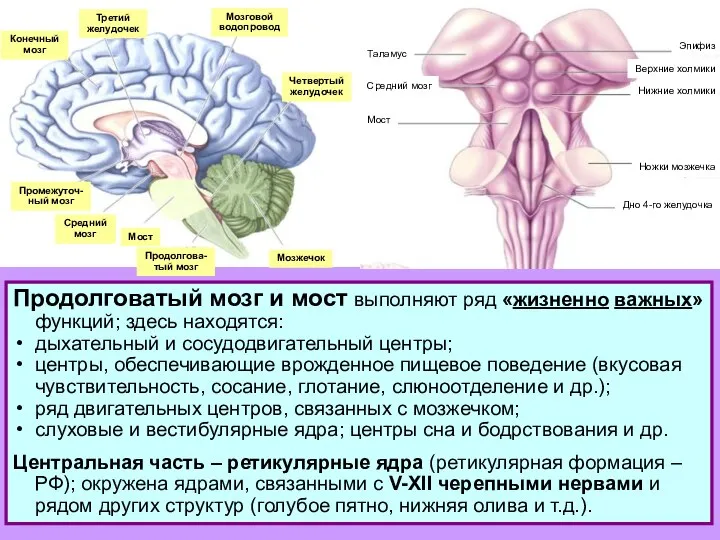
Продолговатый мозг и мост: дыхательный и сосудодвигательный центры; проведение вкусовых, слуховых и вестибулярных сигналов
- Продолговатый мозг и мост: дыхательный и сосудодвигательный центры; проведение вкусовых, слуховых и вестибулярных сигналов

Содержание
- 2. Конечный мозг Третий желудочек Мозговой водопровод Четвертый желудочек Мозжечок Промежуточ- ный мозг Средний мозг Мост Продолгова-
- 3. Четверохолмие – анализ сенсорных сигналов, запуск реакции на новые стимулы (ориентировочный рефлекс – поворот глаз, головы
- 5. Схема расположения дыхательных нейронов продолг. мозга и моста: I – инспираторные (вдох), Е – экспираторные (выдох).
- 6. От клеток-пейсмекеров (генераторов ритма) ПД передаются к другим дыхат. нейронам и мотонейронам шейных и грудных сегментов
- 7. Еще о дыхательных центрах: инспираторные нейроны – это не только пейсмекеры, но и клетки, «зацикливающие» ПД
- 8. Дыхательная кривая (объем грудной клетки) в норме, при метаболичес- ком ацидозе («закислении») и некоторых видах патологии.
- 9. Нервная регуляция сердечных сокращений: 1, 2 – сосудодвигательный центр продолговатого мозга и моста и поступающие из
- 10. Рефлекс Гольтца Рефлекс Даньини-Ашнера Реакции возни-кают за счет распространения сигналов от рецепторов растяжения (например, брыжейки) и
- 11. «норма» Х нерв симп. нерв Барорецепторный рефлекс – компенсаторная реакция на изменение растяжения стенок дуги аорты
- 12. Дыхательной аритмии подвержена активность как симпатических, так и парасимпатических нер-вов, однако только дейст-вие Ацх развивается и
- 13. Основные связи сосудо-двигательного центра продолговатого мозга и моста (на выходе показаны только симпат. эффекты): 1. Барорецепторы
- 14. Вкусовые центры продолговатого мозга и моста (сигналы от языка VII и IX н.; от глотки –
- 15. Вестибулярные и слуховые ядра – по углам ромбовидной ямки; вестиб. ядра ме-диальнее, как эволю-ционно более древние.
- 16. Вестибулярная информация (информация о положении тела в пространстве) необходима для оперативной коррекции движений; в связи с
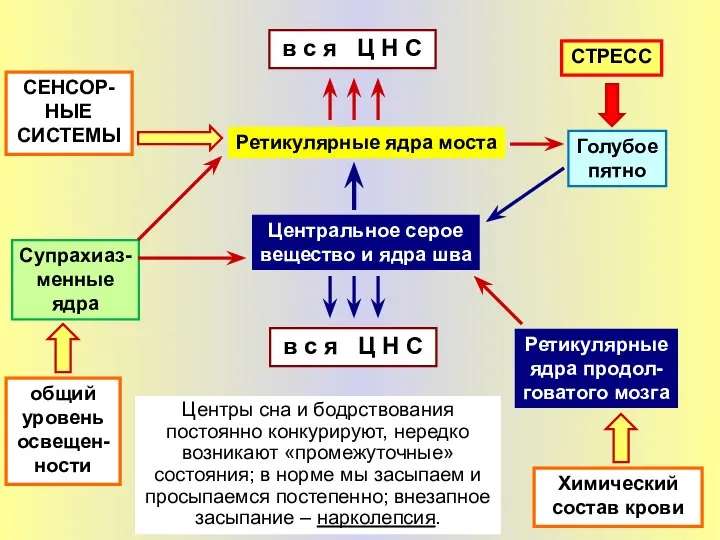
- 17. Центры сна и бодрствования. Эволюционно очень древние, постоянно конкурируют друг с другом, учитывают значительное число факторов
- 18. 1. Главный центр бодрствования: ретикулярные ядра моста; сюда поступает часть информации от всех сенсорных систем; далее
- 19. 3. Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2] за счет выделения NE.
