- Уровни структурной организации белков

Содержание
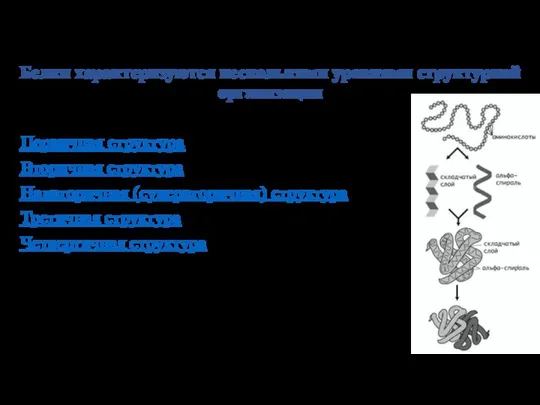
- 2. Белки характеризуются несколькими уровнями структурной организации Первичная структура Вторичная структура Надвторичная (супервторичная) структура Третичная структура Четвертичная
- 3. Первичная структура Первичная структура белка – последовательность аминокислотных остатков в молекуле белка, соединенных пептидными связями. Пептидные
- 4. Пептидная связь очень прочная (так называемая полуторная связь), поэтому полный гидролиз белка до аминокислот в пробирке
- 5. Процесс установления первичной структуры белка получил название «секвенирование». Первоначально секвенирование проводили что называется вручную, осуществляя последовательный
- 6. Вторичная структура Белковые молекулы содержат множество пептидных связей, поляризованные (-С=ОQ- и – NHQ+ и –N-HQ+) группы
- 7. В а-спирали (рис. 1-5) каждая NH – группа полипептидного остова образует водородную связь (-С=ОQ- НQ+ -N-)
- 8. В образовании В-складчатого листа также участвует водородные связи, образованные между –С=О и –N-Н группами (рис. 1-6).
- 9. В тех случаях, когда направление полипептидных цепей одинаково, образуются параллельные В-структуры (см. рис. 1-6а), если же
- 10. Аминокислота пролин, например, после образования пептидной связи не содержит –NH – группы и, следовательно, не может
- 11. Присутствие аминокислотных остатков, не способствующих формированию вторичной структуры, объясняет, что во многих глобулярных белках регулярные элементы
- 12. Для схематического изображения видов вторичной структуры обычно используют следующие символы: а-спираль обычно представляют в виде свернутой
- 13. Надвторичная (супервторичная) структура Следующий за вторичной структурой уровень организации белков получил название надвторичной (супервторичной) структуры. Так
- 14. Надвторичная структура представляет промежуточный этап на пути формирования третичной структуры всей молекулы белка. При этом элементы
- 15. даже если ее вырезать из полипептидной цепи данного белка. Во многих случаях домены выполняют определенные функции.
- 16. Третичная структура белка Под третичной структурой понимают пространственное расположение всех атомов белковой молекулы. Укладку, свойственную каждому
- 18. Третичную структуру стабилизируют ковалентными дисульфидными связями (называют так же дисульфидными мостиками). Они образуются при взаимодействии дисульфидных
- 19. Ионные связи, возникающие в результате электростатического притяжения между противоположно заряженными функциональными группами боковых цепей аминокислотных остатков;
- 20. В зависимости от соотношения регулярных элементов вторичной структуры все типы пространственной структуры разделены на 4 основных
- 21. Четвертичная структура Под четвертичной структурой понимают укладку взаимодействующих между субъединиц в функционально активном белковом комплексе. Субъединицы
- 22. Они и формируют области межсубъединичных контактов, которыми, как правило, и ограничивается взаимодействие субъединиц (рис. 1-11). Межсубъединичные
- 23. Несмотря на самостоятельность субъединиц в четвертичной структуре олигомерного белка, все они представляют «единую команду», выполняющую определенные
