- Репарация генетических повреждений

Содержание
- 2. Часть 1 Мутации и мутагены
- 3. Основные термины Мутации – это явления скачкообразного, прерывистого изменения наследственного признака. (определение Г. Де Фриза) Мутации
- 4. Мутационная теория Основные положения мутационной теории Г. Де Фриза сводятся к следующему: Мутации возникают внезапно, как
- 5. Классификация мутаций В зависимости от факторов, вызывающих мутации, их разделяют на спонтанные и индуцированные. Спонтанные мутации
- 6. Классификация мутаций В зависимости от размеров сегментов генома, подвергающихся преобразованиям, мутации разделяют на геномные, хромосомные и
- 7. Классификация мутаций При хромосомных мутациях происходят как изменение числа отдельных хромосом в геноме (анеуплоидия), так и
- 8. Классификация мутаций Генные мутации - изменения первичной структуры ДНК генов под действием мутагенных факторов (встречаются более
- 9. Классификация мутаций Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций: Синонимическая замена нуклеотида
- 10. Классификация мутаций По влиянию на экспрессию генов мутации разделяют на две категории: Мутации замен пар оснований
- 11. Классификация мутаций Первичную мутацию называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена – обратной мутацией
- 12. Основные источники мутаций В основе мутаций на молекулярном уровне лежат две основные причины: Ошибки репликации Мутагенные
- 13. Ошибки репликации Точность процесса репликации определяется: Различиями в свободной энергии у канонических или ошибочных пар азотистых
- 14. Мутагенные воздействия Усилий систем репликации становится недостаточно в стрессовых ситуациях, когда организм подвергается массированному мутагенному воздействию.
- 15. Ионизирующее излучение Ярко выраженным мутагенным действием обладают: Коротковолновое электромагнитное излучение (УФ-свет, рентгеновские лучи) Элементарные частицы, образующиеся
- 16. Химические мутагены Многие химические соединения обладают способностью взаимодействовать с ДНК или с ее низкомолекулярными предшественниками и
- 17. Алкилирующие агенты Наиболее обширным классом химических мутагенов экзогенного происхождения являются алкилирующие агенты. Механизм повреждающего воздействия заключается
- 18. Алкилирующие агенты Главным источником мутаций, возникающих под действием алкилирующих агентов, является алкилирование O-6 в гуанине и
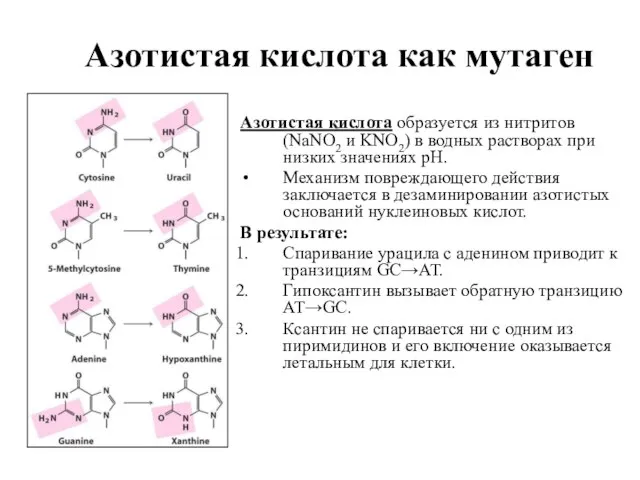
- 20. Азотистая кислота как мутаген Азотистая кислота образуется из нитритов (NaNO2 и KNO2) в водных растворах при
- 21. Дезаминирование основание мутаген (азотистая кислота) модиф. основание партнер по спариванию мутагенный эффект
- 22. Органические перекиси как мутагены Мутагенным действием обладают различные органические перекиси. Азид натрия – мощный ингибитор дыхания,
- 23. Активные формы кислорода В клетках активные формы кислорода (АФК) возникают в реакциях восстановления, в результате которых
- 24. Активные формы кислорода
- 25. Аналоги нуклеозидов и оснований Аналоги нуклеозидов и оснований: 5-бромдезоксиуридин и 2-аминопурин, так же являются сильными мутагенами.
- 26. Интеркалирующие соединения Красители нуклеиновых кислот, обладающие способностью интеркалировать между основаниями ДНК, вызывают мутации со сдвигом рамки
- 28. Метаболическая активация проканцерогенов Для защиты от накопления экзогенных чужеродных химических соединений (ксенобиотиков) у каждого живого организма
- 29. Цитохромы Р-450 В первой фазе метаболизма ксенобиотиков принимают участие цитохромы группы Р-450. Эти ферменты в организме
- 31. Эндогенные мутагены Молекулы ДНК часто претерпевают in vivo тепловую депуринизацию, которая может быть спонтанным внутренним источником
- 32. Эндогенные мутагены Источником эндогенных мутаций служит самопроизвольное дезаминирование цитозина в составе ДНК с образованием урацила. 5-Метилцитозин
- 33. Эндогенные мутагены Источником эндогенных мутагенов в организме является метаболизм нормальной микрофлоры. Некоторые промежуточные соединения, возникающие при
- 34. Часть 2 Механизмы репарации
- 35. Репарация генетических повреждений – это свойство живых организмов восстанавливать повреждения, возникшие в ДНК в результате ошибок
- 36. Общие сведения В настоящее время описано множество различных систем репарации, часто принципиально отличных друг от друга.
- 37. Основной принцип репарации Основан на двуспиральном строении ДНК. В большинстве случаев поврежденной оказывается только одна цепь
- 38. Повреждения ДНК Появление различно модифицированных оснований: Пиримидиновые димеры. Алкилированые производные. Дезаминированые основания. Различные таутомерные формы. Появление
- 39. Основные повреждающие факторы Ионизирующие агенты: Ультрафиолетовый свет. Радиоактивные вещества. Активные формы кислорода. Химические мутагены (например алкилирующие
- 40. Пиримидиновые димеры Расстояние между параллельными плоскостями оснований в В-форме оказывается как раз таким, чтобы освободившиеся при
- 41. Таутомерные переходы Таутомерия (от греч. tautós — тот же самый и méros — доля, часть), быстрая
- 45. Разнообразие систем репарации Существует огромное количество самых различных систем репарации. Все эти системы появлялись в эволюции
- 46. Разнообразие систем репарации Прямая репарация: Фотореактивация. Дезалкилирование модифицированных нуклеотидов. Сшивание однонитевых разрывов. Прямая вставка оснований в
- 47. Фотореактивация В фотолиазе есть участок, служащий светочувствительным центром, который способен адсорбировать фотоны. Метенилтетрагидрофолатное производное выполняет роль
- 48. Фотореактивация Система ферментативной фотореактивации ДНК (photoreactivation – PHR), основным компонентом которой является ДНК-фотолиаза, разделяет пиримидиновые димеры,
- 49. Структура фотолиазы Фотолиаза содержит два кофактора: светоулавливающий метенилтетрагидрофолат (HDF) и генерирующий свободные радикалы FADH–. Оба они
- 50. Репарация алкилированных оснований В клетках синтезируются белки метилтрансферазы, которые могут захватывать метильные группы от модифицированного основания
- 51. Сшивание однонитевых разрывов: Этот тип реакций прямой репарации был обнаружен для однонитевых разрывов ДНК, индуцируемых ионизирующим
- 52. Вставка оснований в АП-сайт Ковалентная связь между основанием и сахаром (β-гликозид-ная связь) может рваться. Тогда в
- 53. Эксцизионная репарация ДНК путем удаления поврежденных азотистых оснований (BER)
- 54. Base excision repair – BER Система BER обеспечивает защиту геномной ДНК от повреждений, вызываемых главным образом
- 55. Разнообразие гликозилаз
- 56. Существует много ДНК-гликозилаз, которые адресно узнают различные повреждения
- 57. Структура гликозилаз
- 58. Механизм работы гликозилаз Механизм связывания поврежден-ного основания гликозилазой имеет много сходных моментов с меха-низмом захвата метилазами
- 59. ДНК гликозилазы «выворачивают» модифицированное основание наружу и отщепляют его от сахаро-фосфатного остова
- 60. Base excision repair – BER Гликозилазы присоединяются модифицированным основаниям и гидролизуют β-N- гликозидные связи между основанием
- 61. Base excision repair – BER Появившаяся брешь в одной цепи ДНК размером в один нуклеотид застраивается
- 62. Эксцизионная репарация ДНК путем удаления нуклеотидов (NER)
- 63. Nucleotide excision repair – NER Процесс NER условно можно разделить на четыре этапа: Распознавание поврежденного участка
- 64. Nucleotide excision repair – NER В отличии от BER, субстратами системы NER являются не только поврежденные
- 65. Механизм работы Белковые ножницы, содержащие две копии белка UvrA и одну копию UvrB, узнают поврежденный участок
- 66. Механизм работы Белок UvrC присоединяется к комплексу UvrВ – ДНК; белок UvrВ делает 3’-надрез, а UvrC
- 67. У эукариот механизм эксцизии нуклеотидов в общих чертах схож с прокариотическим, но существенно отличается в деталях
- 68. Повреждения ДНК могут вызвать остановку элонгирующей РНК-полимеразы Ферменты NER узнают такой задержанный комплекс и процессируют его
- 69. XPB & XPD – TFII H DNA helicase XPC - damage recognition XPA stabilisation of SS
- 70. Различия NER у про- и эукариот Гены NER у E. coli uvrA, uvrB и uvrC не
- 71. Репарация ошибочно спаренных нуклеотидов (MMR)
- 72. Mismatch repair - MMR В отличие от NER, так же удаляющей неправильно спаренные основания, MMR может
- 73. Метилирование матричных цепей Обычно у E. coli ДНК метилирована Dam-метилазой по сайтам GATC. После завершения репликации
- 74. Механизм работы На начальных этапах система MMR задействует белковые продукты четырех генов: mutH, mutL, mutS и
- 75. Механизм работы После полной сборки комплекса MutHLS, активации эндонуклеазной активности MutH и внесения разрывов в GATC-сайты
- 76. Другие системы У E. coli существуют два других специфических пути репарации ошибочно спаренных нуклеотидов: Система VSP
- 77. гомологи MutS (MSH — MutS homolog) образуют два гетеродимерных комплекса -- MSH2-MSH6 (MutSα) узнает неспаренные нуклеотиды
- 78. Non-Homologous End Joining (Double Strand Breaks)
- 80. Пострепликативная (рекомбинационная) репарация
- 81. Рекомбинационная репарация Быстро делящиеся бактериальные клетки, содержащие несколько репликонов, образованных недореплицированными хромосомами, более устойчивы к действию
- 82. Рекомбинационная репарация Рекомбинационная репарация необходима в случае, когда повреждены обе цепи ДНК, что может быть результатом:
- 83. Рекомбинационная репарация Второй дуплекс ДНК, полученный в результате репликации, не содержит в себе разрывов или модифицированных
- 84. Механизм процесса Главный этап рекомбинационной репарации заключается в активации генов rec, участвующих так же и в
- 85. Механизм процесса В синаптической фазе наблюдается синапсис гомологичных участков двух молекул ДНК с вхождением комплементарного одноцепочечного
- 86. Рекомбинационная репарация
- 87. Часть 3 SOS-репарация и SOS-мутагенез
- 88. SOS-репарация В клетках организма, подвергнутого сильному мутагенному воздействию, образование мутаций происходит в основном по механизму SOS-мутагенеза.
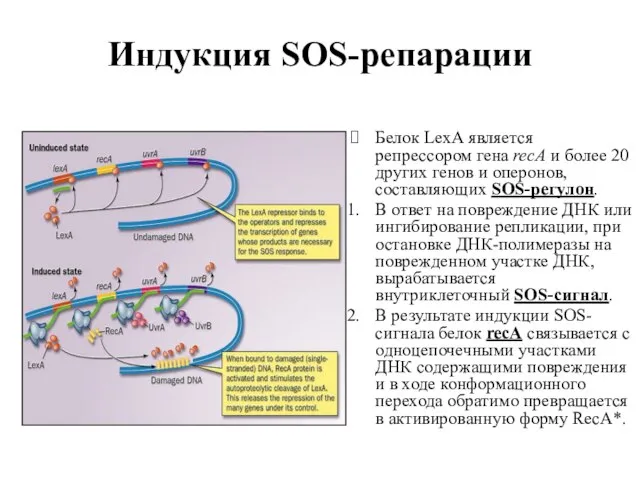
- 89. Индукция SOS-репарации Белок LexA является репрессором гена recA и более 20 других генов и оперонов, составляющих
- 90. Активация белка RecA Точно еще не выяснено как индукции SOS-сигнала, как и его природа, влияет на
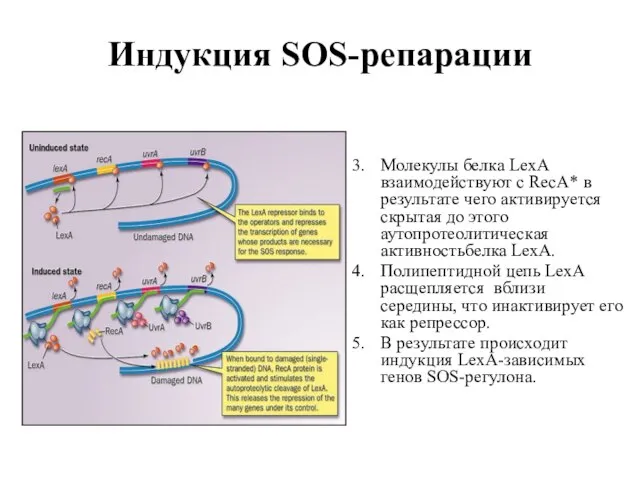
- 91. Индукция SOS-репарации Молекулы белка LexA взаимодействуют с RecA* в результате чего активируется скрытая до этого аутопротеолитическая
- 92. Основные термины SOS-репарация (мутагенез) – это согласованная индукция множества ферментов, включая ферменты репарации, в ответ на
- 93. Индукция SOS-репарации Ключевыми белками, задействоваными в процессе введения ошибок ДНК-полимеразой, являются продукты двух генов – umuD
- 94. ДНК-полимераза 5 Настоящая роль белкового комплекса (UmuD’)2–UmuC в процессе SOS-репарации неизвестна. Существует две основных концепции: Комплекс
- 95. Индукция SOS-репарации Когда репликаза (ДНК-полимераза 3) доходит до некоторого повреждения, например тиминового димера, она останавливается. Белок
- 96. Таким образом белок RecA играет ключевую роль в индукции SOS-репарации, выполняя следующие важные функции: Взаимодействует с
- 97. ДНК-полимераза 4 ДНК-полимераза IV E. coli, кодируемая геном dinB, также участвует в SOS-ответе бактерий: Она не
- 98. После того, как весь геном был успешно реплицирован, путем введения мутации в последовательности, которые блокировали продвижение
- 99. Вырезание профагов Активация RecA так же может являться причиной расщепления и других репрессорных белков, например репрессоров
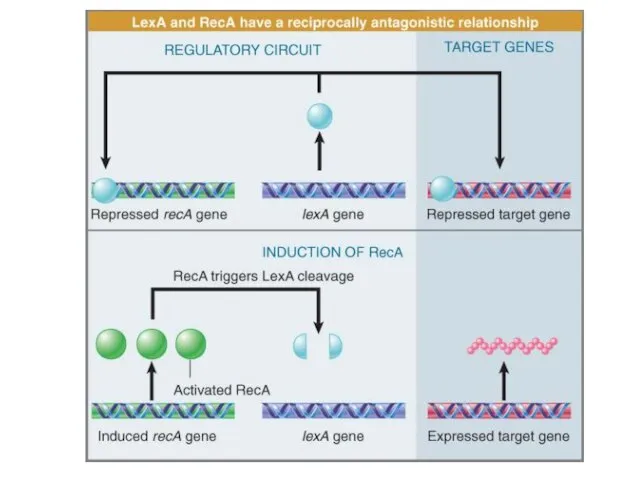
- 100. Прежде всего об остановке репликации надо сообщить Rec A Cвязывается с однонитевой ДНК и образует ДНК-белковые
- 101. lexA, recA, uvrA, uvrB, and uvrD Филаменты Rec A стимулируют аутопротеолиз Lex A При снижении концентрации
- 103. UmuD повергается автопротеолитическому расщеплению с образованием активного фрагмента UmuD’ UmuD’ активирует черезблоковую полимеразу UmuC комплекс (UmuD’)2-UmuC
- 106. Скачать презентацию