- 20. 4. Супрахиазменные ядра переднего гипоталамуса: находятся напротив перекреста зрительных нервов, получают информацию об общем уровне освещенности
- 21. Но все еще сложнее, и во время сна выделяют стадии (на основе анализа ЭЭГ): Бодрствование: альфа-ритм
- 22. Но все еще сложнее, и во время сна выделяют стадии (на основе анализа ЭЭГ): Бодрствование: альфа-ритм
- 23. REM: rapid eye movement СНЫ – «окно в бессознательное», «дефрагментация диска», продолжение ментальных процессов в ином
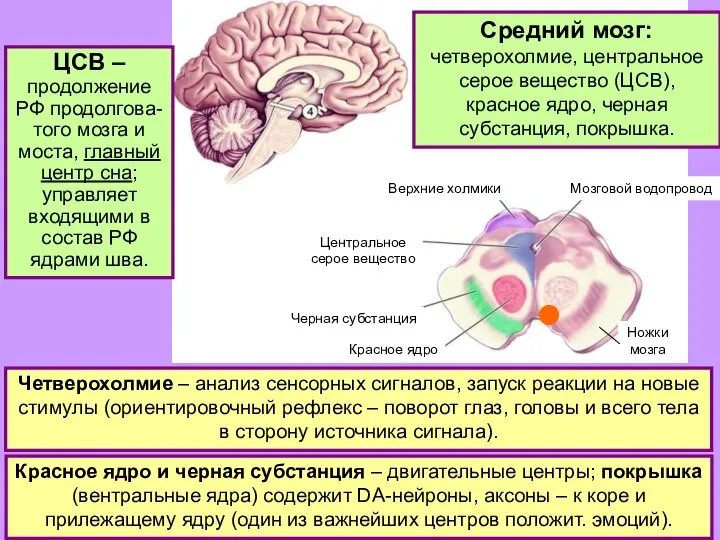
- 24. СРЕДНИЙ МОЗГ. Центральное серое вещество: собирает большое число информационных потоков и через ядра шва влияет на
- 25. Четверохолмие: реакция на новизну; верхние холмики – на новые зрительные стимулы; нижние холмики – на новые
- 26. Движения глаз: с каждым глазом связано по 6 мышц, управляемых III, IV и VI нервами; два
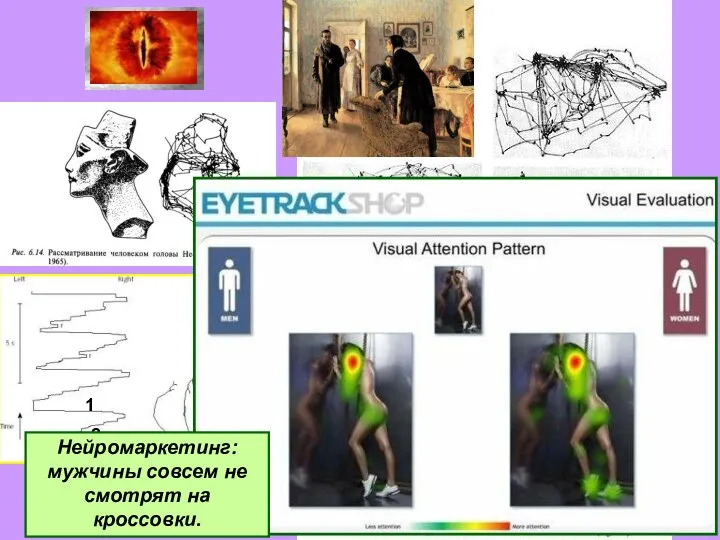
- 27. Нейромаркетинг: мужчины совсем не смотрят на кроссовки.
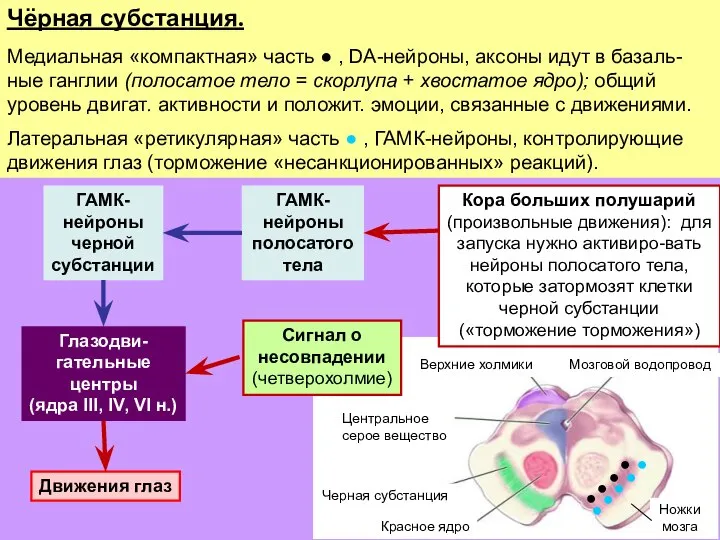
- 28. Чёрная субстанция. Медиальная «компактная» часть ● , DA-нейроны, аксоны идут в базаль-ные ганглии (полосатое тело =
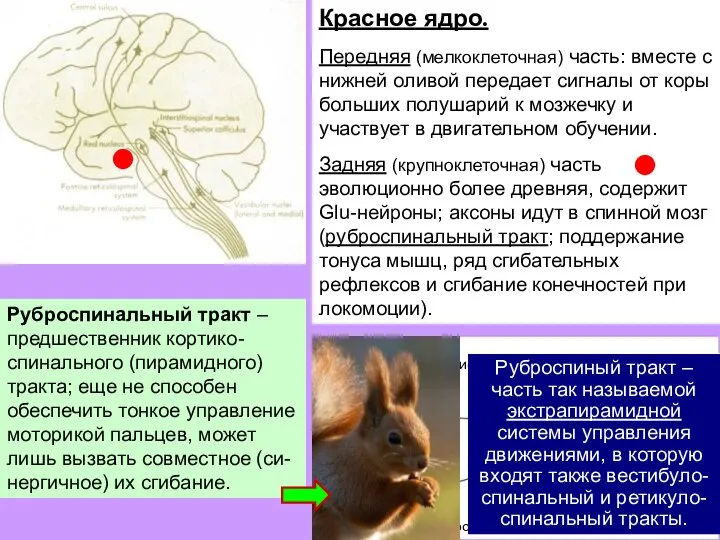
- 29. Красное ядро. Передняя (мелкоклеточная) часть: вместе с нижней оливой передает сигналы от коры больших полушарий к
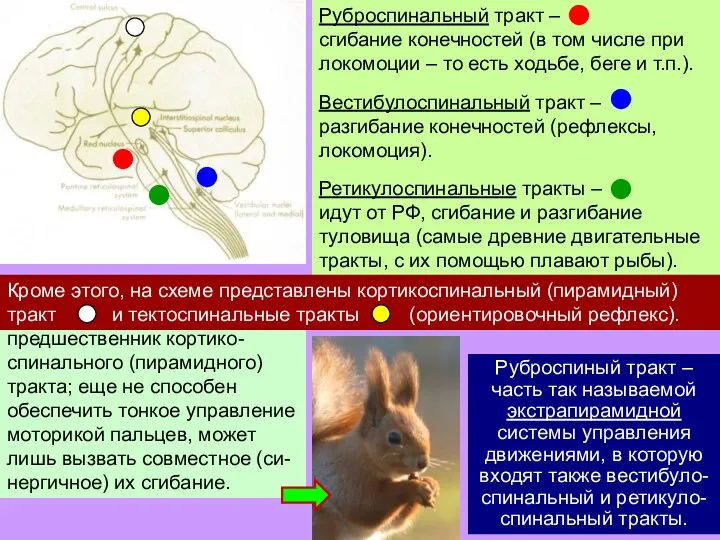
