- Молекулярные машины

Содержание
- 2. Marcello Malpighi 10.03.1628 – 29.11.1694 “Nature, in order to carry out the marvelous operations in animals
- 3. Bruce Alberts “The entire cell can be viewed as a factory that contains an elaborate network
- 4. Как устроены сложные механизмы?
- 5. Биологические моторы
- 6. Молекулярные машины - основные функции: упаковщики, носильщики, сборщики, уничтожители
- 7. Мотор «Топливо» Продукт Механический
- 8. Примеры молекулярных моторов 1. Cytoskeletal motors 2. Nucleic acid-based motors Причины интереса к молекулярным моторам: 1.
- 9. Цитоскелетная транспортная система = Мотор + Путь + Топливо
- 10. Цитоскелетные моторы Носильщики Гребцы Animated cartoon: MCRI, U.K. Kinesin-1 Myosin-V Myosin-II Science, 27 June (2003)
- 11. Woehlke and Schliwa (2000) Цитоскелетные моторы http://www.proweb.org/kinesin/CrystalStruc/Dimer-down-rotaxis.jpg
- 12. Линейные биологические моторы Все линейные биологические моторы превращают энергию гидролиза АТФ в механическую работу Белки-моторы передвигаются
- 13. Кинезин Кинезины — суперсемейство моторных белков эукариотических клеток. Кинезины двигаются по микротрубочкам, используя энергию гидролиза АТФ.
- 14. Кинезин – линейный биологический мотор
- 15. Миозин Миозины — семейство белков, являющихся моторами цитоскелета системы микрофиламентов. Миозин — фибриллярный белок, один из
- 16. Взаимодействие миозина и актина – часть мотора мышц
- 17. Миозин Единичная головка миозина двигается с регулярностью шага 5,5 нм вдоль нити актина Молекула миозина генерирует
- 18. Динеин Динеины — группа моторных белков, способных перемещаться по поверхности микротрубочек цитоскелета, и транформирующих химическую энергию,
- 19. Молекулярные машины процессинга нуклеиновых кислот
- 20. Комплекс репликации ДНК Репликация ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы
- 21. Комплекс репликации ДНК
- 22. Синтез белка на рибосоме Синтез белка является основой жизнедеятельности клетки. Для осуществления этого процесса в клетках
- 23. Трансляция
- 24. Роторные молекулярные моторы – АТФ-синтаза В 60-70 годах XX века Пол Бойер (Paul Delos Boyer) -
- 25. Основные субъединицы (α3β3) компонента F1 соединены дополнительной боковой ножкой с неподвижным участком FO, что предотвращает их
- 26. Роторные молекулярные моторы – АТФ-синтаза
- 27. Роторные молекулярные моторы – жгутиковый мотор E. coli
- 28. Роторные молекулярные моторы – жгутики Жгутик — поверхностная структура, присутствующая у многих прокариотических и эукариотических клеток
- 31. Роторные молекулярные моторы – жгутиковый мотор E. coli
- 32. В E. coli жгутиковый мотор может вращаться в обоих направлениях. Жгутиковые моторы могут развивать скорость 300
- 33. Перспективы На основе принципов работы биологических молекулярных моторов, с использованием достижений нанотехнологии создаются миниатюрные механические наномоторы
- 35. Скачать презентацию
Слайд 2Marcello Malpighi
10.03.1628 – 29.11.1694
“Nature, in order to carry out the marvelous operations
Marcello Malpighi
10.03.1628 – 29.11.1694
“Nature, in order to carry out the marvelous operations

Слайд 3Bruce Alberts
“The entire cell can be viewed as a factory that
Bruce Alberts
“The entire cell can be viewed as a factory that

Слайд 4Как устроены сложные механизмы?
Как устроены сложные механизмы?