Слайд 3Основные термины
Мутации – это явления скачкообразного, прерывистого изменения наследственного признака. (определение Г.
Основные термины
Мутации – это явления скачкообразного, прерывистого изменения наследственного признака. (определение Г.

Слайд 4Мутационная теория
Основные положения мутационной теории Г. Де Фриза сводятся к следующему:
Мутации возникают
Мутационная теория
Основные положения мутационной теории Г. Де Фриза сводятся к следующему:
Мутации возникают

Слайд 5Классификация мутаций
В зависимости от факторов, вызывающих мутации, их разделяют на спонтанные и
Классификация мутаций
В зависимости от факторов, вызывающих мутации, их разделяют на спонтанные и

Слайд 6Классификация мутаций
В зависимости от размеров сегментов генома, подвергающихся преобразованиям, мутации разделяют на
Классификация мутаций
В зависимости от размеров сегментов генома, подвергающихся преобразованиям, мутации разделяют на

Слайд 7Классификация мутаций
При хромосомных мутациях происходят как изменение числа отдельных хромосом в геноме
Классификация мутаций
При хромосомных мутациях происходят как изменение числа отдельных хромосом в геноме

Слайд 8Классификация мутаций
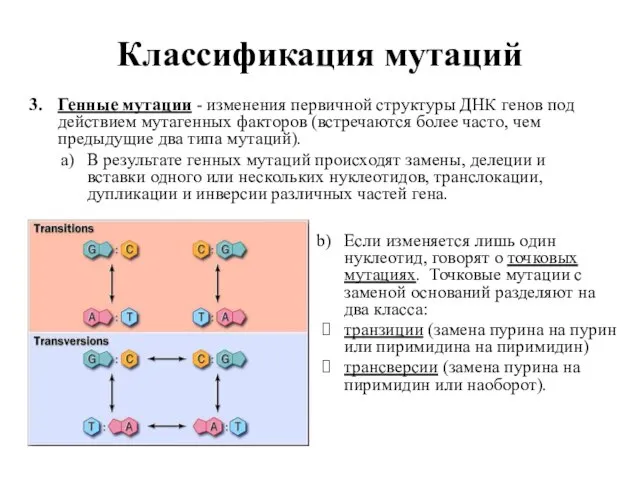
Генные мутации - изменения первичной структуры ДНК генов под действием мутагенных
Классификация мутаций
Генные мутации - изменения первичной структуры ДНК генов под действием мутагенных
Слайд 9Классификация мутаций
Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций:
Классификация мутаций
Из-за вырожденности генетического кода могут быть три генетических последствия точковых мутаций:

Слайд 10Классификация мутаций
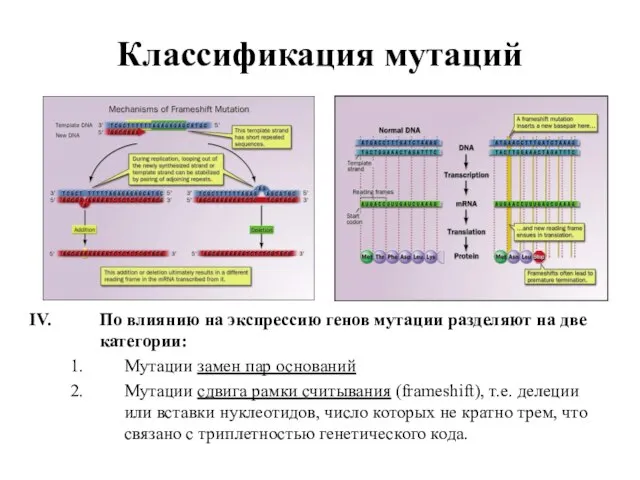
По влиянию на экспрессию генов мутации разделяют на две категории:
Мутации замен
Классификация мутаций
По влиянию на экспрессию генов мутации разделяют на две категории:
Мутации замен
Слайд 11Классификация мутаций
Первичную мутацию называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена
Классификация мутаций
Первичную мутацию называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена

Слайд 12Основные источники мутаций
В основе мутаций на молекулярном уровне лежат две основные причины:
Основные источники мутаций
В основе мутаций на молекулярном уровне лежат две основные причины:

Слайд 13Ошибки репликации
Точность процесса репликации определяется:
Различиями в свободной энергии у канонических или ошибочных
Ошибки репликации
Точность процесса репликации определяется:
Различиями в свободной энергии у канонических или ошибочных

Слайд 14Мутагенные воздействия
Усилий систем репликации становится недостаточно в стрессовых ситуациях, когда организм подвергается
Мутагенные воздействия
Усилий систем репликации становится недостаточно в стрессовых ситуациях, когда организм подвергается