- 30. Руброспиный тракт – часть так называемой экстрапирамидной системы управления движениями, в которую входят также вестибуло-спинальный и
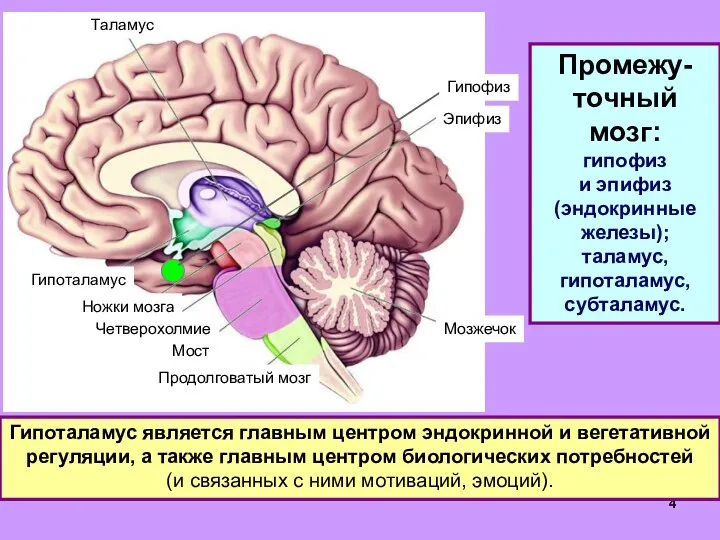
- 31. Гипоталамус и терморегуляция. В передней части гипоталамуса (преоптическая область) – нейроны-терморецепторы, постоянно измеряют температуру крови, 80%
- 32. Закаливание – тренировка систем терморегуляции; снижает вероятность простудных заболеваний. У животных – особые органы теплоотдачи (хвосты,
- 34. Скачать презентацию
Слайд 2Конечный
мозг
Третий
желудочек
Мозговой
водопровод
Четвертый
желудочек
Мозжечок
Промежуточ-
ный мозг
Средний
мозг
Мост
Продолгова-
тый мозг
Продолговатый
мозг и мост:
ядра черепных нервов +
ретикулярные ядра
Конечный
мозг
Третий
желудочек
Мозговой
водопровод
Четвертый
желудочек
Мозжечок
Промежуточ-
ный мозг
Средний
мозг
Мост
Продолгова-
тый мозг
Продолговатый
мозг и мост:
ядра черепных нервов +
ретикулярные ядра
Слайд 3Четверохолмие – анализ сенсорных сигналов, запуск реакции на новые стимулы (ориентировочный рефлекс
Четверохолмие – анализ сенсорных сигналов, запуск реакции на новые стимулы (ориентировочный рефлекс
Слайд 5Схема расположения дыхательных нейронов продолг. мозга и моста:
I – инспираторные (вдох),
Е
Схема расположения дыхательных нейронов продолг. мозга и моста:
I – инспираторные (вдох),
Е
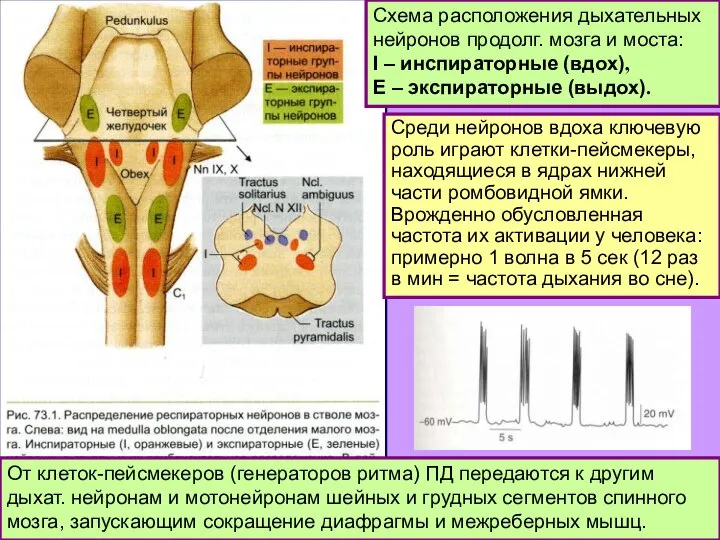
Слайд 6От клеток-пейсмекеров (генераторов ритма) ПД передаются к другим дыхат. нейронам и мотонейронам
От клеток-пейсмекеров (генераторов ритма) ПД передаются к другим дыхат. нейронам и мотонейронам
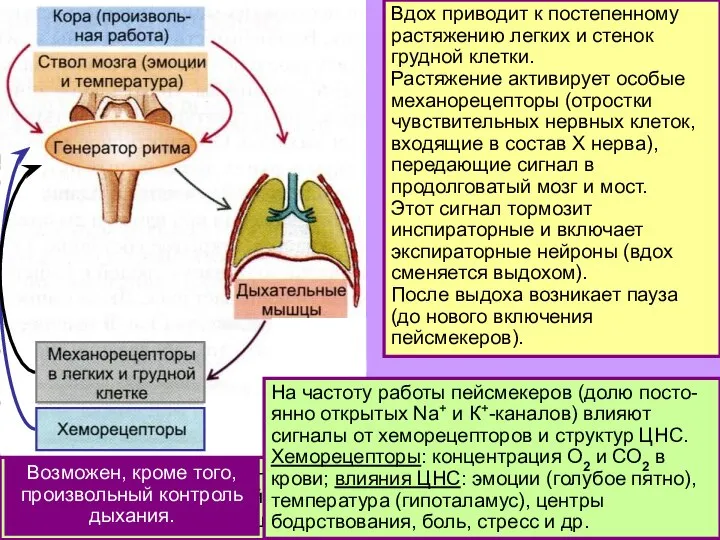
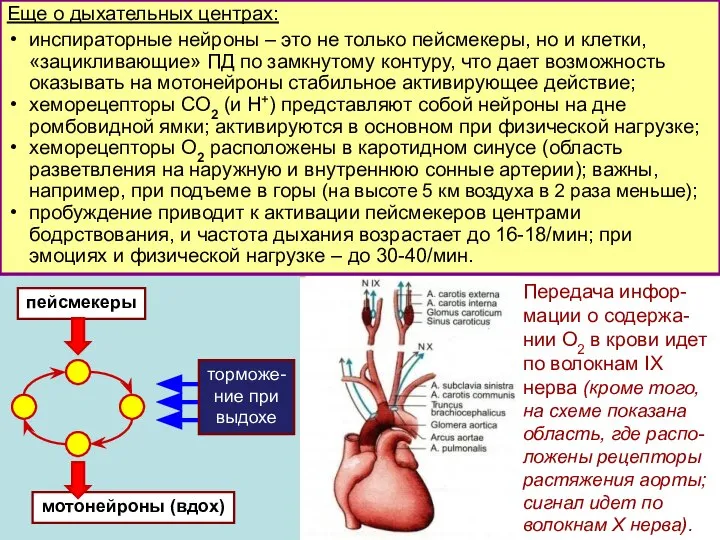
Слайд 7Еще о дыхательных центрах:
инспираторные нейроны – это не только пейсмекеры, но и
Еще о дыхательных центрах:
инспираторные нейроны – это не только пейсмекеры, но и
Слайд 8Дыхательная кривая (объем грудной
клетки) в норме, при метаболичес-
ком ацидозе («закислении») и
Дыхательная кривая (объем грудной
клетки) в норме, при метаболичес-
ком ацидозе («закислении») и

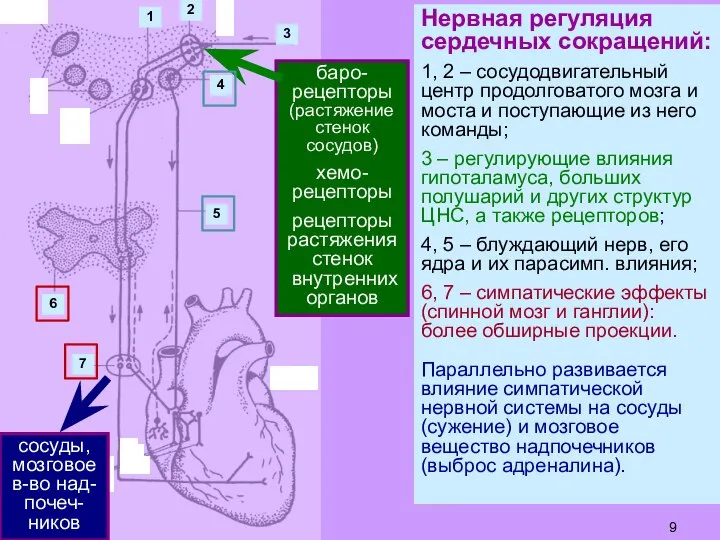
Слайд 9Нервная регуляция сердечных сокращений:
1, 2 – сосудодвигательный центр продолговатого мозга и моста
Нервная регуляция сердечных сокращений:
1, 2 – сосудодвигательный центр продолговатого мозга и моста
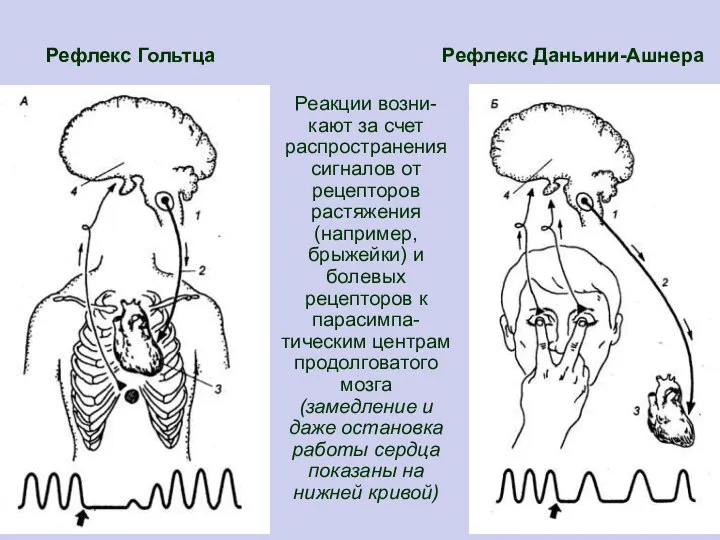
Слайд 10 Рефлекс Гольтца Рефлекс Даньини-Ашнера
Реакции возни-кают за счет распространения сигналов от рецепторов
Рефлекс Гольтца Рефлекс Даньини-Ашнера
Реакции возни-кают за счет распространения сигналов от рецепторов
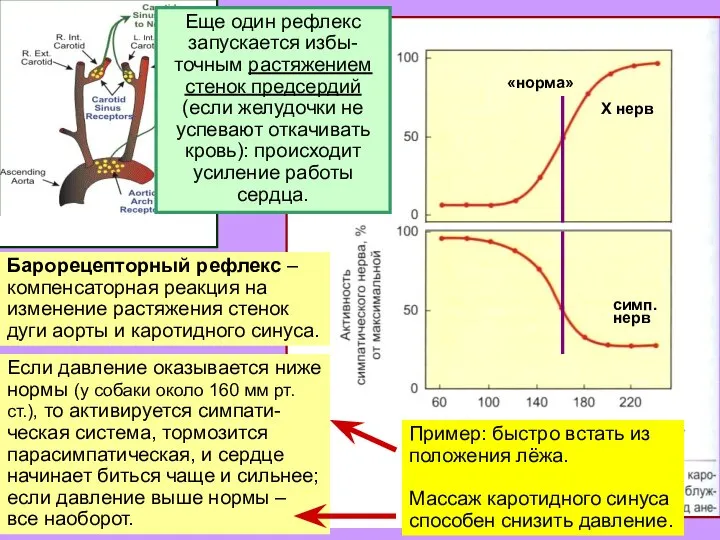
Слайд 11«норма»
Х нерв
симп.
