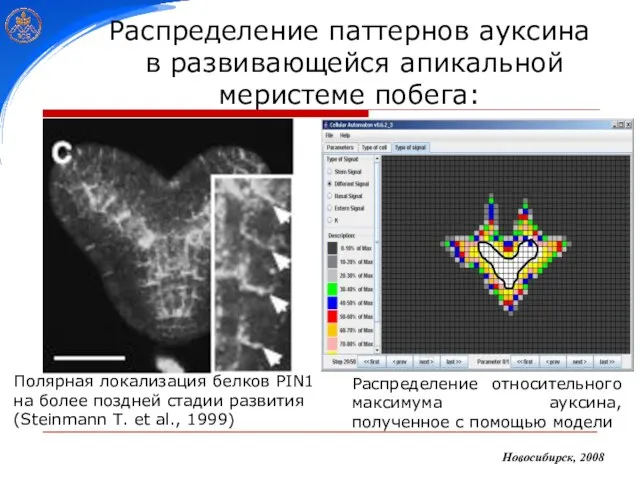
- 31. Распределение паттернов ауксина в развивающейся апикальной меристеме побега: Полярная локализация белков PIN1 на более поздней стадии
- 32. Новосибирск, 2008 Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие этапы: 1) автоматическая генерация
- 33. Новосибирск, 2008 Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма ауксина дополнена моделями, описывающими
- 34. Новосибирск, 2008 Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana. Для пространственно-распределённой модели подобран оптимальный
- 36. Скачать презентацию
Слайд 2Объект исследования:
Arabidopsis thaliana
Arabidopsis был первым растением, геном которого был полностью секвенирован.
Объект исследования:
Arabidopsis thaliana
Arabidopsis был первым растением, геном которого был полностью секвенирован.

Слайд 3Меристема побега Arabidopsis thaliana:
Новосибирск, 2008
Апикальная меристема побега (АМП)
с формирующимися листьями
Меристема побега Arabidopsis thaliana:
Новосибирск, 2008
Апикальная меристема побега (АМП)
с формирующимися листьями
Слайд 4Основные гормоны,
регулирующие рост и развитие растений:
Новосибирск, 2008
Ауксин – основной гормон растений,
Основные гормоны,
регулирующие рост и развитие растений:
Новосибирск, 2008
Ауксин – основной гормон растений,
Слайд 5Новосибирск, 2008
Цель:
Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при развитии
Новосибирск, 2008
Цель:
Теоретический анализ регуляторных механизмов поддержания тотипотентности и дифференцировки клеток при развитии

Слайд 6Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина и
Модель роли ауксина в эмбриогенезе; Зеленым цветом обозначены места накопления ауксина и
Слайд 7Химические структуры веществ, обладающих активностью ауксина:
Новосибирск, 2008
Ben L. Pederson, 2007
Химические структуры веществ, обладающих активностью ауксина:
Новосибирск, 2008
Ben L. Pederson, 2007
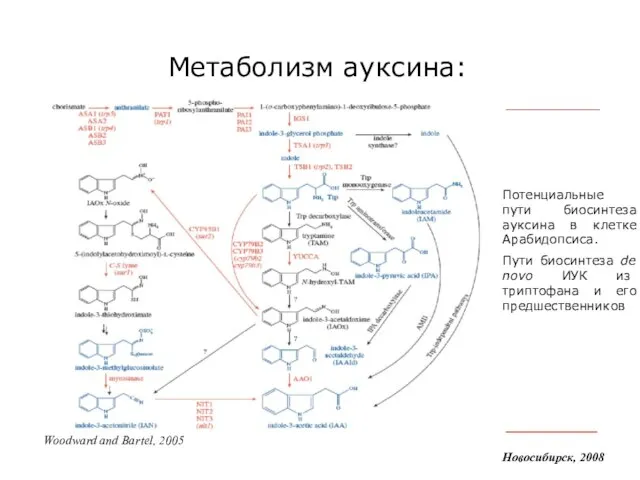
Слайд 8Метаболизм ауксина:
Новосибирск, 2008
Woodward and Bartel, 2005
Потенциальные пути биосинтеза ауксина в клетке Арабидопсиса.