Слайд 15Ионизирующее излучение
Ярко выраженным мутагенным действием обладают:
Коротковолновое электромагнитное излучение (УФ-свет, рентгеновские лучи)
Элементарные частицы,
Ионизирующее излучение
Ярко выраженным мутагенным действием обладают:
Коротковолновое электромагнитное излучение (УФ-свет, рентгеновские лучи)
Элементарные частицы,

Слайд 16Химические мутагены
Многие химические соединения обладают способностью взаимодействовать с ДНК или с ее
Химические мутагены
Многие химические соединения обладают способностью взаимодействовать с ДНК или с ее

Слайд 17Алкилирующие агенты
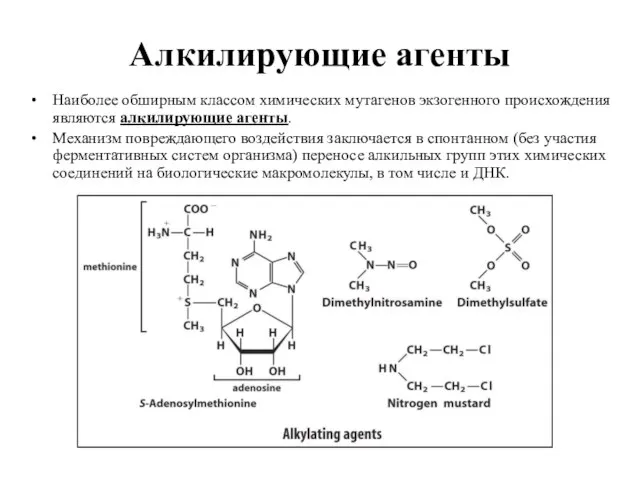
Наиболее обширным классом химических мутагенов экзогенного происхождения являются алкилирующие агенты.
Механизм повреждающего
Алкилирующие агенты
Наиболее обширным классом химических мутагенов экзогенного происхождения являются алкилирующие агенты.
Механизм повреждающего
Слайд 18Алкилирующие агенты
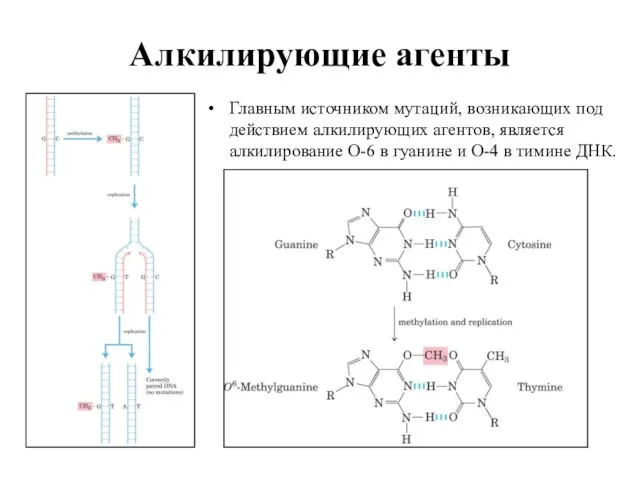
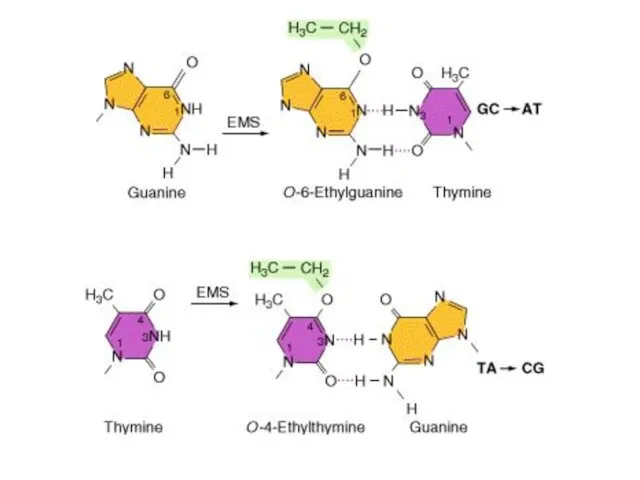
Главным источником мутаций, возникающих под действием алкилирующих агентов, является алкилирование O-6
Алкилирующие агенты
Главным источником мутаций, возникающих под действием алкилирующих агентов, является алкилирование O-6
Слайд 20Азотистая кислота как мутаген
Азотистая кислота образуется из нитритов (NaNO2 и KNO2) в
Азотистая кислота как мутаген
Азотистая кислота образуется из нитритов (NaNO2 и KNO2) в
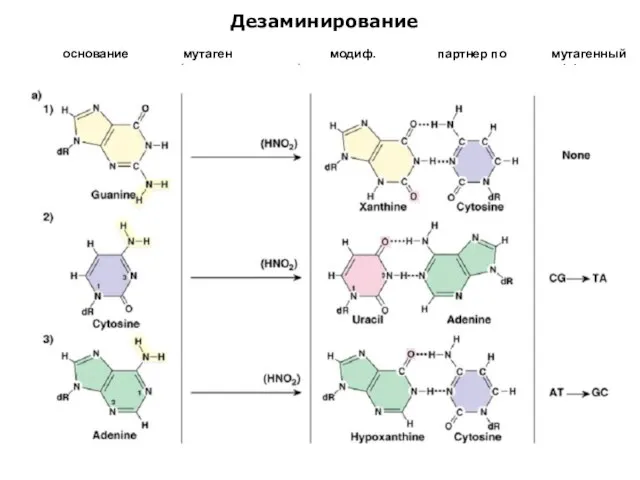
Слайд 21Дезаминирование
основание
мутаген
(азотистая кислота)
модиф.
основание
партнер по
спариванию
мутагенный
эффект
Дезаминирование
основание
мутаген
(азотистая кислота)
модиф.
основание
партнер по
спариванию
мутагенный
эффект
Слайд 22Органические перекиси как мутагены
Мутагенным действием обладают различные органические перекиси.
Азид натрия –
Органические перекиси как мутагены
Мутагенным действием обладают различные органические перекиси.
Азид натрия –

Слайд 23Активные формы кислорода
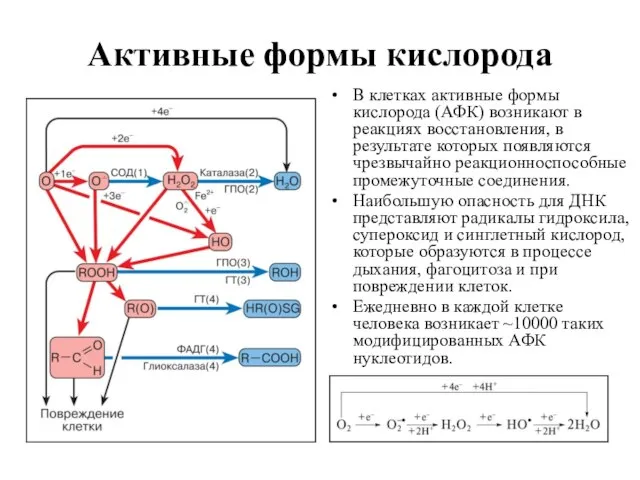
В клетках активные формы кислорода (АФК) возникают в реакциях восстановления,
Активные формы кислорода
В клетках активные формы кислорода (АФК) возникают в реакциях восстановления,
Слайд 24Активные формы кислорода
Активные формы кислорода
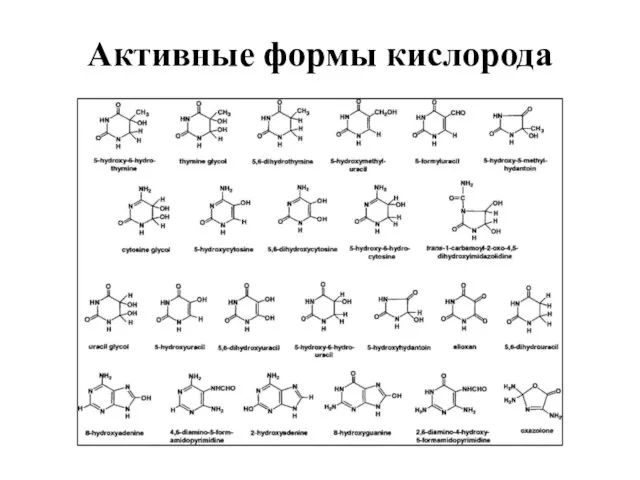
Слайд 25Аналоги нуклеозидов и оснований
Аналоги нуклеозидов и оснований: 5-бромдезоксиуридин и 2-аминопурин, так же
Аналоги нуклеозидов и оснований
Аналоги нуклеозидов и оснований: 5-бромдезоксиуридин и 2-аминопурин, так же
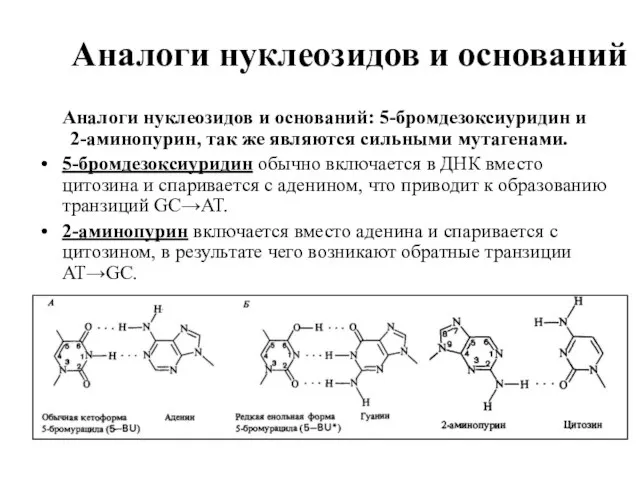
Слайд 26Интеркалирующие соединения
Красители нуклеиновых кислот, обладающие способностью интеркалировать между основаниями ДНК, вызывают мутации
Интеркалирующие соединения
Красители нуклеиновых кислот, обладающие способностью интеркалировать между основаниями ДНК, вызывают мутации
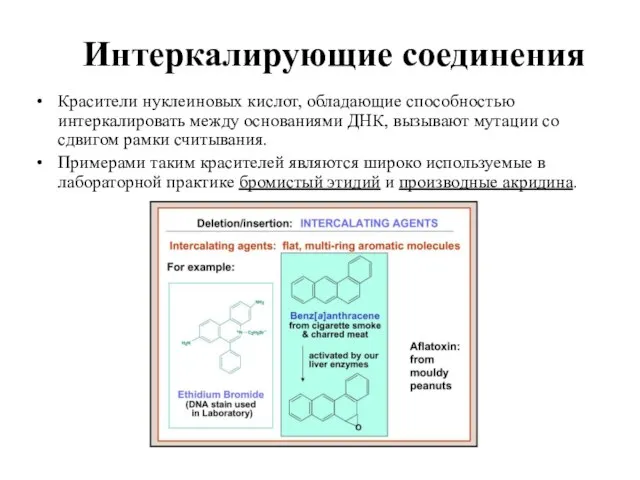
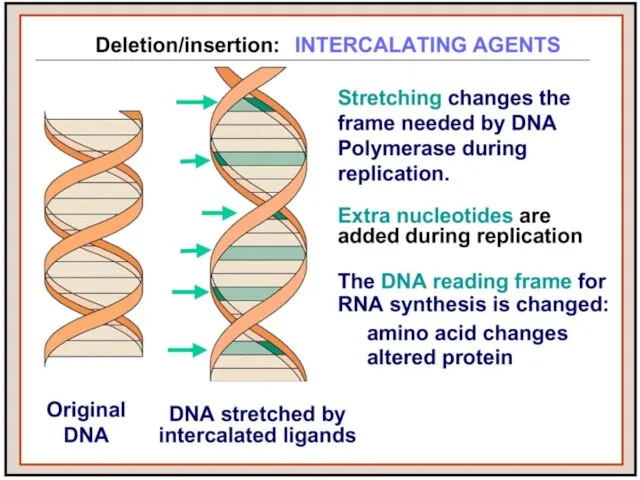
Слайд 28Метаболическая активация проканцерогенов
Для защиты от накопления экзогенных чужеродных химических соединений (ксенобиотиков) у
Метаболическая активация проканцерогенов
Для защиты от накопления экзогенных чужеродных химических соединений (ксенобиотиков) у

