- Эффект кворума

Содержание
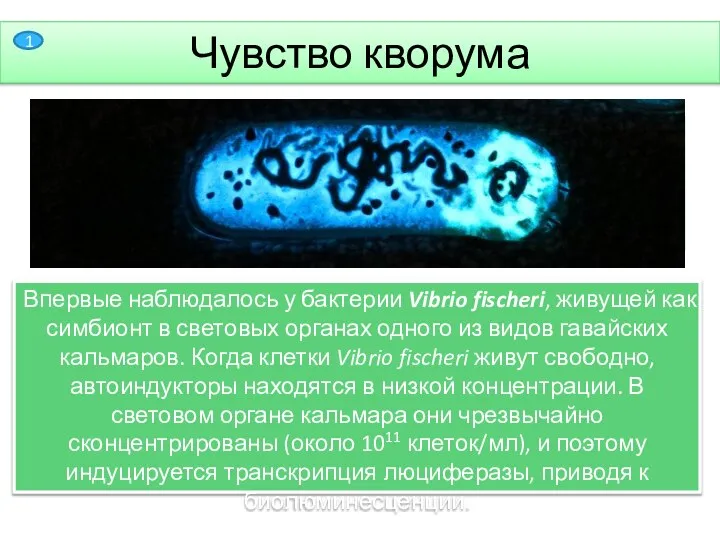
- 2. Чувство кворума Чувство кворума— способность некоторых микроорганизмов «общаться» и координировать своё поведение за счёт секреции молекулярных
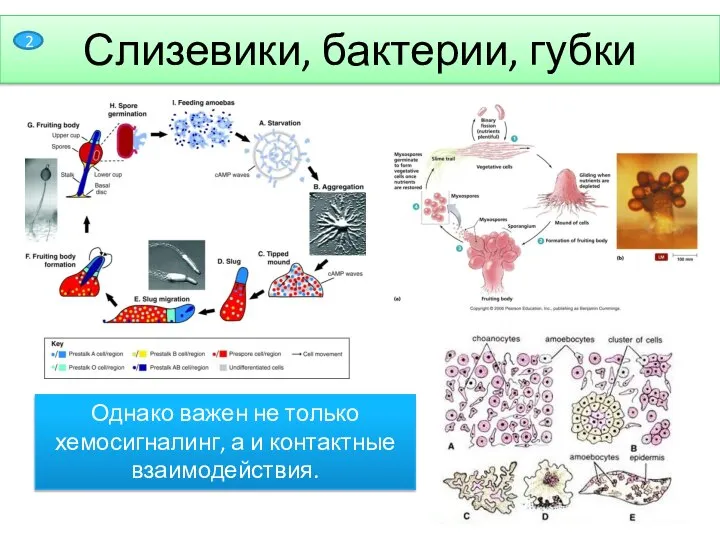
- 3. Слизевики, бактерии, губки Однако важен не только хемосигналинг, а и контактные взаимодействия. 2
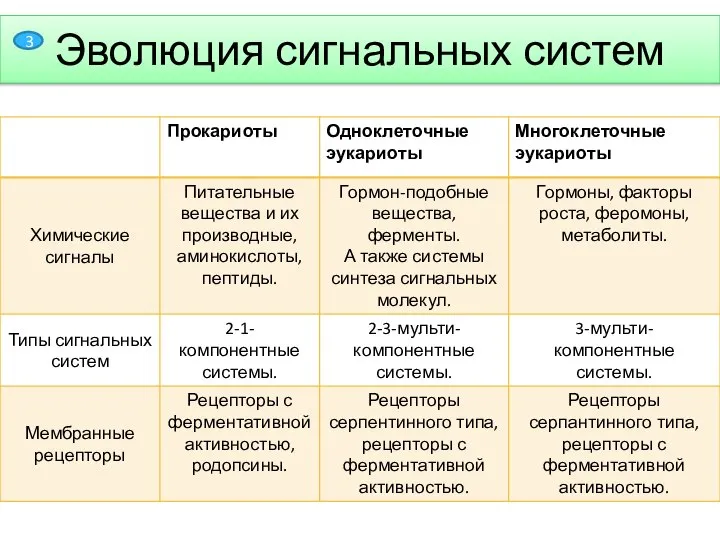
- 4. Эволюция сигнальных систем 3
- 5. До недавнишнего времени считалось, что хемосигналинг прокариот и эукариот не имеет ничего общего Геномы прокариот и
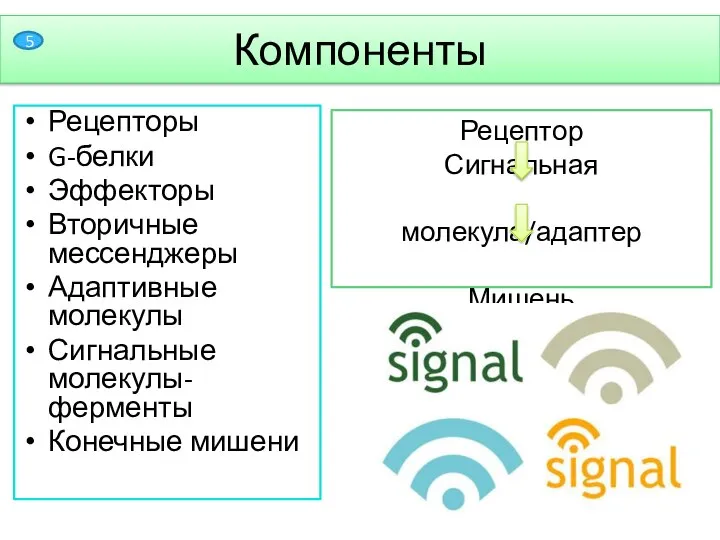
- 6. Рецепторы G-белки Эффекторы Вторичные мессенджеры Адаптивные молекулы Сигнальные молекулы-ферменты Конечные мишени Рецептор Сигнальная молекула/адаптер Мишень Компоненты
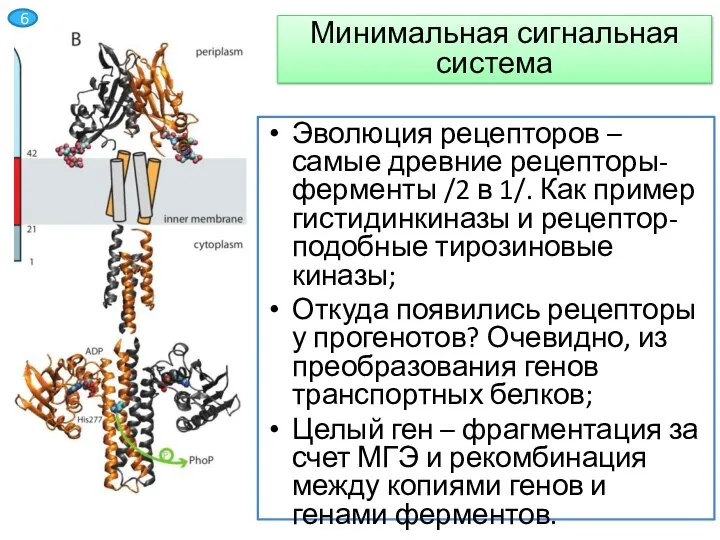