Пути
Метаболизм ауксина:
Новосибирск, 2008
Woodward and Bartel, 2005
Потенциальные пути биосинтеза ауксина в клетке Арабидопсиса.
Пути
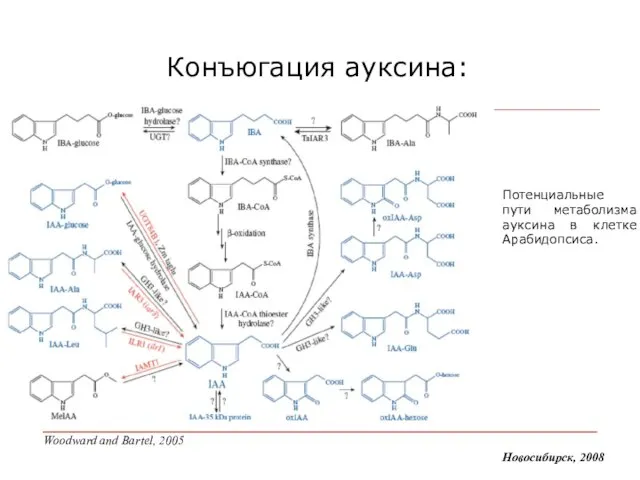
Слайд 9Конъюгация ауксина:
Новосибирск, 2008
Потенциальные пути метаболизма ауксина в клетке Арабидопсиса.
Woodward and Bartel, 2005
Конъюгация ауксина:
Новосибирск, 2008
Потенциальные пути метаболизма ауксина в клетке Арабидопсиса.
Woodward and Bartel, 2005
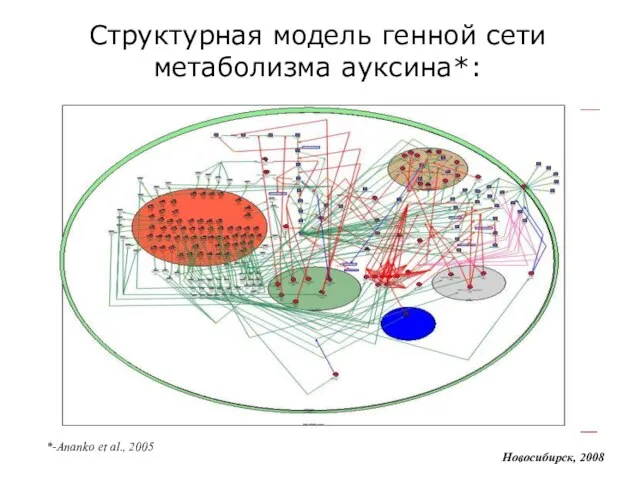
Слайд 10Структурная модель генной сети метаболизма ауксина*:
Новосибирск, 2008
*-Ananko et al., 2005
Структурная модель генной сети метаболизма ауксина*:
Новосибирск, 2008
*-Ananko et al., 2005
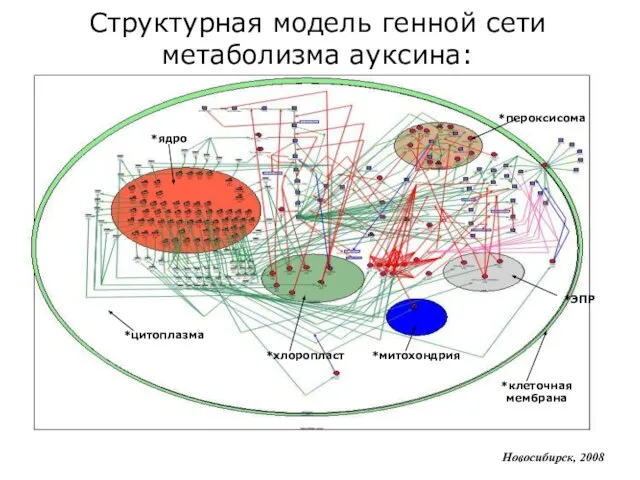
Слайд 11*цитоплазма
*ядро
*клеточная мембрана
*пероксисома
*ЭПР
*хлоропласт
*митохондрия
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
*цитоплазма
*ядро
*клеточная мембрана
*пероксисома
*ЭПР
*хлоропласт
*митохондрия
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
Слайд 12Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
Новосибирск, 2008
Структурная модель генной сети метаболизма ауксина:
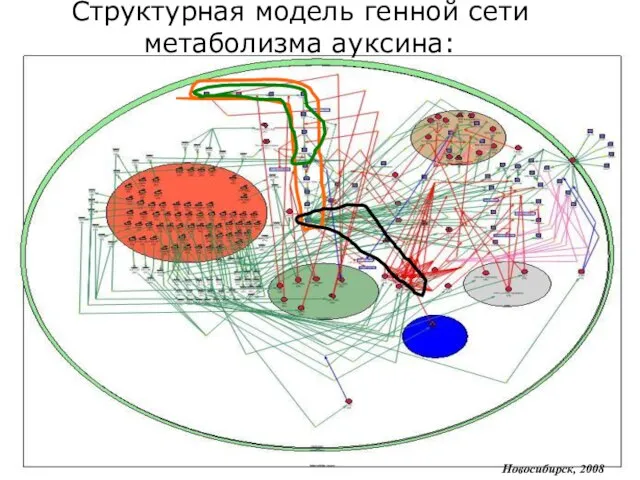
Слайд 13Новосибирск, 2008
Редуцированная структурная модель генной сети метаболизма ауксина:
Новосибирск, 2008
Редуцированная структурная модель генной сети метаболизма ауксина:
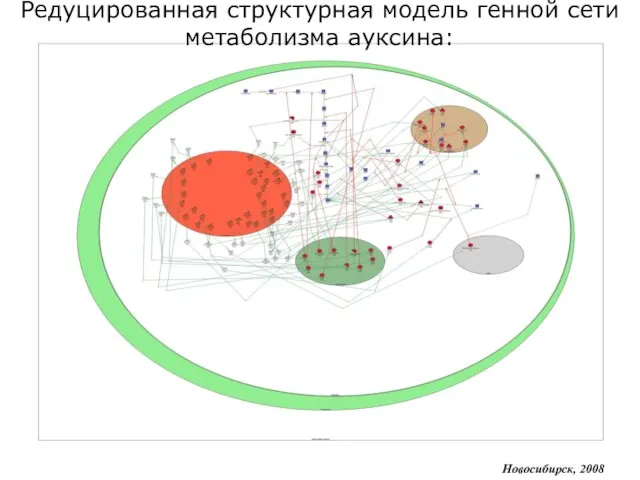
Слайд 14MGSgenerator: блок автоматической конвертации формата ГС в формат моделей:
Новосибирск, 2008
MGSgenerator: блок автоматической конвертации формата ГС в формат моделей:
Новосибирск, 2008
Слайд 15MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем:
Новосибирск, 2008
MGSmodeller: Компьютерная система для конструирования, расчета и анализа моделей молекулярно-генетических систем:
Новосибирск, 2008
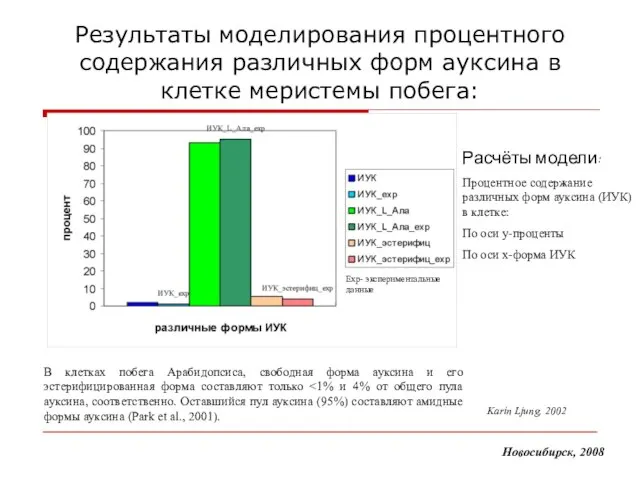
Слайд 16Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега:
В клетках
Результаты моделирования процентного содержания различных форм ауксина в клетке меристемы побега:
В клетках
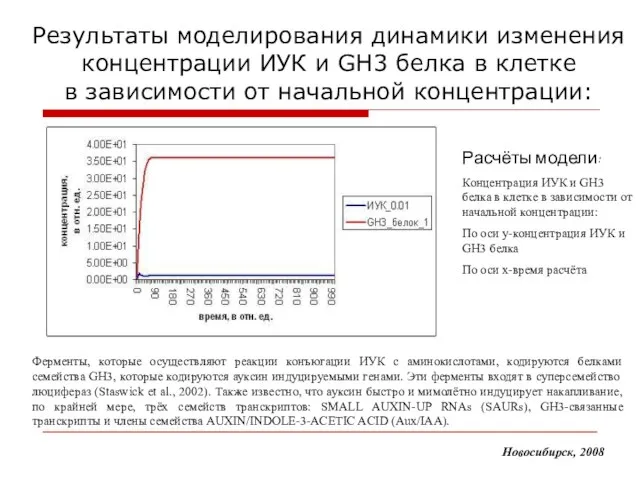
Слайд 17Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке
в
Результаты моделирования динамики изменения концентрации ИУК и GH3 белка в клетке в