Слайд 29Цитохромы Р-450
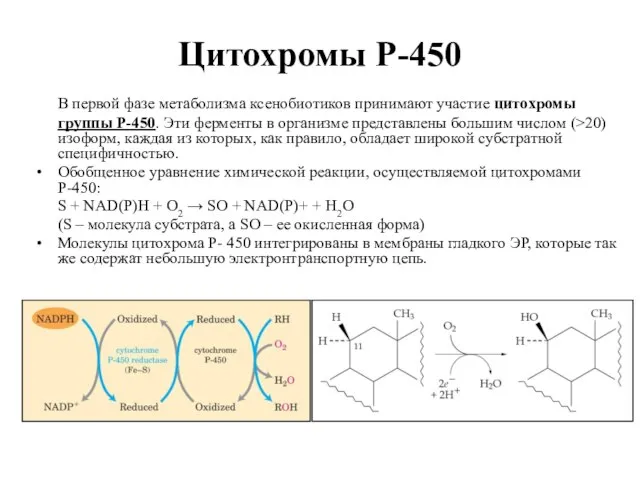
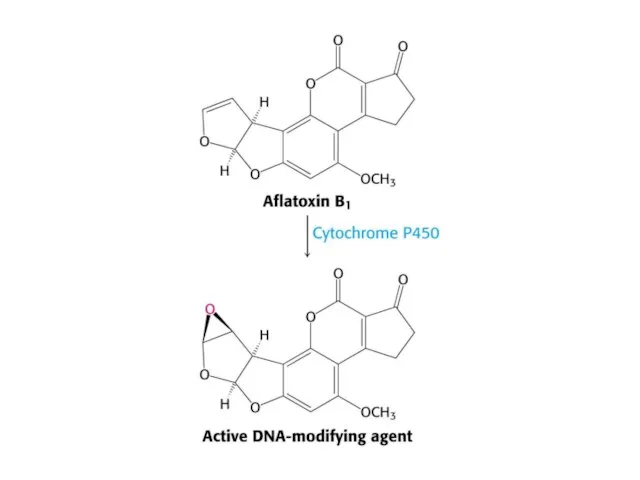
В первой фазе метаболизма ксенобиотиков принимают участие цитохромы группы Р-450. Эти
Цитохромы Р-450
В первой фазе метаболизма ксенобиотиков принимают участие цитохромы группы Р-450. Эти
Слайд 31Эндогенные мутагены
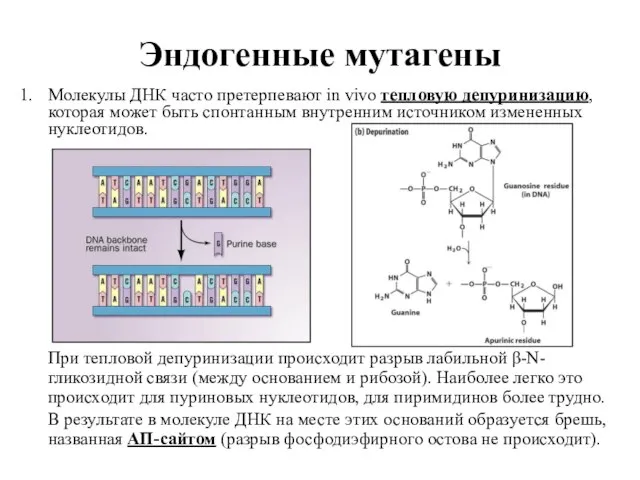
Молекулы ДНК часто претерпевают in vivo тепловую депуринизацию, которая может быть спонтанным
Эндогенные мутагены
Молекулы ДНК часто претерпевают in vivo тепловую депуринизацию, которая может быть спонтанным
Слайд 32Эндогенные мутагены
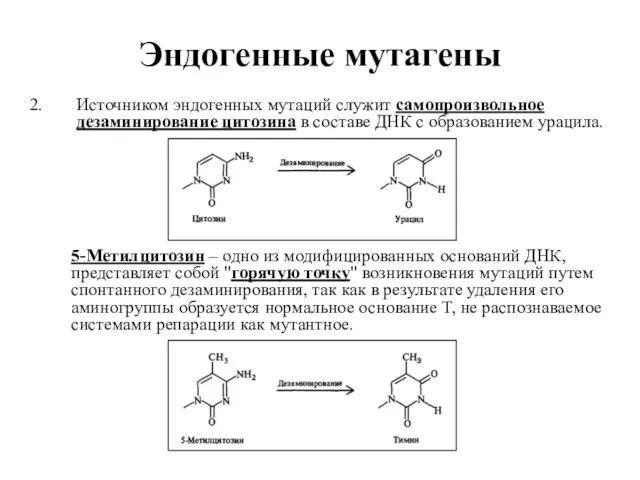
Источником эндогенных мутаций служит самопроизвольное дезаминирование цитозина в составе ДНК с
Эндогенные мутагены
Источником эндогенных мутаций служит самопроизвольное дезаминирование цитозина в составе ДНК с
Слайд 33Эндогенные мутагены
Источником эндогенных мутагенов в организме является метаболизм нормальной микрофлоры. Некоторые промежуточные
Эндогенные мутагены
Источником эндогенных мутагенов в организме является метаболизм нормальной микрофлоры. Некоторые промежуточные

Слайд 34Часть 2
Механизмы репарации
Часть 2
Механизмы репарации

Слайд 35 Репарация генетических повреждений – это свойство живых организмов восстанавливать повреждения, возникшие в
Репарация генетических повреждений – это свойство живых организмов восстанавливать повреждения, возникшие в

Слайд 36Общие сведения
В настоящее время описано множество различных систем репарации, часто принципиально отличных
Общие сведения
В настоящее время описано множество различных систем репарации, часто принципиально отличных

Слайд 37Основной принцип репарации
Основан на двуспиральном строении ДНК.
В большинстве случаев поврежденной оказывается только
Основной принцип репарации
Основан на двуспиральном строении ДНК.
В большинстве случаев поврежденной оказывается только

Слайд 38Повреждения ДНК
Появление различно модифицированных оснований:
Пиримидиновые димеры.
Алкилированые производные.
Дезаминированые основания.
Различные таутомерные формы.
Появление неспаренных оснований
Повреждения ДНК
Появление различно модифицированных оснований:
Пиримидиновые димеры.
Алкилированые производные.
Дезаминированые основания.
Различные таутомерные формы.
Появление неспаренных оснований

Слайд 39Основные повреждающие факторы
Ионизирующие агенты:
Ультрафиолетовый свет.
Радиоактивные вещества.
Активные формы кислорода.
Химические мутагены (например алкилирующие агенты).
Эндогенные
Основные повреждающие факторы
Ионизирующие агенты:
Ультрафиолетовый свет.
Радиоактивные вещества.
Активные формы кислорода.
Химические мутагены (например алкилирующие агенты).
Эндогенные