- 7. Эволюция рецепторов – самые древние рецепторы-ферменты /2 в 1/. Как пример гистидинкиназы и рецептор-подобные тирозиновые киназы;
- 8. ЭВОЛЮЦИЯ Ген транспортного белка Рецепторный участок Домен локализации Эволюция генов Ген ферментов киназ Каталитический промежуток, которые
- 9. Каталитический домен Субстрат-распознающий домен Присоединение остатка органической кислоты к аминокислоте Новые измененные вещества, но сходные со
- 10. Несколько этапов становления многоклеточности 840-740 млн лет назад появились первые эволюционные пробы многоклеточности - хайнаньская биота
- 11. R Пептиды, АК R Участки белка R Углеводы R Полисахариды R Гликопротеины, гликолипиды Рекомбинация генов Контактные
- 12. Кроме хемосигналинга многоклеточным организмам необходимы клеточные взаимодействия; Примитивная сигнальная система может состоять из 2-3 компонентов; Самыми
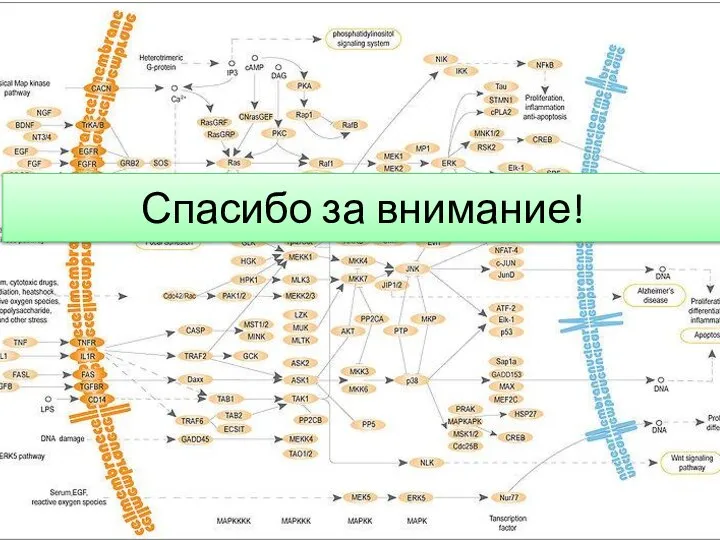
- 13. Спасибо за внимание!