Слайд 5Биологические моторы
Биологические моторы

Слайд 6Молекулярные машины - основные функции:
упаковщики,
носильщики,
сборщики,
уничтожители
Молекулярные машины - основные функции:
упаковщики,
носильщики,
сборщики,
уничтожители

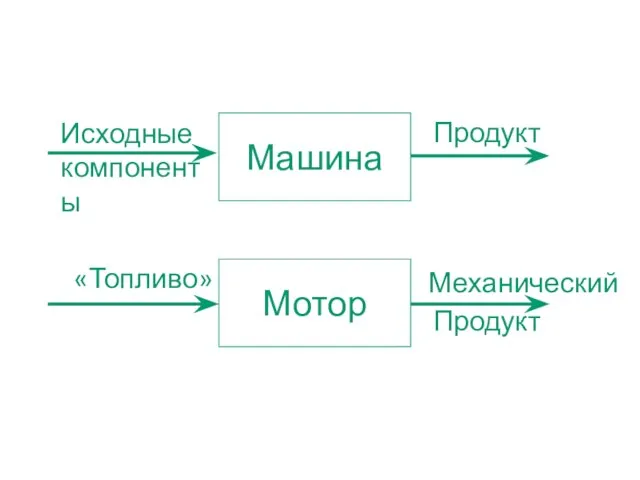
Слайд 7Мотор
«Топливо»
Продукт
Механический
Мотор
«Топливо»
Продукт
Механический
Слайд 8Примеры молекулярных моторов
1. Cytoskeletal motors
2. Nucleic acid-based motors
Причины интереса к молекулярным
Примеры молекулярных моторов
1. Cytoskeletal motors
2. Nucleic acid-based motors
Причины интереса к молекулярным

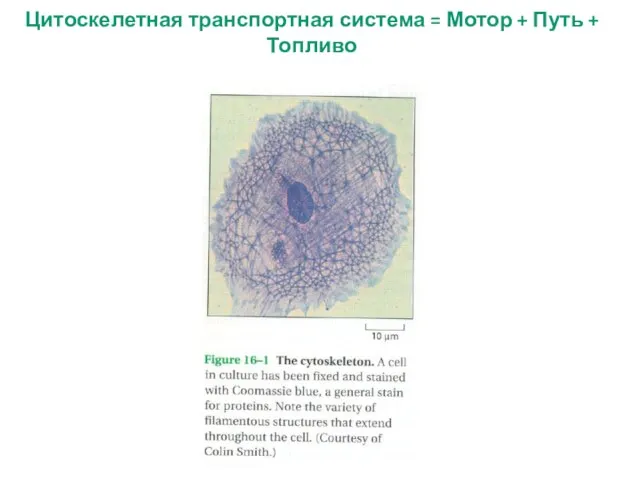
Слайд 9Цитоскелетная транспортная система = Мотор + Путь + Топливо
Цитоскелетная транспортная система = Мотор + Путь + Топливо
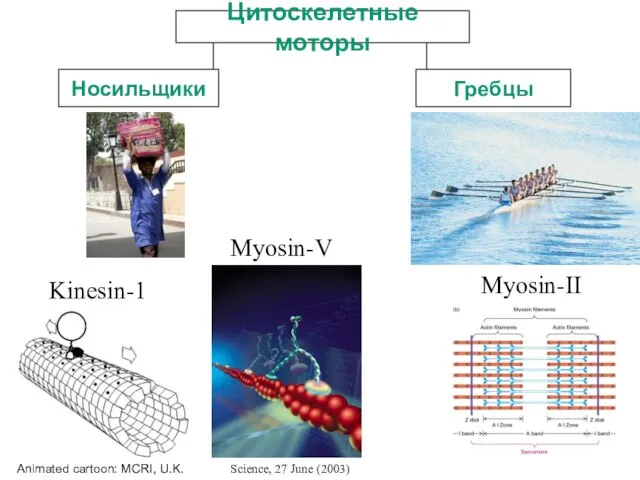
Слайд 10Цитоскелетные моторы
Носильщики
Гребцы
Animated cartoon: MCRI, U.K.
Kinesin-1
Myosin-V
Myosin-II
Science, 27 June (2003)
Цитоскелетные моторы
Носильщики
Гребцы
Animated cartoon: MCRI, U.K.
Kinesin-1
Myosin-V
Myosin-II
Science, 27 June (2003)
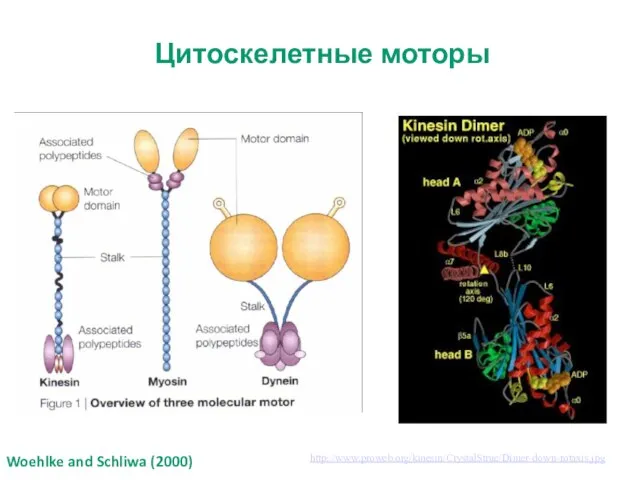
Слайд 11Woehlke and Schliwa (2000)
Цитоскелетные моторы
http://www.proweb.org/kinesin/CrystalStruc/Dimer-down-rotaxis.jpg
Woehlke and Schliwa (2000)
Цитоскелетные моторы
http://www.proweb.org/kinesin/CrystalStruc/Dimer-down-rotaxis.jpg
Слайд 12Линейные биологические моторы
Все линейные биологические моторы превращают энергию гидролиза АТФ в механическую
Линейные биологические моторы
Все линейные биологические моторы превращают энергию гидролиза АТФ в механическую