нерв
Барорецепторный рефлекс –
компенсаторная реакция на изменение растяжения стенок дуги аорты
«норма»
Х нерв
симп.
нерв
Барорецепторный рефлекс –
компенсаторная реакция на изменение растяжения стенок дуги аорты
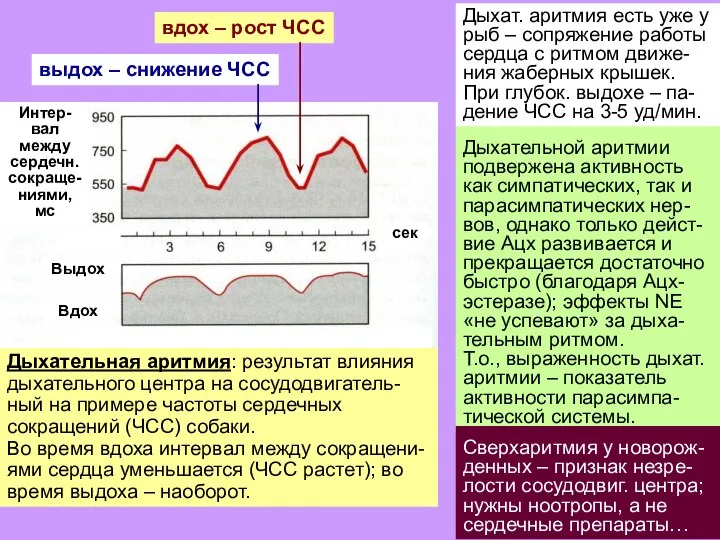
Слайд 12Дыхательной аритмии
подвержена активность как симпатических, так и парасимпатических нер-вов, однако только дейст-вие
подвержена активность как симпатических, так и парасимпатических нер-вов, однако только дейст-вие
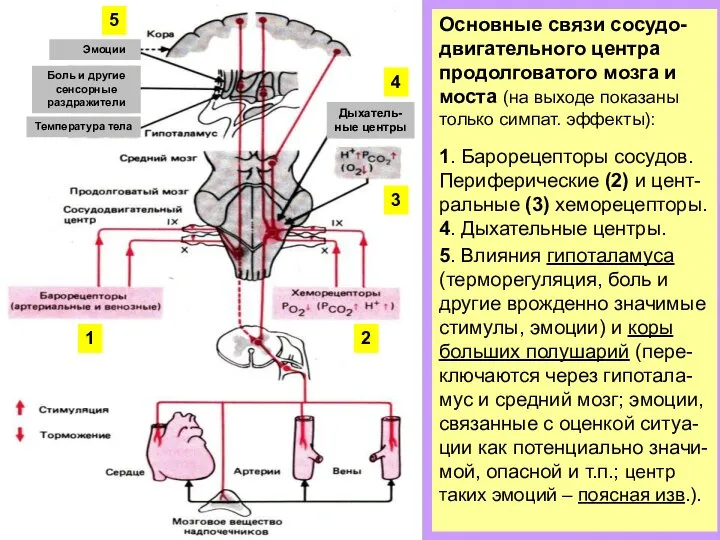
Слайд 13Основные связи сосудо-двигательного центра продолговатого мозга и моста (на выходе показаны только
Основные связи сосудо-двигательного центра продолговатого мозга и моста (на выходе показаны только
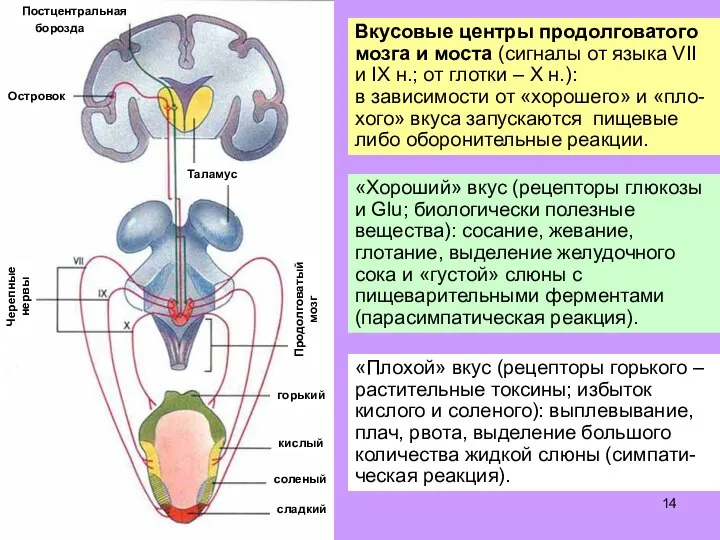
Слайд 14Вкусовые центры продолговатого мозга и моста (сигналы от языка VII и IX
Вкусовые центры продолговатого мозга и моста (сигналы от языка VII и IX
Слайд 15Вестибулярные и слуховые ядра –
по углам ромбовидной ямки; вестиб. ядра ме-диальнее,
Вестибулярные и слуховые ядра –
по углам ромбовидной ямки; вестиб. ядра ме-диальнее,
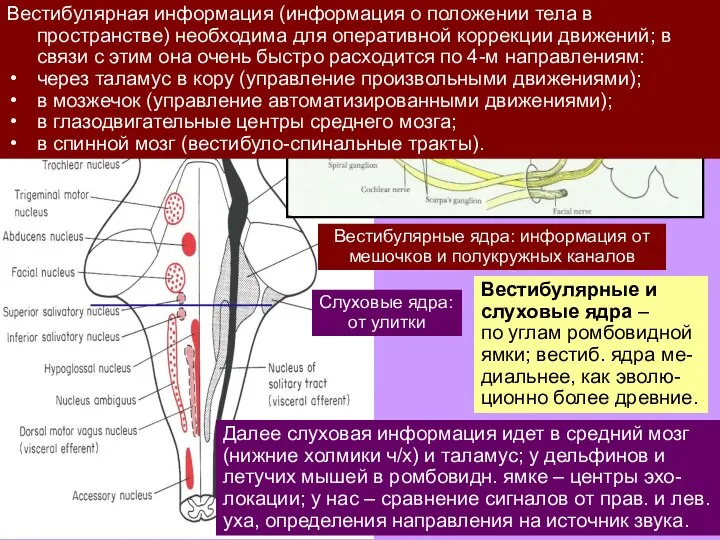
Слайд 16Вестибулярная информация (информация о положении тела в пространстве) необходима для оперативной коррекции
Вестибулярная информация (информация о положении тела в пространстве) необходима для оперативной коррекции

Слайд 17Центры сна и бодрствования.