- 15. Скачать презентацию
Слайд 2Чувство кворума
Чувство кворума— способность некоторых микроорганизмов «общаться» и координировать своё поведение за
Чувство кворума
Чувство кворума— способность некоторых микроорганизмов «общаться» и координировать своё поведение за
Слайд 3Слизевики, бактерии, губки
Однако важен не только хемосигналинг, а и контактные взаимодействия.
2
Слизевики, бактерии, губки
Однако важен не только хемосигналинг, а и контактные взаимодействия.
2
Слайд 4Эволюция сигнальных систем
3
Эволюция сигнальных систем
3
Слайд 5До недавнишнего времени считалось, что хемосигналинг прокариот и эукариот не имеет ничего
До недавнишнего времени считалось, что хемосигналинг прокариот и эукариот не имеет ничего

Слайд 6Рецепторы
G-белки
Эффекторы
Вторичные мессенджеры
Адаптивные молекулы
Сигнальные молекулы-ферменты
Конечные мишени
Рецептор
Сигнальная молекула/адаптер
Мишень
Компоненты
5
Рецепторы
G-белки
Эффекторы
Вторичные мессенджеры
Адаптивные молекулы
Сигнальные молекулы-ферменты
Конечные мишени
Рецептор
Сигнальная молекула/адаптер
Мишень
Компоненты
5
Слайд 7Эволюция рецепторов – самые древние рецепторы-ферменты /2 в 1/. Как пример гистидинкиназы
Эволюция рецепторов – самые древние рецепторы-ферменты /2 в 1/. Как пример гистидинкиназы
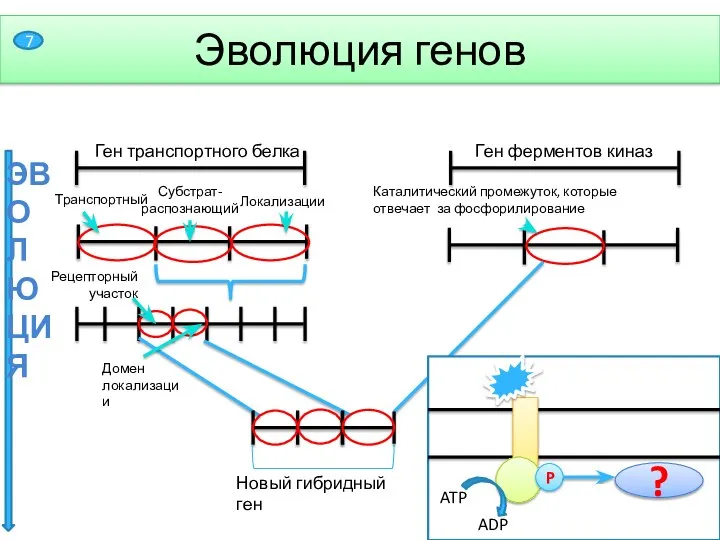
Слайд 8ЭВОЛЮЦИЯ
Ген транспортного белка
Рецепторный участок
Домен локализации
Эволюция генов
Ген ферментов киназ
Каталитический промежуток, которые отвечает за
ЭВОЛЮЦИЯ
Ген транспортного белка
Рецепторный участок
Домен локализации
Эволюция генов
Ген ферментов киназ
Каталитический промежуток, которые отвечает за
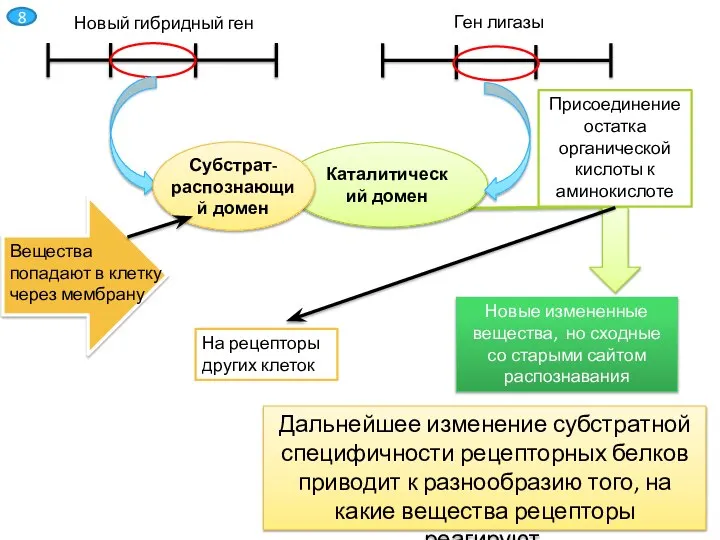
Слайд 9Каталитический домен
Субстрат-распознающий домен
Присоединение остатка органической кислоты к аминокислоте
Новые измененные вещества, но сходные
Каталитический домен
Субстрат-распознающий домен
Присоединение остатка органической кислоты к аминокислоте
Новые измененные вещества, но сходные
Слайд 10Несколько этапов становления многоклеточности
840-740 млн лет назад появились первые эволюционные пробы многоклеточности
Несколько этапов становления многоклеточности
840-740 млн лет назад появились первые эволюционные пробы многоклеточности

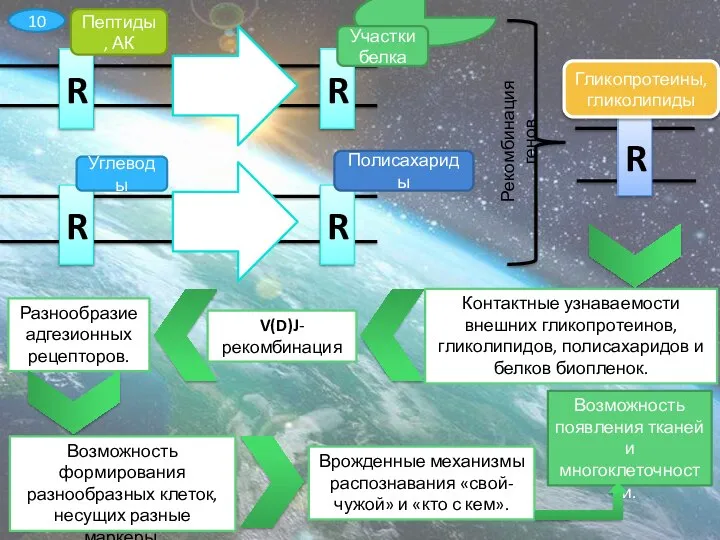
Слайд 11R
Пептиды, АК
R
Участки белка
R
Углеводы
R
Полисахариды
R
Гликопротеины, гликолипиды
Рекомбинация генов
Контактные узнаваемости внешних гликопротеинов, гликолипидов, полисахаридов и белков