Слайд 13Кинезин
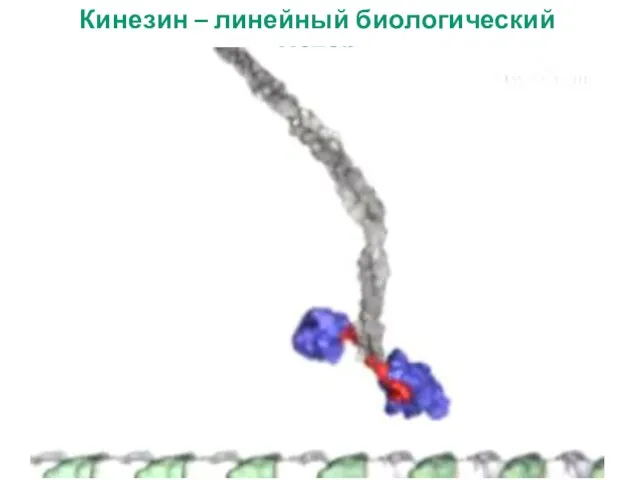
Кинезины — суперсемейство моторных белков эукариотических клеток. Кинезины двигаются по микротрубочкам, используя
Кинезин
Кинезины — суперсемейство моторных белков эукариотических клеток. Кинезины двигаются по микротрубочкам, используя

Слайд 14Кинезин – линейный биологический мотор
Кинезин – линейный биологический мотор
Слайд 15Миозин
Миозины — семейство белков, являющихся
моторами цитоскелета системы микрофиламентов. Миозин — фибриллярный белок,
Миозин
Миозины — семейство белков, являющихся
моторами цитоскелета системы микрофиламентов. Миозин — фибриллярный белок,