Эволюционно очень древние, постоянно конкурируют друг с другом,
Центры сна и бодрствования.
Эволюционно очень древние, постоянно конкурируют друг с другом,
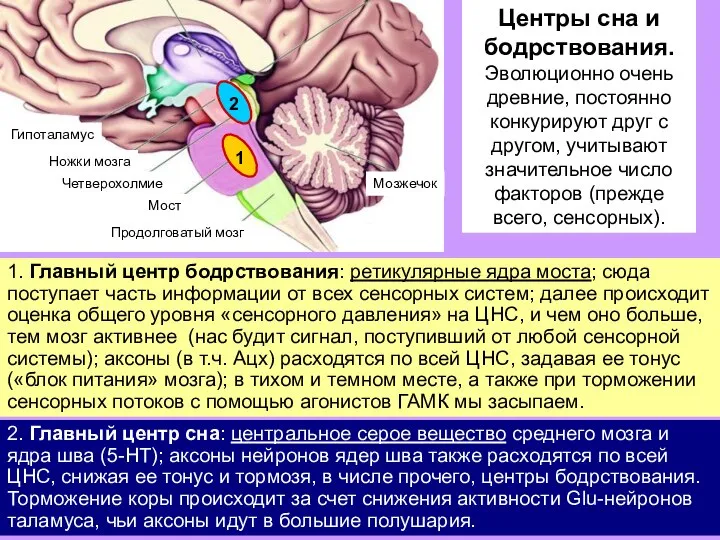
Слайд 181. Главный центр бодрствования: ретикулярные ядра моста; сюда поступает часть информации от
1. Главный центр бодрствования: ретикулярные ядра моста; сюда поступает часть информации от
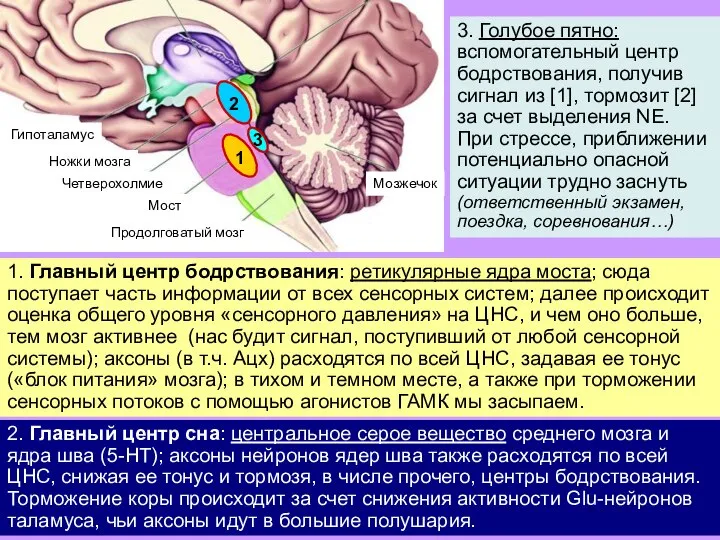
Слайд 193. Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]
3. Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]
![3. Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/1046280/slide-18.jpg)
Слайд 204. Супрахиазменные ядра переднего гипоталамуса: находятся напротив перекреста зрительных нервов, получают информацию
4. Супрахиазменные ядра переднего гипоталамуса: находятся напротив перекреста зрительных нервов, получают информацию
Слайд 21Но все еще сложнее, и во время сна выделяют стадии (на основе
Но все еще сложнее, и во время сна выделяют стадии (на основе
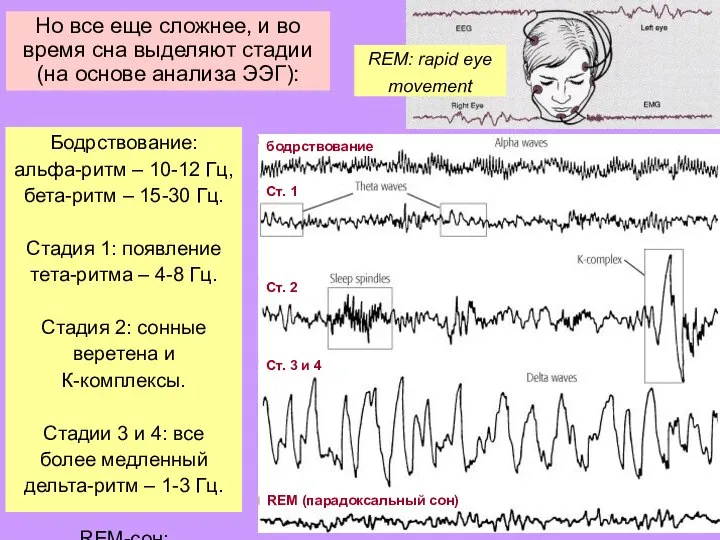
Слайд 22Но все еще сложнее, и во время сна выделяют стадии (на основе
Но все еще сложнее, и во время сна выделяют стадии (на основе
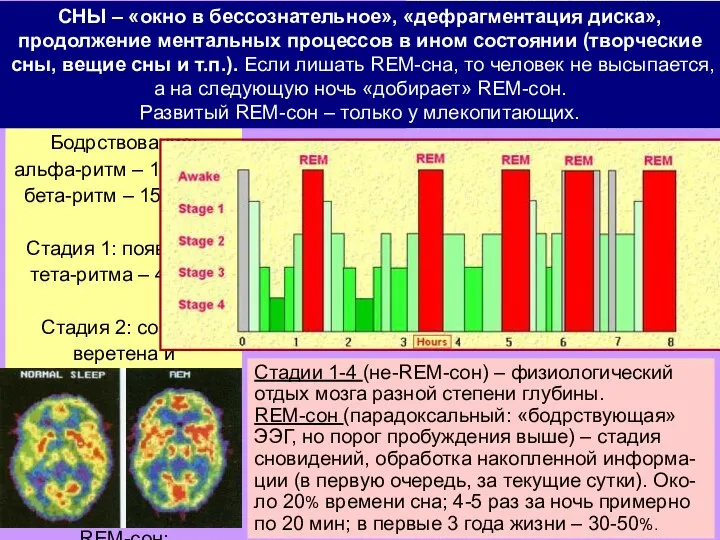
Слайд 23REM: rapid eye movement
СНЫ – «окно в бессознательное», «дефрагментация диска», продолжение ментальных
REM: rapid eye movement
СНЫ – «окно в бессознательное», «дефрагментация диска», продолжение ментальных

Слайд 24СРЕДНИЙ МОЗГ.