R
Пептиды, АК
R
Участки белка
R
Углеводы
R
Полисахариды
R
Гликопротеины, гликолипиды
Рекомбинация генов
Контактные узнаваемости внешних гликопротеинов, гликолипидов, полисахаридов и белков
Слайд 12Кроме хемосигналинга многоклеточным организмам необходимы клеточные взаимодействия;
Примитивная сигнальная система может состоять из
Кроме хемосигналинга многоклеточным организмам необходимы клеточные взаимодействия;
Примитивная сигнальная система может состоять из

Слайд 13Спасибо за внимание!
Спасибо за внимание!
 По воде на лыжах. Клопы-водомерки
По воде на лыжах. Клопы-водомерки Биологический кросссворд
Биологический кросссворд Температура, как экологический фактор
Температура, как экологический фактор Палеонтологические доказательства эволюции
Палеонтологические доказательства эволюции Ромашка аптечная (ромашка лекарственная) Matricaria charnomilla L. (M. recutita L.)
Ромашка аптечная (ромашка лекарственная) Matricaria charnomilla L. (M. recutita L.) Сукцессия. Саморазвитие экосистемы
Сукцессия. Саморазвитие экосистемы Физиология возбудимых тканей: общие вопросы. Физиология биологических мембран
Физиология возбудимых тканей: общие вопросы. Физиология биологических мембран Все ли йогурты полезны?
Все ли йогурты полезны? Фотосинтез
Фотосинтез Органические вещества
Органические вещества Опорно-двигательная система
Опорно-двигательная система Презентация на тему Методы селекции растений
Презентация на тему Методы селекции растений  Kā darbojas sirds? Kas izraisa sirds saraušanos?
Kā darbojas sirds? Kas izraisa sirds saraušanos? Основы генетики
Основы генетики Клетка живого организма
Клетка живого организма Презентация на тему Семейство Бобовые и Пасленовые
Презентация на тему Семейство Бобовые и Пасленовые  Строение цветкового растения
Строение цветкового растения Бактерии и вирусы
Бактерии и вирусы Қоршаған орта факторларының екіншілік ИТЖ дамуындағы маңызы
Қоршаған орта факторларының екіншілік ИТЖ дамуындағы маңызы Вьюрковые
Вьюрковые Тварина Австралії - намбат
Тварина Австралії - намбат СОН
СОН Обеспечение клеток энергией
Обеспечение клеток энергией Слуховой анализатор и орган равновесия
Слуховой анализатор и орган равновесия Класс Птицы. Общая характеристика класса
Класс Птицы. Общая характеристика класса Типы размножения
Типы размножения Этапы формирования и развития представлений о клетке
Этапы формирования и развития представлений о клетке Способы размножения водорослей
Способы размножения водорослей