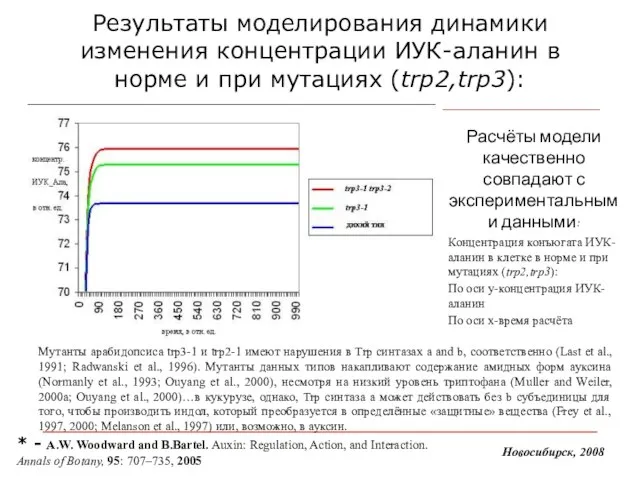
Слайд 18Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях (trp2,trp3):
Расчёты
Результаты моделирования динамики изменения концентрации ИУК-аланин в норме и при мутациях (trp2,trp3):
Расчёты
Слайд 19Эмбриональное развитие
меристемы побега Arabidopsis Thaliana
Область моделирования
Новосибирск, 2008
Эмбриональное развитие
меристемы побега Arabidopsis Thaliana
Область моделирования
Новосибирск, 2008
Слайд 20Разработка пространственно – распределённой модели:
Основные принципы:
Клетки автомата могут обмениваться химическими сигналами. Было
Разработка пространственно – распределённой модели:
Основные принципы:
Клетки автомата могут обмениваться химическими сигналами. Было

Слайд 21Типы клеток автомата:
Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS и
Типы клеток автомата:
Промеристем - клетки меристемы зародыша. Эти клетки продуцируют SS и

Слайд 22Type – тип клетки
BS0, SS0, SD0 – значения сигналов продуцируемых данной клеткой.
BS,
Type – тип клетки
BS0, SS0, SD0 – значения сигналов продуцируемых данной клеткой.