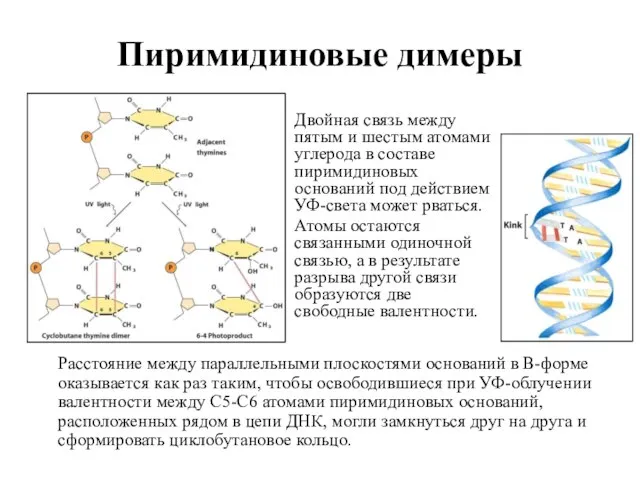
Слайд 40Пиримидиновые димеры
Расстояние между параллельными плоскостями оснований в В-форме оказывается как раз таким,
Пиримидиновые димеры
Расстояние между параллельными плоскостями оснований в В-форме оказывается как раз таким,
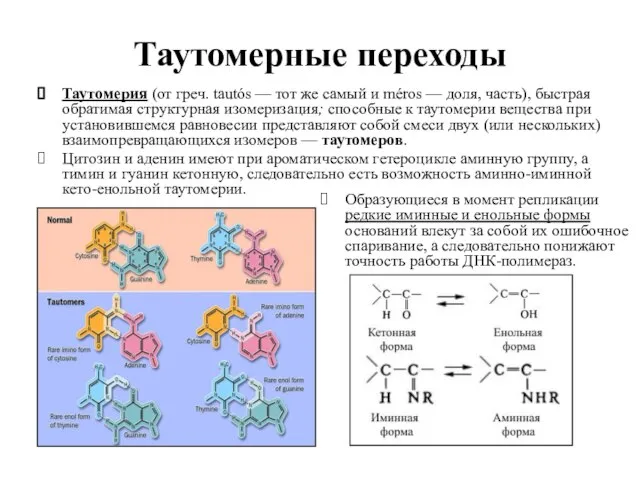
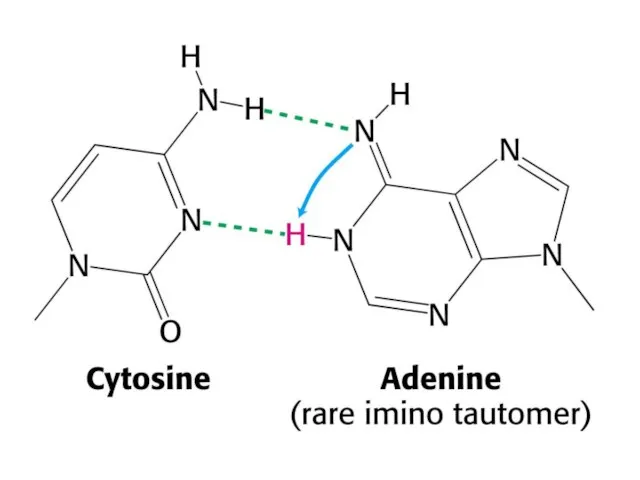
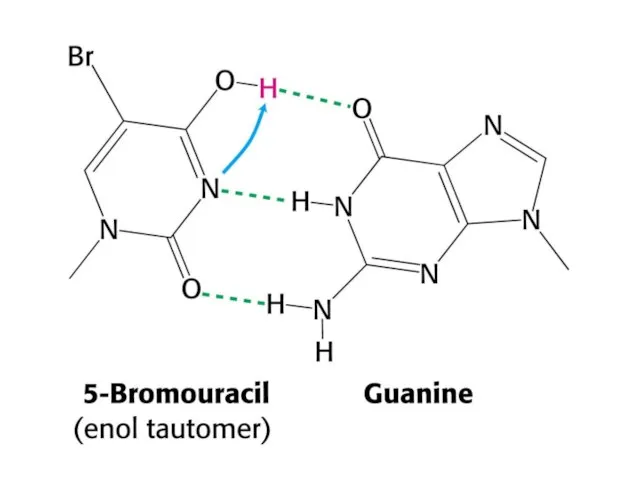
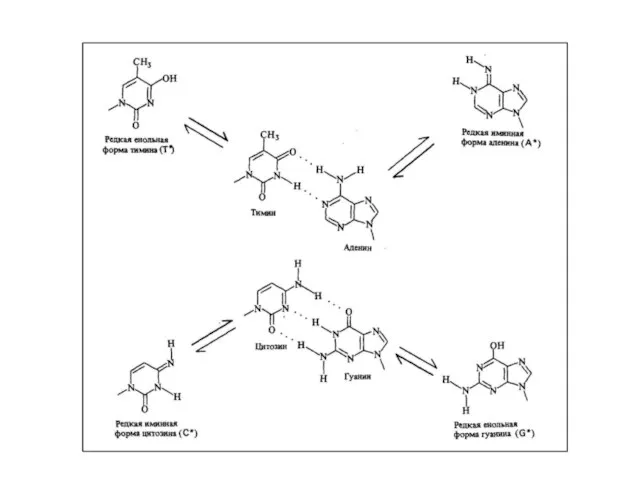
Слайд 41Таутомерные переходы
Таутомерия (от греч. tautós — тот же самый и méros —
Таутомерные переходы
Таутомерия (от греч. tautós — тот же самый и méros —
Слайд 45Разнообразие систем репарации
Существует огромное количество самых различных систем репарации. Все эти системы
Разнообразие систем репарации
Существует огромное количество самых различных систем репарации. Все эти системы

Слайд 46Разнообразие систем репарации
Прямая репарация:
Фотореактивация.
Дезалкилирование модифицированных нуклеотидов.
Сшивание однонитевых разрывов.
Прямая вставка оснований в АП-сайт.
Непрямая
Разнообразие систем репарации
Прямая репарация:
Фотореактивация.
Дезалкилирование модифицированных нуклеотидов.
Сшивание однонитевых разрывов.
Прямая вставка оснований в АП-сайт.
Непрямая

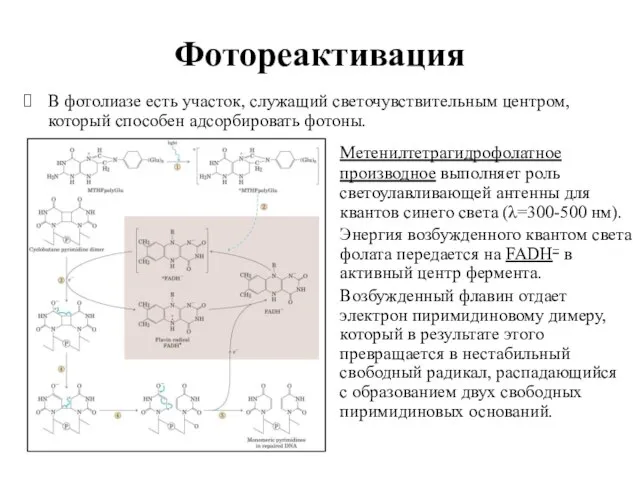
Слайд 47Фотореактивация
В фотолиазе есть участок, служащий светочувствительным центром, который способен адсорбировать фотоны.
Метенилтетрагидрофолатное производное
Фотореактивация
В фотолиазе есть участок, служащий светочувствительным центром, который способен адсорбировать фотоны.
Метенилтетрагидрофолатное производное
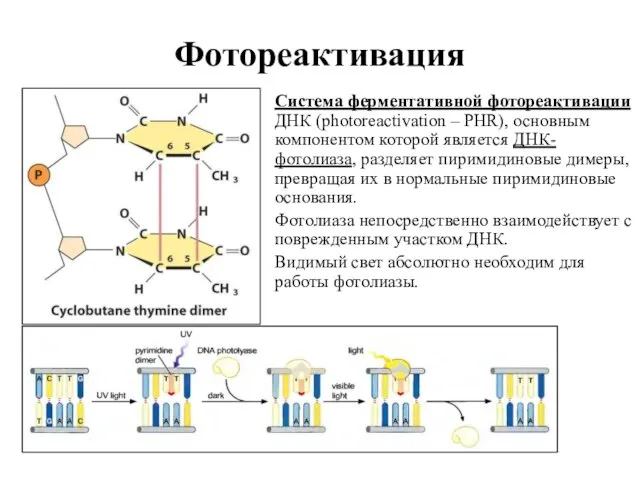
Слайд 48Фотореактивация
Система ферментативной фотореактивации ДНК (photoreactivation – PHR), основным компонентом которой является ДНК-фотолиаза,
Фотореактивация
Система ферментативной фотореактивации ДНК (photoreactivation – PHR), основным компонентом которой является ДНК-фотолиаза,
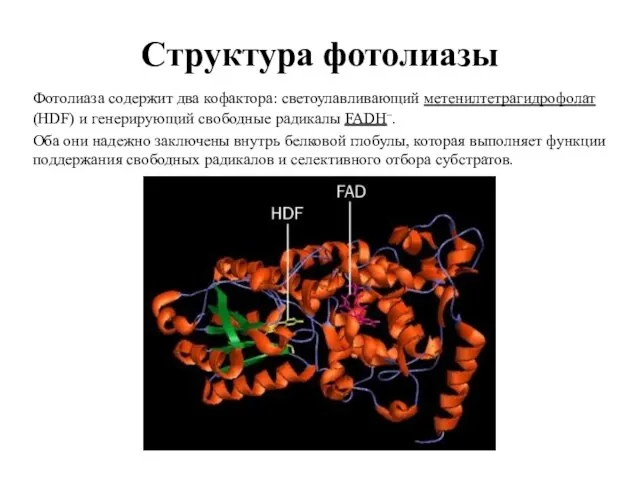
Слайд 49Структура фотолиазы
Фотолиаза содержит два кофактора: светоулавливающий метенилтетрагидрофолат (HDF) и генерирующий свободные радикалы
Структура фотолиазы
Фотолиаза содержит два кофактора: светоулавливающий метенилтетрагидрофолат (HDF) и генерирующий свободные радикалы
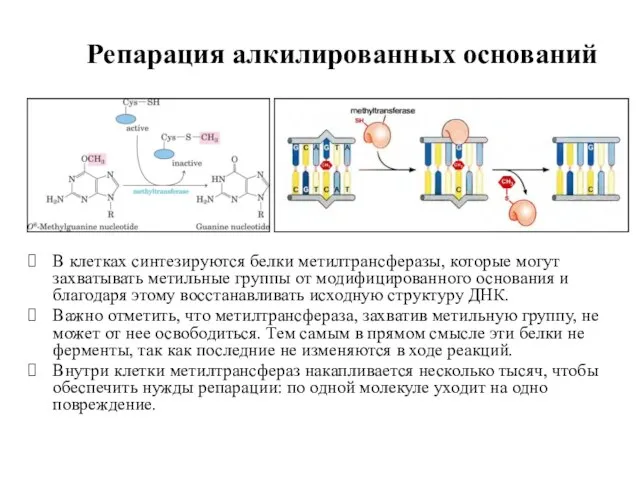
Слайд 50Репарация алкилированных оснований
В клетках синтезируются белки метилтрансферазы, которые могут захватывать метильные группы
Репарация алкилированных оснований
В клетках синтезируются белки метилтрансферазы, которые могут захватывать метильные группы
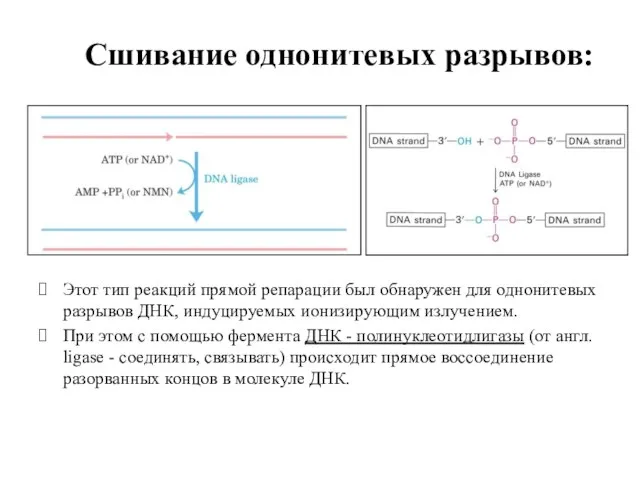
Слайд 51Сшивание однонитевых разрывов:
Этот тип реакций прямой репарации был обнаружен для однонитевых разрывов
Сшивание однонитевых разрывов:
Этот тип реакций прямой репарации был обнаружен для однонитевых разрывов
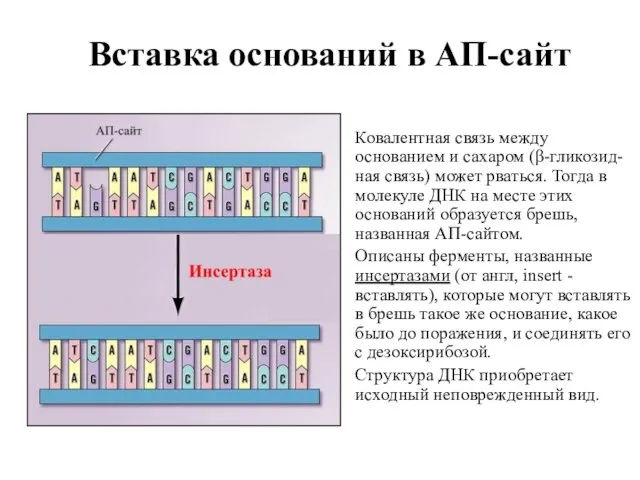
Слайд 52Вставка оснований в АП-сайт
Ковалентная связь между основанием и сахаром (β-гликозид-ная связь) может
Вставка оснований в АП-сайт
Ковалентная связь между основанием и сахаром (β-гликозид-ная связь) может
Слайд 53Эксцизионная репарация ДНК путем удаления поврежденных азотистых оснований (BER)
Эксцизионная репарация ДНК путем удаления поврежденных азотистых оснований (BER)