Центральное серое вещество:
собирает большое число информационных потоков и через ядра шва
СРЕДНИЙ МОЗГ.
Центральное серое вещество:
собирает большое число информационных потоков и через ядра шва
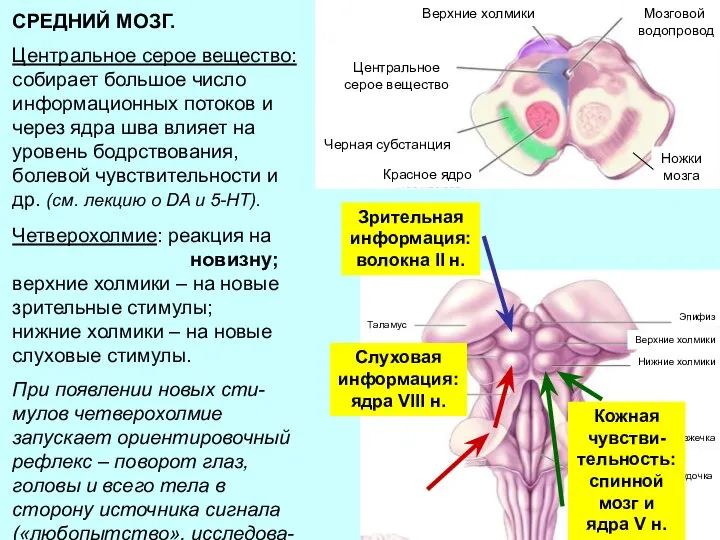
Слайд 25Четверохолмие: реакция на
новизну;
верхние холмики – на новые зрительные стимулы;
нижние холмики
новизну;
верхние холмики – на новые зрительные стимулы;
нижние холмики

Слайд 26Движения глаз:
с каждым глазом связано по 6 мышц, управляемых III, IV и
Движения глаз:
с каждым глазом связано по 6 мышц, управляемых III, IV и
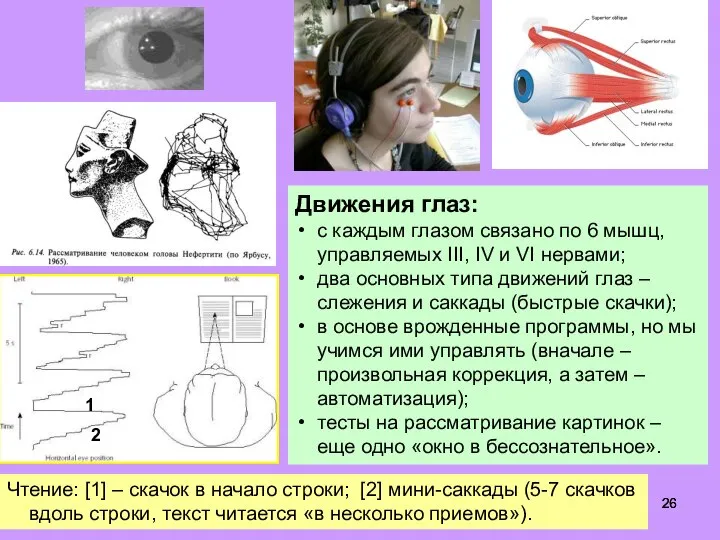
Слайд 27Нейромаркетинг: мужчины совсем не смотрят на кроссовки.
Нейромаркетинг: мужчины совсем не смотрят на кроссовки.
Слайд 28Чёрная субстанция.
Медиальная «компактная» часть ● , DA-нейроны, аксоны идут в базаль-ные ганглии
Чёрная субстанция.
Медиальная «компактная» часть ● , DA-нейроны, аксоны идут в базаль-ные ганглии
Слайд 29Красное ядро.
Передняя (мелкоклеточная) часть: вместе с нижней оливой передает сигналы от коры
Красное ядро.
Передняя (мелкоклеточная) часть: вместе с нижней оливой передает сигналы от коры
Слайд 30Руброспиный тракт – часть так называемой экстрапирамидной системы управления движениями, в которую
Руброспиный тракт – часть так называемой экстрапирамидной системы управления движениями, в которую
Слайд 31Гипоталамус и терморегуляция.
В передней части гипоталамуса (преоптическая область) – нейроны-терморецепторы, постоянно измеряют
Гипоталамус и терморегуляция.