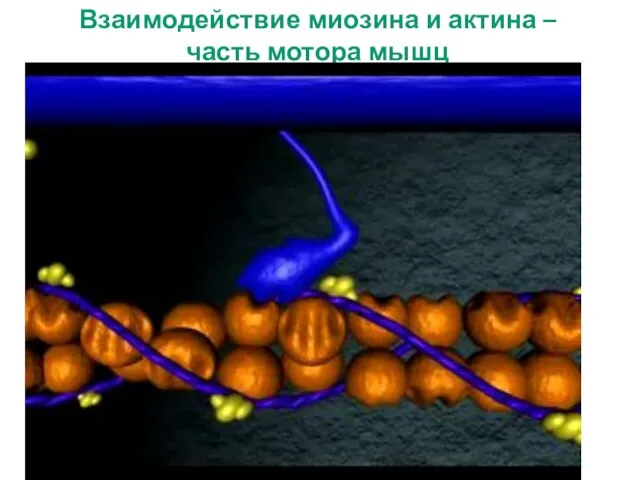
Слайд 16Взаимодействие миозина и актина – часть мотора мышц
Взаимодействие миозина и актина – часть мотора мышц
Слайд 17Миозин
Единичная головка миозина двигается с регулярностью шага 5,5 нм вдоль нити актина
Молекула
Миозин
Единичная головка миозина двигается с регулярностью шага 5,5 нм вдоль нити актина
Молекула

Слайд 18Динеин
Динеины — группа моторных белков, способных перемещаться по поверхности микротрубочек цитоскелета, и
Динеин
Динеины — группа моторных белков, способных перемещаться по поверхности микротрубочек цитоскелета, и

Слайд 19Молекулярные машины процессинга нуклеиновых кислот
Молекулярные машины процессинга нуклеиновых кислот

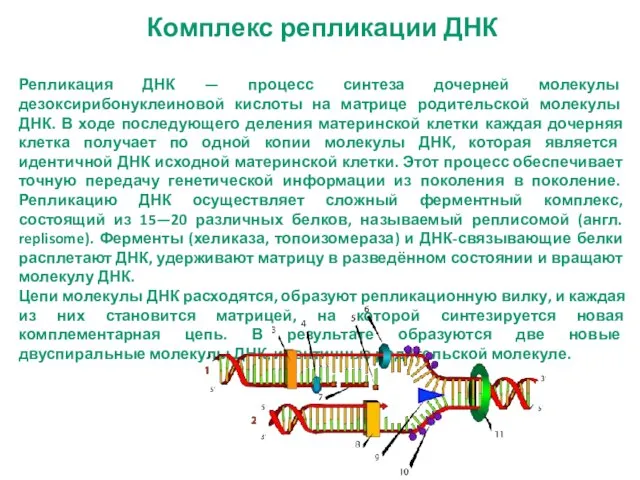
Слайд 20Комплекс репликации ДНК
Репликация ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на
Комплекс репликации ДНК
Репликация ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на
Слайд 21Комплекс репликации ДНК
Комплекс репликации ДНК

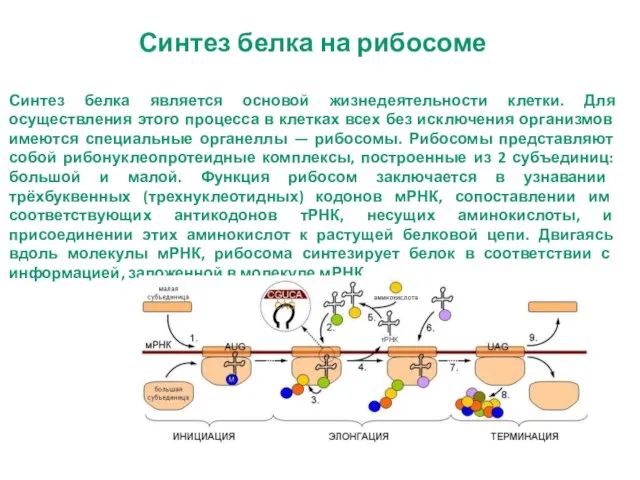
Слайд 22Синтез белка на рибосоме
Синтез белка является основой жизнедеятельности клетки. Для осуществления этого
Синтез белка на рибосоме
Синтез белка является основой жизнедеятельности клетки. Для осуществления этого
Слайд 23Трансляция
Трансляция