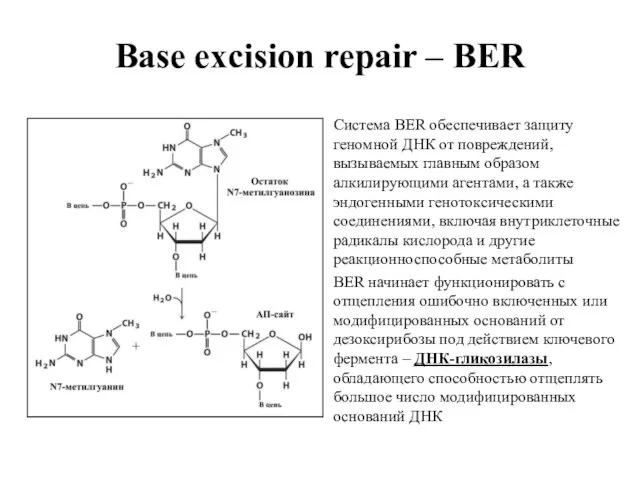
Слайд 54Base excision repair – BER
Система BER обеспечивает защиту геномной ДНК от
Base excision repair – BER
Система BER обеспечивает защиту геномной ДНК от
Слайд 55Разнообразие гликозилаз
Разнообразие гликозилаз

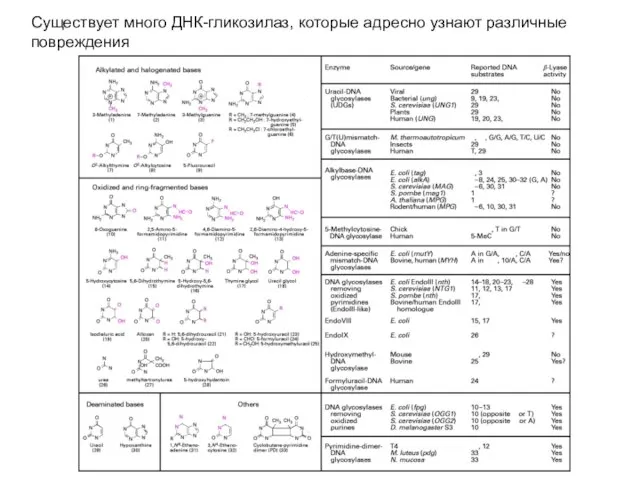
Слайд 56Существует много ДНК-гликозилаз, которые адресно узнают различные
повреждения
Существует много ДНК-гликозилаз, которые адресно узнают различные
повреждения
Слайд 57Структура гликозилаз
Структура гликозилаз

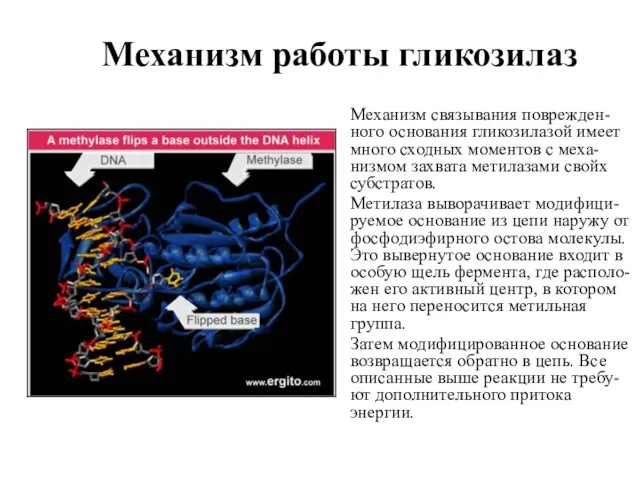
Слайд 58Механизм работы гликозилаз
Механизм связывания поврежден-ного основания гликозилазой имеет много сходных моментов с
Механизм работы гликозилаз
Механизм связывания поврежден-ного основания гликозилазой имеет много сходных моментов с
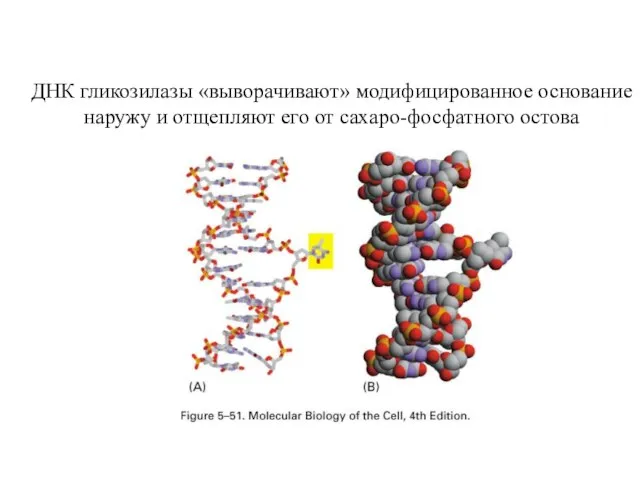
Слайд 59ДНК гликозилазы «выворачивают» модифицированное основание
наружу и отщепляют его от сахаро-фосфатного остова
ДНК гликозилазы «выворачивают» модифицированное основание
наружу и отщепляют его от сахаро-фосфатного остова
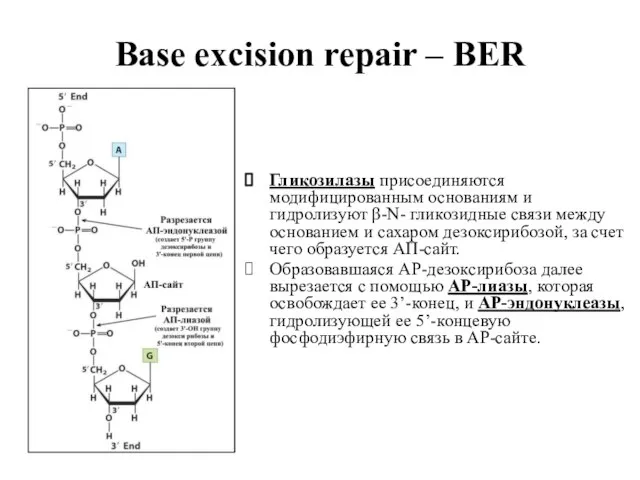
Слайд 60Base excision repair – BER
Гликозилазы присоединяются модифицированным основаниям и гидролизуют β-N- гликозидные
Base excision repair – BER
Гликозилазы присоединяются модифицированным основаниям и гидролизуют β-N- гликозидные
Слайд 61Base excision repair – BER
Появившаяся брешь в одной цепи ДНК размером в
Base excision repair – BER
Появившаяся брешь в одной цепи ДНК размером в

Слайд 62Эксцизионная репарация ДНК путем удаления нуклеотидов (NER)
Эксцизионная репарация ДНК путем удаления нуклеотидов (NER)

Слайд 63Nucleotide excision repair – NER
Процесс NER условно можно разделить на четыре
Nucleotide excision repair – NER
Процесс NER условно можно разделить на четыре

Слайд 64Nucleotide excision repair – NER
В отличии от BER, субстратами системы NER являются
Nucleotide excision repair – NER
В отличии от BER, субстратами системы NER являются