BS,
Слайд 23Новосибирск, 2008
Темпы делений
клеток меристемы побега:
Промеристем
Л2Меристем
Л3Меристем
Транзитные
Латеральные
Новосибирск, 2008
Темпы делений
клеток меристемы побега:
Промеристем
Л2Меристем
Л3Меристем
Транзитные
Латеральные
Слайд 24Взаимодействие «клеток» модели:
Глобальное взаимодействие
Новосибирск, 2008
Суммарное влияние на клетку с координатой (i,
Взаимодействие «клеток» модели:
Глобальное взаимодействие
Новосибирск, 2008
Суммарное влияние на клетку с координатой (i,
Слайд 25Stem signal
Differentiation signal
Basal signal
Визуализация модели
клеточного автомата:
Новосибирск, 2008
Stem signal
Differentiation signal
Basal signal
Визуализация модели
клеточного автомата:
Новосибирск, 2008
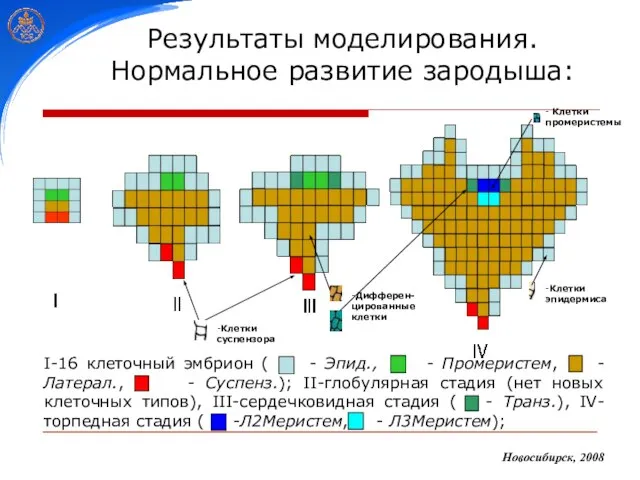
Слайд 26Результаты моделирования.
Нормальное развитие зародыша:
Новосибирск, 2008
Результаты моделирования.
Нормальное развитие зародыша:
Новосибирск, 2008
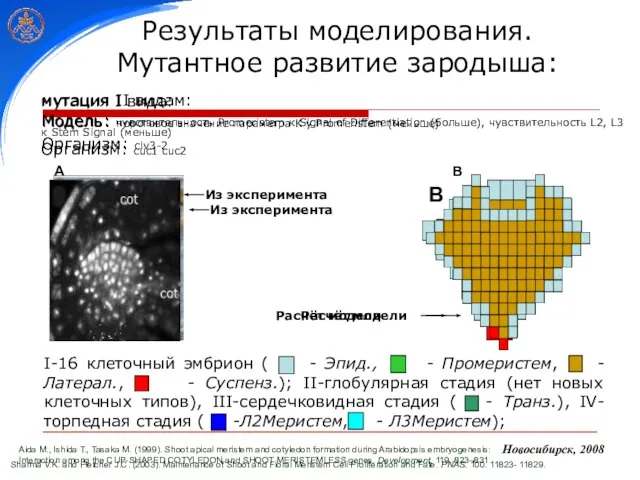
Слайд 27 Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and Floral
Sharma V.K. and Fletcher J.C. (2003). Maintenance of Shoot and Floral
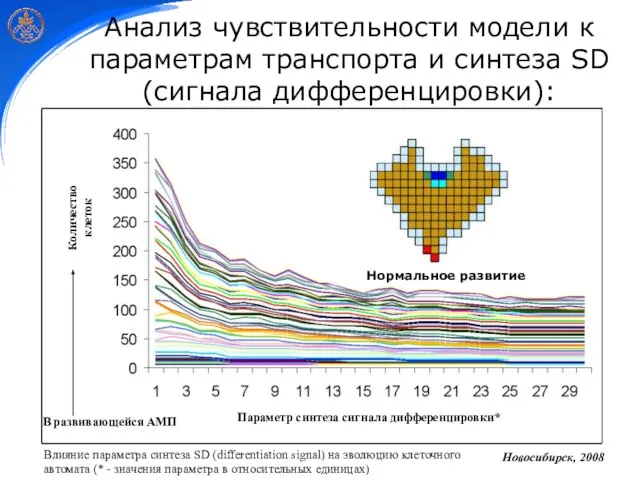
Слайд 28Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
В развивающейся
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
В развивающейся
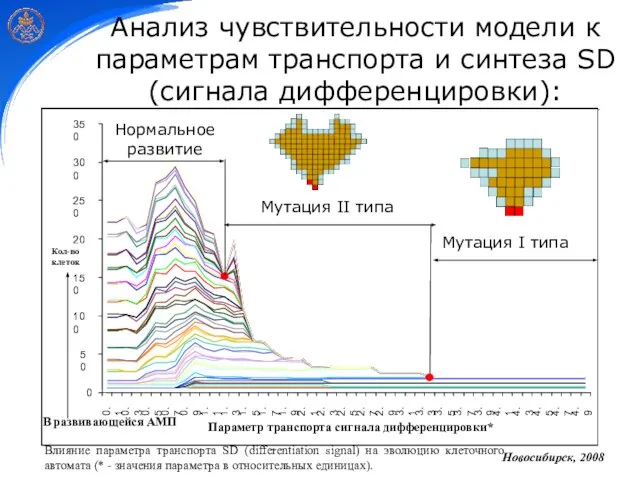
Слайд 29Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
Нормальное развитие
Мутация
Анализ чувствительности модели к параметрам транспорта и синтеза SD (сигнала дифференцировки):
Нормальное развитие
Мутация
Слайд 30Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Иммунная локализация белков PIN1 в
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Иммунная локализация белков PIN1 в
Слайд 31Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Полярная локализация белков PIN1 на
Распределение паттернов ауксина
в развивающейся апикальной меристеме побега:
Полярная локализация белков PIN1 на
Слайд 32Новосибирск, 2008
Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие этапы:
Новосибирск, 2008
Разработана конвейерная технология конструирования математических моделей генных сетей, включающая следующие этапы:

Слайд 33Новосибирск, 2008
Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма ауксина
Новосибирск, 2008