Слайд 24Роторные молекулярные моторы – АТФ-синтаза
В 60-70 годах XX века Пол Бойер (Paul
Роторные молекулярные моторы – АТФ-синтаза
В 60-70 годах XX века Пол Бойер (Paul

Слайд 25Основные субъединицы (α3β3) компонента F1 соединены дополнительной боковой ножкой с неподвижным участком
Основные субъединицы (α3β3) компонента F1 соединены дополнительной боковой ножкой с неподвижным участком
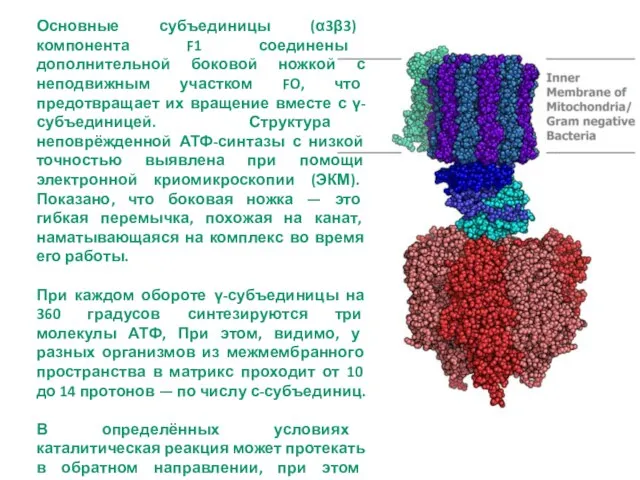
Слайд 26Роторные молекулярные моторы – АТФ-синтаза
Роторные молекулярные моторы – АТФ-синтаза

Слайд 27Роторные молекулярные моторы – жгутиковый мотор E. coli
Роторные молекулярные моторы – жгутиковый мотор E. coli
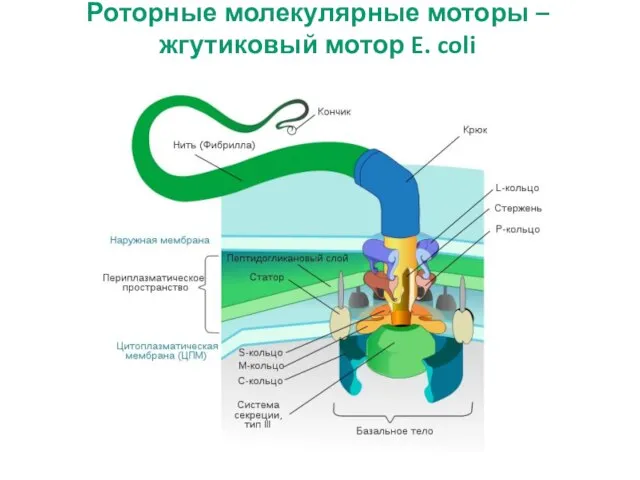
Слайд 28Роторные молекулярные моторы – жгутики
Жгутик — поверхностная структура, присутствующая у многих прокариотических
Роторные молекулярные моторы – жгутики
Жгутик — поверхностная структура, присутствующая у многих прокариотических


Слайд 31Роторные молекулярные моторы – жгутиковый мотор E. coli
Роторные молекулярные моторы – жгутиковый мотор E. coli

Слайд 32В E. coli жгутиковый мотор может вращаться в обоих направлениях. Жгутиковые моторы
В E. coli жгутиковый мотор может вращаться в обоих направлениях. Жгутиковые моторы

Слайд 33Перспективы
На основе принципов работы биологических молекулярных моторов, с использованием достижений нанотехнологии создаются
Перспективы
На основе принципов работы биологических молекулярных моторов, с использованием достижений нанотехнологии создаются