Слайд 65Механизм работы
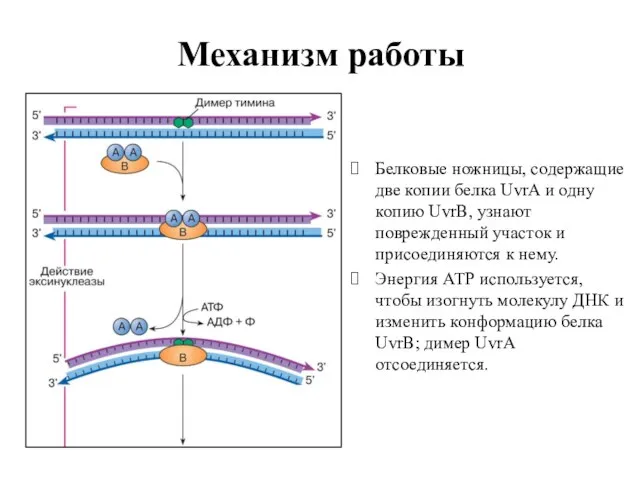
Белковые ножницы, содержащие две копии белка UvrA и одну копию UvrB,
Механизм работы
Белковые ножницы, содержащие две копии белка UvrA и одну копию UvrB,
Слайд 66Механизм работы
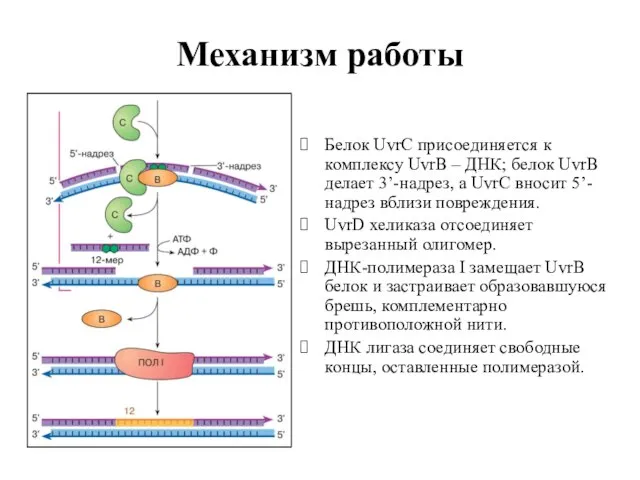
Белок UvrC присоединяется к комплексу UvrВ – ДНК; белок UvrВ делает
Механизм работы
Белок UvrC присоединяется к комплексу UvrВ – ДНК; белок UvrВ делает
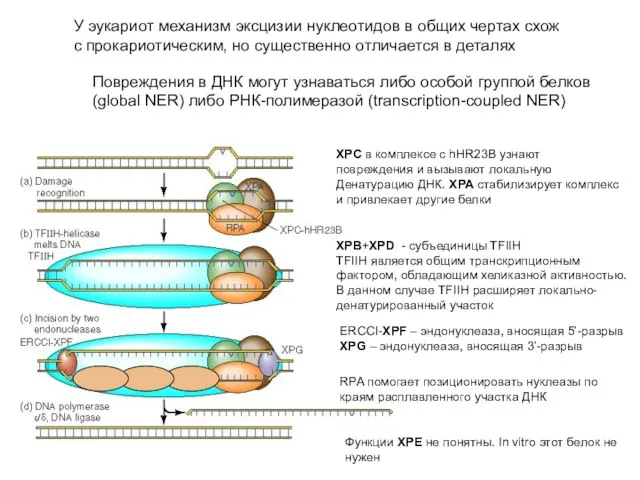
Слайд 67У эукариот механизм эксцизии нуклеотидов в общих чертах схож
с прокариотическим, но
У эукариот механизм эксцизии нуклеотидов в общих чертах схож
с прокариотическим, но
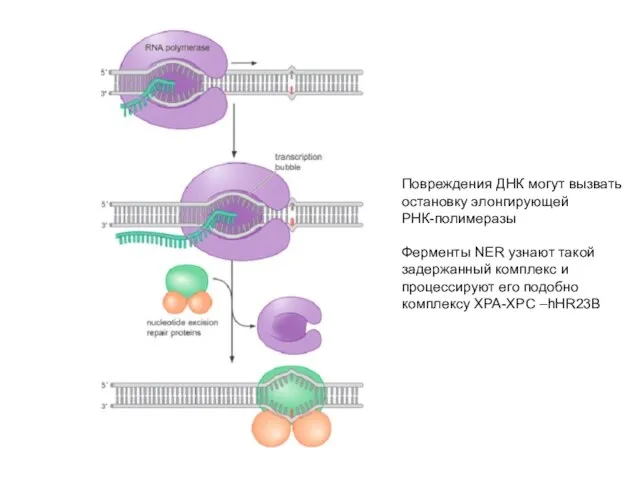
Слайд 68Повреждения ДНК могут вызвать
остановку элонгирующей
РНК-полимеразы
Ферменты NER узнают такой
задержанный комплекс и
Повреждения ДНК могут вызвать
остановку элонгирующей
РНК-полимеразы
Ферменты NER узнают такой
задержанный комплекс и
Слайд 69XPB & XPD – TFII H DNA helicase
XPC - damage recognition
XPA stabilisation
XPB & XPD – TFII H DNA helicase
XPC - damage recognition
XPA stabilisation
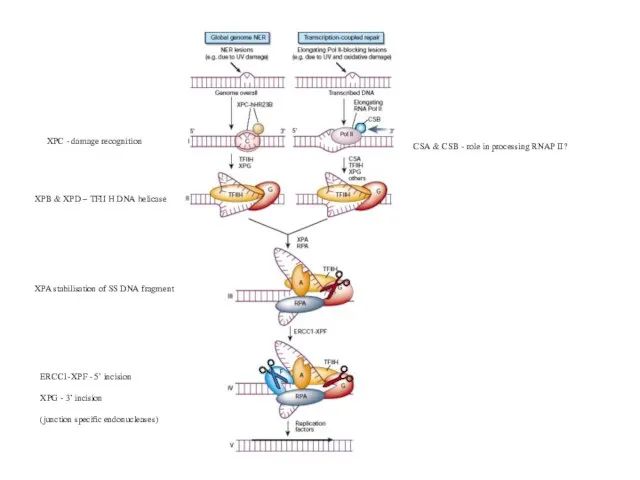
Слайд 70Различия NER у про- и эукариот
Гены NER у E. coli uvrA, uvrB и
Различия NER у про- и эукариот
Гены NER у E. coli uvrA, uvrB и

Слайд 71Репарация ошибочно спаренных нуклеотидов (MMR)
Репарация ошибочно спаренных нуклеотидов (MMR)

Слайд 72Mismatch repair - MMR
В отличие от NER, так же удаляющей неправильно спаренные
Mismatch repair - MMR
В отличие от NER, так же удаляющей неправильно спаренные

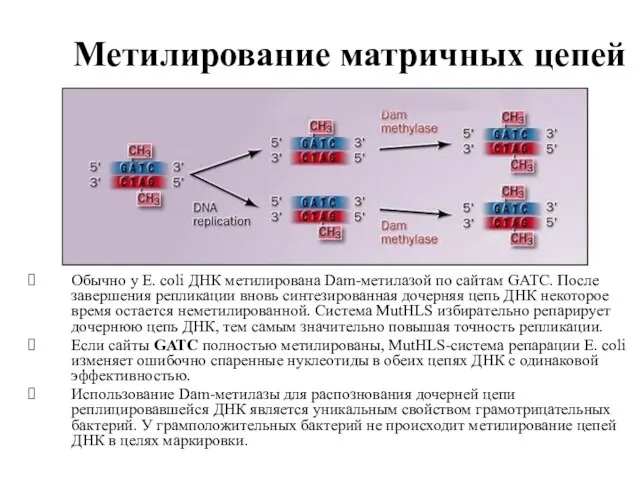
Слайд 73Метилирование матричных цепей
Обычно у E. coli ДНК метилирована Dam-метилазой по сайтам GATC. После
Метилирование матричных цепей
Обычно у E. coli ДНК метилирована Dam-метилазой по сайтам GATC. После
Слайд 74Механизм работы
На начальных этапах система MMR задействует белковые продукты четырех генов: mutH,
Механизм работы
На начальных этапах система MMR задействует белковые продукты четырех генов: mutH,
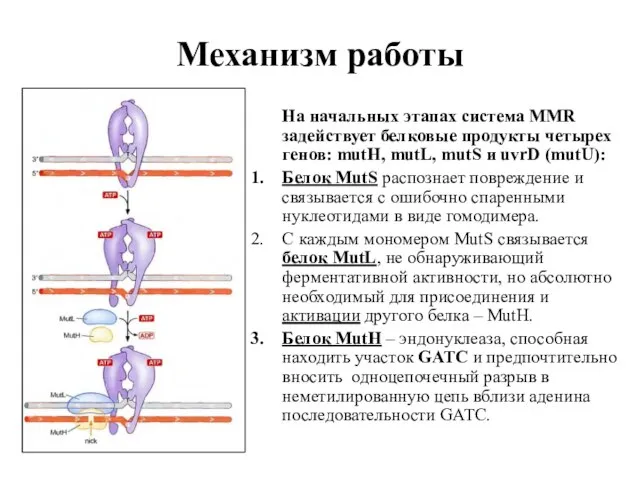
Слайд 75Механизм работы
После полной сборки комплекса MutHLS, активации эндонуклеазной активности MutH и внесения
Механизм работы
После полной сборки комплекса MutHLS, активации эндонуклеазной активности MutH и внесения
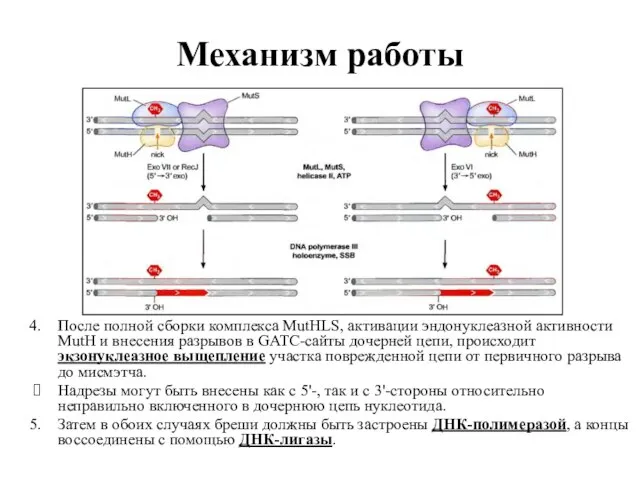
Слайд 76Другие системы
У E. coli существуют два других специфических пути репарации ошибочно спаренных нуклеотидов:
Система
Другие системы
У E. coli существуют два других специфических пути репарации ошибочно спаренных нуклеотидов:
Система

Слайд 77гомологи MutS (MSH — MutS homolog) образуют два гетеродимерных комплекса
-- MSH2-MSH6
гомологи MutS (MSH — MutS homolog) образуют два гетеродимерных комплекса
-- MSH2-MSH6

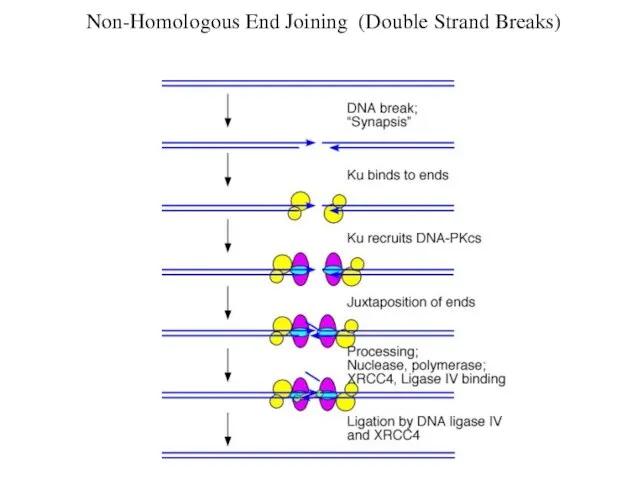
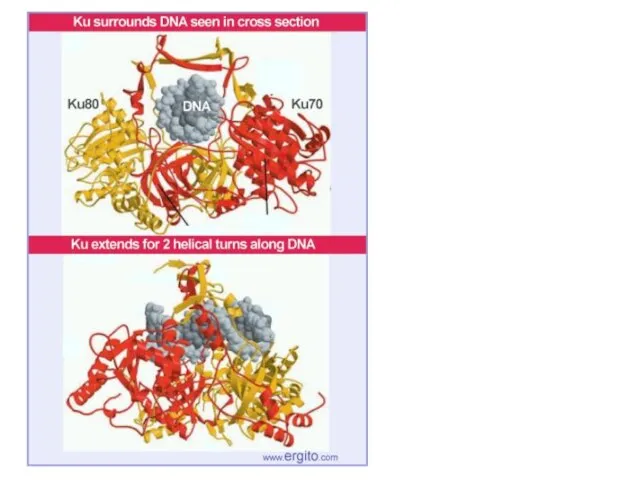
Слайд 78Non-Homologous End Joining (Double Strand Breaks)
Non-Homologous End Joining (Double Strand Breaks)
Слайд 80Пострепликативная (рекомбинационная) репарация
Пострепликативная (рекомбинационная) репарация