В передней части гипоталамуса (преоптическая область) – нейроны-терморецепторы, постоянно измеряют
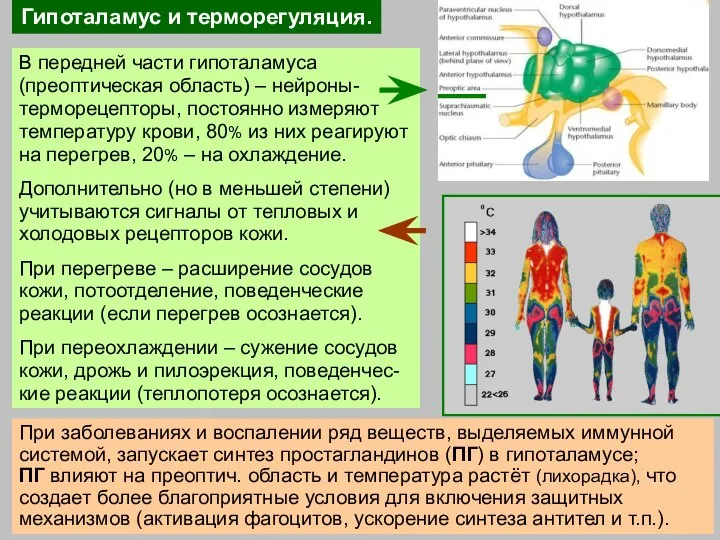
Слайд 32Закаливание – тренировка систем терморегуляции; снижает вероятность простудных заболеваний.
У животных – особые
Закаливание – тренировка систем терморегуляции; снижает вероятность простудных заболеваний.
У животных – особые

 Искусственный отбор
Искусственный отбор Генотип и фенотип. Типы взаимодействия генов. Основы менделевской генетики. Первый и второй закон Менделя
Генотип и фенотип. Типы взаимодействия генов. Основы менделевской генетики. Первый и второй закон Менделя Биологическая изменчивость людей в связи с биогеографическими особенностями среды
Биологическая изменчивость людей в связи с биогеографическими особенностями среды Сахар, соли, белки, аминокислоты
Сахар, соли, белки, аминокислоты Лягушки Евразии
Лягушки Евразии Презентация на тему Селекция микроорганизмов. Биотехнология
Презентация на тему Селекция микроорганизмов. Биотехнология  Волшебный цветок
Волшебный цветок Гуморальная регуляция
Гуморальная регуляция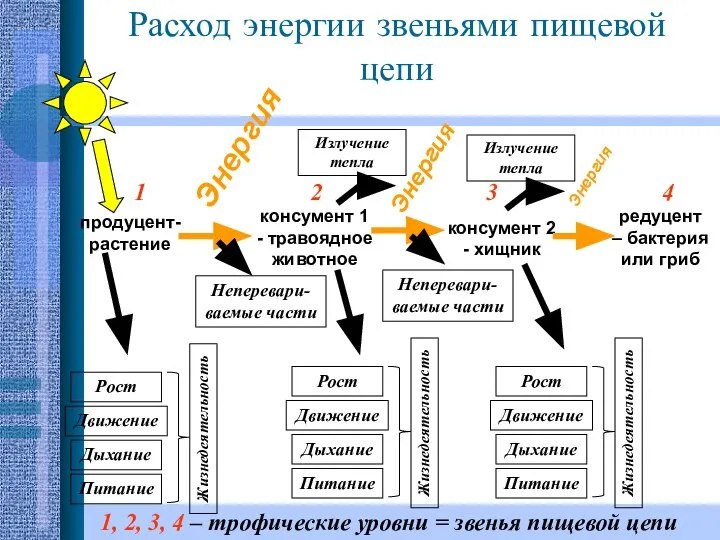 Расход энергии звеньями пищевой цепи
Расход энергии звеньями пищевой цепи Презентация на тему "Проект "Ні шкідливим звичкам!"" - презентации по Биологии
Презентация на тему "Проект "Ні шкідливим звичкам!"" - презентации по Биологии Строение клетки
Строение клетки В подводном царстве, в загадочном государстве. Всемирный день китов и дельфинов
В подводном царстве, в загадочном государстве. Всемирный день китов и дельфинов Жизненные циклы высших растений
Жизненные циклы высших растений Lac-оперон. Структура Lac-оперона
Lac-оперон. Структура Lac-оперона Экология внутренней среды организма человека
Экология внутренней среды организма человека Ксения Петровна Гемп. Начальник водорослей
Ксения Петровна Гемп. Начальник водорослей Внутренняя среда организма. 8 класс
Внутренняя среда организма. 8 класс Характеристика царства бактерий. Биология (5 класс)
Характеристика царства бактерий. Биология (5 класс) Міні-проект “Майстернісь маскування”
Міні-проект “Майстернісь маскування” Презентация на тему Растения сада
Презентация на тему Растения сада 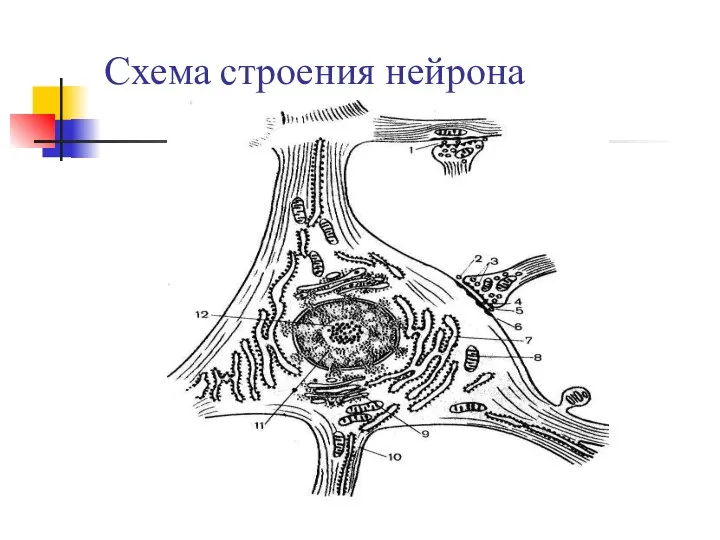 Схема строения нейрона
Схема строения нейрона Классификация и номенклатура биоорганических соединений. Структура и функции биолекул
Классификация и номенклатура биоорганических соединений. Структура и функции биолекул Оплодотворение. Биологическая сущность
Оплодотворение. Биологическая сущность Развитие жизни в кайнозое
Развитие жизни в кайнозое Признаки живых организмов
Признаки живых организмов Виды газонов, классификация газонов и характеристика каждого вида. Виды рулонного газона
Виды газонов, классификация газонов и характеристика каждого вида. Виды рулонного газона Чудо-амфибии
Чудо-амфибии Поджелудочная железа
Поджелудочная железа