 Презентация по литературному чтению на тему _Биография И.С. Никитина_
Презентация по литературному чтению на тему _Биография И.С. Никитина_ Бренд The Macallan
Бренд The Macallan body language
body language  = 600 : 72
= 600 : 72 Государственные награды за воинские отличия в России
Государственные награды за воинские отличия в России NON ANCORA RITOCCATA
NON ANCORA RITOCCATA МОДЕЛИ ЗАРАБОТКА В ИНТЕРНЕТЕ
МОДЕЛИ ЗАРАБОТКА В ИНТЕРНЕТЕ Презентация на тему Духовный мир древних обществ
Презентация на тему Духовный мир древних обществ ПРОФИЛАКТИКА ТУБЕРКУЛЕЗА
ПРОФИЛАКТИКА ТУБЕРКУЛЕЗА Инклюзивное дошкольное образование
Инклюзивное дошкольное образование 24 мая-день славянской письменности
24 мая-день славянской письменности Справочные сведения о МКОУСОШ №3
Справочные сведения о МКОУСОШ №3 История школьной оценки
История школьной оценки ЭМОЦИОНАЛЬНАЯ ЗАВИСИМОСТЬ
ЭМОЦИОНАЛЬНАЯ ЗАВИСИМОСТЬ Презентация на тему Песни и стихи о Великой Отечественной Войне
Презентация на тему Песни и стихи о Великой Отечественной Войне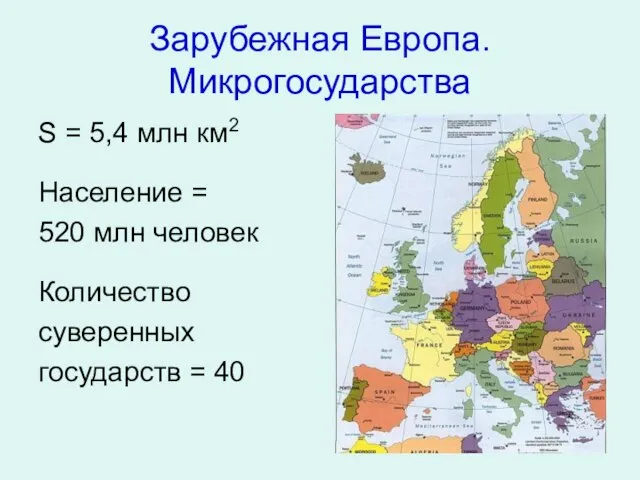 Зарубежная Европа. Микрогосударства
Зарубежная Европа. Микрогосударства 7.6.
7.6. ДОКЛАД ОБ ЭФФЕКТИВНОСТИ ДЕЯТЕЛЬНОСТИ ОРГАНОВ ИСПОЛНИТЕЛЬНОЙ ВЛАСТИ СУБЪЕКТОВ РОССИЙСКОЙ ФЕДЕРАЦИИ по итогам 2010 года
ДОКЛАД ОБ ЭФФЕКТИВНОСТИ ДЕЯТЕЛЬНОСТИ ОРГАНОВ ИСПОЛНИТЕЛЬНОЙ ВЛАСТИ СУБЪЕКТОВ РОССИЙСКОЙ ФЕДЕРАЦИИ по итогам 2010 года Ф.М. Достоевский "Преступление и наказание"
Ф.М. Достоевский "Преступление и наказание" Фризский язык
Фризский язык Качественное образование и успешная профессиональная карьера
Качественное образование и успешная профессиональная карьера Красота простоты
Красота простоты ЖУМАГАЛИЕВА ПРЕЗЕНТАЦИЯ
ЖУМАГАЛИЕВА ПРЕЗЕНТАЦИЯ Презентация на тему Засоби I допомоги при захворюваннях серцево-судинної системи
Презентация на тему Засоби I допомоги при захворюваннях серцево-судинної системи  Об участии в государственной итоговой аттестации выпускников 9-го класса в 2022 году
Об участии в государственной итоговой аттестации выпускников 9-го класса в 2022 году Туристско-рекреационный паспорт территории
Туристско-рекреационный паспорт территории Музеи Прибайкалья
Музеи Прибайкалья Музеи мира
Музеи мира