Слайд 81Рекомбинационная репарация
Быстро делящиеся бактериальные клетки, содержащие несколько репликонов, образованных недореплицированными хромосомами, более
Рекомбинационная репарация
Быстро делящиеся бактериальные клетки, содержащие несколько репликонов, образованных недореплицированными хромосомами, более

Слайд 82Рекомбинационная репарация
Рекомбинационная репарация необходима в случае, когда повреждены обе цепи ДНК, что
Рекомбинационная репарация
Рекомбинационная репарация необходима в случае, когда повреждены обе цепи ДНК, что

Слайд 83Рекомбинационная репарация
Второй дуплекс ДНК, полученный в результате репликации, не содержит в себе
Рекомбинационная репарация
Второй дуплекс ДНК, полученный в результате репликации, не содержит в себе

Слайд 84Механизм процесса
Главный этап рекомбинационной репарации заключается в активации генов rec, участвующих так
Механизм процесса
Главный этап рекомбинационной репарации заключается в активации генов rec, участвующих так

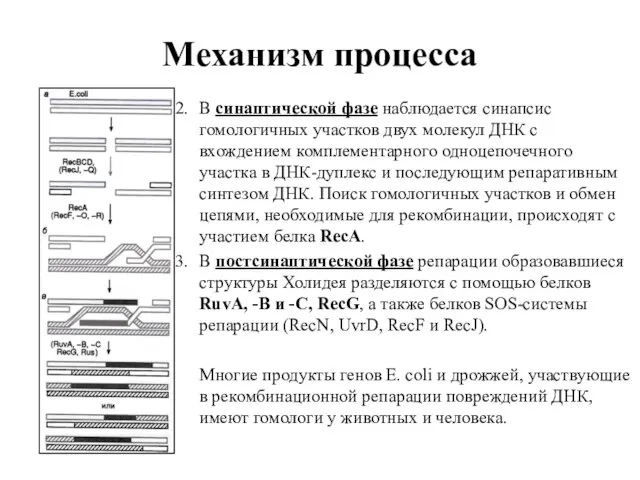
Слайд 85Механизм процесса
В синаптической фазе наблюдается синапсис гомологичных участков двух молекул ДНК с
Механизм процесса
В синаптической фазе наблюдается синапсис гомологичных участков двух молекул ДНК с
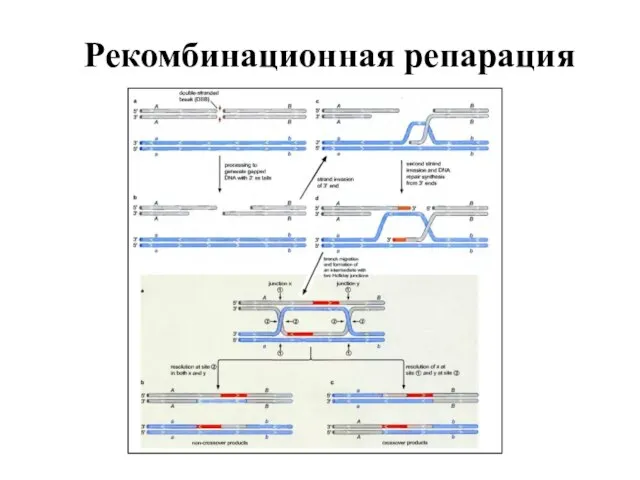
Слайд 86Рекомбинационная репарация
Рекомбинационная репарация
Слайд 87Часть 3
SOS-репарация и SOS-мутагенез
Часть 3
SOS-репарация и SOS-мутагенез

Слайд 88SOS-репарация
В клетках организма, подвергнутого сильному мутагенному воздействию, образование мутаций происходит в основном
SOS-репарация
В клетках организма, подвергнутого сильному мутагенному воздействию, образование мутаций происходит в основном

Слайд 89Индукция SOS-репарации
Белок LexA является репрессором гена recA и более 20 других генов
Индукция SOS-репарации
Белок LexA является репрессором гена recA и более 20 других генов
Слайд 90Активация белка RecA
Точно еще не выяснено как индукции SOS-сигнала, как и его
Активация белка RecA
Точно еще не выяснено как индукции SOS-сигнала, как и его

Слайд 91Индукция SOS-репарации
Молекулы белка LexA взаимодействуют с RecA* в результате чего активируется скрытая
Индукция SOS-репарации
Молекулы белка LexA взаимодействуют с RecA* в результате чего активируется скрытая
Слайд 92Основные термины
SOS-репарация (мутагенез) – это согласованная индукция множества ферментов, включая ферменты репарации,
Основные термины
SOS-репарация (мутагенез) – это согласованная индукция множества ферментов, включая ферменты репарации,

Слайд 93Индукция SOS-репарации
Ключевыми белками, задействоваными в процессе введения ошибок ДНК-полимеразой, являются продукты двух
Индукция SOS-репарации
Ключевыми белками, задействоваными в процессе введения ошибок ДНК-полимеразой, являются продукты двух

Слайд 94ДНК-полимераза 5
Настоящая роль белкового комплекса (UmuD’)2–UmuC в процессе SOS-репарации неизвестна. Существует две
ДНК-полимераза 5
Настоящая роль белкового комплекса (UmuD’)2–UmuC в процессе SOS-репарации неизвестна. Существует две

Слайд 95Индукция SOS-репарации
Когда репликаза (ДНК-полимераза 3) доходит до некоторого повреждения, например тиминового димера,
Индукция SOS-репарации
Когда репликаза (ДНК-полимераза 3) доходит до некоторого повреждения, например тиминового димера,

Слайд 96 Таким образом белок RecA играет ключевую роль в индукции SOS-репарации, выполняя следующие
Таким образом белок RecA играет ключевую роль в индукции SOS-репарации, выполняя следующие

Слайд 97ДНК-полимераза 4
ДНК-полимераза IV E. coli, кодируемая геном dinB, также участвует в SOS-ответе бактерий:
Она не
ДНК-полимераза 4
ДНК-полимераза IV E. coli, кодируемая геном dinB, также участвует в SOS-ответе бактерий:
Она не

Слайд 98После того, как весь геном был успешно реплицирован, путем введения мутации в
После того, как весь геном был успешно реплицирован, путем введения мутации в

Слайд 99Вырезание профагов
Активация RecA так же может являться причиной расщепления и других репрессорных
Вырезание профагов
Активация RecA так же может являться причиной расщепления и других репрессорных

Слайд 100Прежде всего об остановке репликации надо сообщить
Rec A
Cвязывается с однонитевой
ДНК и образует
Прежде всего об остановке репликации надо сообщить
Rec A
Cвязывается с однонитевой
ДНК и образует

Слайд 101 lexA, recA, uvrA, uvrB, and uvrD
Филаменты Rec A стимулируют аутопротеолиз Lex
lexA, recA, uvrA, uvrB, and uvrD
Филаменты Rec A стимулируют аутопротеолиз Lex

Слайд 103UmuD повергается автопротеолитическому расщеплению с образованием
активного фрагмента UmuD’
UmuD’ активирует черезблоковую полимеразу UmuC
комплекс
UmuD повергается автопротеолитическому расщеплению с образованием
активного фрагмента UmuD’
UmuD’ активирует черезблоковую полимеразу UmuC
комплекс


 Тип Кишечнополостные животные
Тип Кишечнополостные животные Сервировка стола к завтраку
Сервировка стола к завтраку Архитектурные постройки
Архитектурные постройки Макияж при близорукости, дальнозоркости, для тех, кто носит линзы
Макияж при близорукости, дальнозоркости, для тех, кто носит линзы الخطأ السياقي
الخطأ السياقي Презентация по обществознанию на тему _Мировоззрение и его роль в жизни человека_ (10
Презентация по обществознанию на тему _Мировоззрение и его роль в жизни человека_ (10 Spotlight
Spotlight Создай образ с TRESemme
Создай образ с TRESemme Linkin park
Linkin park Презентация на тему Степные животные
Презентация на тему Степные животные Сочинение –описание животного.Моя собака Крош.
Сочинение –описание животного.Моя собака Крош. Предпринимательский модуль. Мастерская агробизнеса
Предпринимательский модуль. Мастерская агробизнеса Обзор методологии Scrum Auriga Inc. Дмитрий Сидоренко. - презентация
Обзор методологии Scrum Auriga Inc. Дмитрий Сидоренко. - презентация Обращение к психологу, страшно или нет
Обращение к психологу, страшно или нет Салон штор и гардин LINENS™
Салон штор и гардин LINENS™ Непроектное мероприятие «Образование детей-инвалидов»
Непроектное мероприятие «Образование детей-инвалидов» Гимнастика. Виды гимнастики
Гимнастика. Виды гимнастики Балет - один из языков истории
Балет - один из языков истории Отзывы о Чувышии
Отзывы о Чувышии Сигналы тревоги.
Сигналы тревоги. Олимпиаде в Сочи посвящается… Кроссворд Олимпийские игры
Олимпиаде в Сочи посвящается… Кроссворд Олимпийские игры Современный дизайн. Место дизайна в современном мире. Тренды в графическом дизайне
Современный дизайн. Место дизайна в современном мире. Тренды в графическом дизайне Время Present Perfect
Время Present Perfect Бережливое управление МБУ УБГБ (1)
Бережливое управление МБУ УБГБ (1) Лавис и резерваж. Манеры офорта
Лавис и резерваж. Манеры офорта Презентация на тему Фотосинтез и дыхание растений (6 класс)
Презентация на тему Фотосинтез и дыхание растений (6 класс)  Страна под настроение
Страна под настроение Создание системы бюджетного управления
Создание системы бюджетного управления