Автоматически сгенерированная база математических моделей элементарных подсистем генной сети метаболизма ауксина

Слайд 34Новосибирск, 2008
Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana. Для
Новосибирск, 2008
Разработана пространственно-распределённая математическая модель развития меристемы побега Arabidopsis thaliana. Для

 Правовая политика Республики Казахстан и ее направления
Правовая политика Республики Казахстан и ее направления Компания Medical Icons
Компания Medical Icons Общее понятие контроля
Общее понятие контроля Формы записи алгоритмов
Формы записи алгоритмов Посадка под школу биохакинга
Посадка под школу биохакинга Автоматизация звука Р в середине слова
Автоматизация звука Р в середине слова В библиотеке Мурманского государственного технического университета с 15.11.2011 по 15.11.2012 открыт доступ к Электронно-библиотечной
В библиотеке Мурманского государственного технического университета с 15.11.2011 по 15.11.2012 открыт доступ к Электронно-библиотечной  Стратегический план развития
Стратегический план развития Правописание окончаний имен существительных 1-ого и 3-го склонения в Р.п., Д.п.
Правописание окончаний имен существительных 1-ого и 3-го склонения в Р.п., Д.п. ПЛАН 2022 2023
ПЛАН 2022 2023 Александр Иванович Куприн 11 класс
Александр Иванович Куприн 11 класс otnositelnost_dvizheniya
otnositelnost_dvizheniya Татьянин день
Татьянин день Производная – это сложно, но прожить без нее не возможно?
Производная – это сложно, но прожить без нее не возможно? Давньогрецький театр та його особливості
Давньогрецький театр та його особливості Современные VPD технологии MSC.Software для университетов
Современные VPD технологии MSC.Software для университетов Excel
Excel Применение Клеточной Хемилюминесценции
Применение Клеточной Хемилюминесценции Economic Justice
Economic Justice Фотоальбом
Фотоальбом Упорство и талант
Упорство и талант Путешествие в Царство Плутона
Путешествие в Царство Плутона простое решение с большими возможностями!
простое решение с большими возможностями! Эскалация_14.10
Эскалация_14.10 Гроза и молния
Гроза и молния Влияние добычи и переработки полезных ископаемых на окружающую среду
Влияние добычи и переработки полезных ископаемых на окружающую среду Алгоритмы канального уровня
Алгоритмы канального уровня Козловская Диана. Грамоты
Козловская Диана. Грамоты