- 25. Формирование метаболических и транспортных туннелей. В уже упоминаемой синтезе жирных кислот промежуточные продукты синтеза жирных кислот
- 26. В результате присоединения последующих молекул кислорода к остальным субъединицам происходит намного эффективнее. Этот эффект получил название
- 27. Субъединицы гемоглобина имеют третичную структуру, очень схожую с миоглобином – кислородпереносящим белком мышц, который принимает кислород
- 28. Надмолекулярные комплексы Четвертичная структура имеет ряд преимуществ, которые были рассмотрены в предыдущем разделе. Как оказалось, многие
- 29. Денатурация и ренатурация белка Денатурация – нарушение третичной и вторичной структур белка, приводящее к утрате его
- 30. последними, вызывают нарушение системы водородных связей. В результате этого происходит разупорядочивание структуры белковой молекулы. Органические растворители
- 32. Наиболее демонстративным приемом успешной ренатурации считают рефолдинг in vitro фермента панкреатической рибонуклеазы, денатурированной под действием высоких
- 33. Молекулярные шапероны Шапероны – специализированные внутриклеточные белки, ответственные за быстрое нахождение правильной пространственной структуры. Они могут
- 34. По мере усложнения структурной организации образовываться несколько промежуточных форм сворачиваемого белка, которые в конце концов будут
- 35. Прионы – белки, которые могут находиться в одной из двух различных конформаций. В третичной структуре нормального
- 36. Нуклеиновые кислоты Нуклеиновые кислоты - это водорастворимые высокомолекулярные соединения, макромолекулы которых состоят из многократно повторяющихся звеньев
- 37. Фрагмент макромолекулы нуклеиновой кислоты моносахарид-основание называется нуклеозидом, а фрагмент моносахарид-основание-фосфат - нуклеотидом. Нуклеиновые кислоты присутствуют в
- 38. Компоненты нуклеиновых кислот Азотистые основания. В состав нуклеиновых кислот входят гетероциклические основания пиримидинового ряда: урацил, тимин,
- 39. Углеводы Углеводным компонентом нуклеиновых кислот являются пентозы: D-рибоза или 2-дезокси-D-рибоза. Обе пентозы всегда находятся в фуранозной
- 40. Фосфорная кислота Остатки фосфорной кислоты (H3PO4) входят в состав нуклеотидов и этерифицируют D-рибозу и 2-дезокси-D-рибозу в
- 41. Нуклеозиды Нуклеозиды состоят из гетероциклических оснований, связанных с моносахаридом N-гликозидной связью. В образовании этой связи участвуют
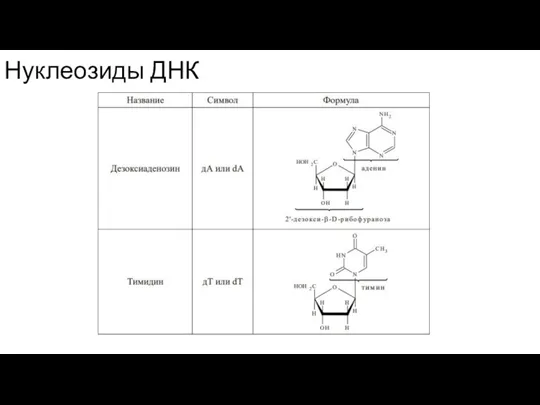
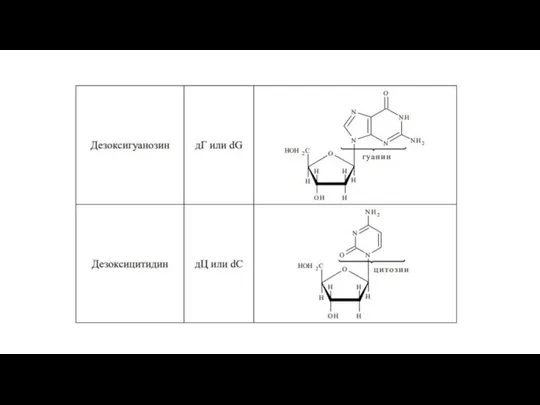
- 42. В зависимости от углеводного остатка нуклеозиды делятся на рибонуклеозиды и дезоксирибонуклеозиды. Номенклатурные названия нуклеозидов строятся аналогично
- 43. Нуклеозиды ДНК
- 45. Нуклеозиды РНК.
- 47. Нуклеозиды значительно лучше растворимы в воде, чем исходные азотистые основания. Подобно всем гликозидам, нуклеозиды устойчивы к
- 48. Нуклеотиды
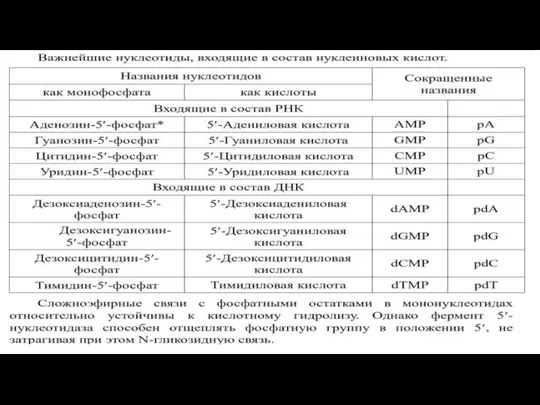
- 49. Мононуклеотиды Мононуклеотиды - это повторяющиеся мономерные единицы олигонуклеотидов и полинуклеотидов. Нуклеозидмонофосфаты. Нуклеозидмонофосфаты содержат только один остаток
- 52. Нуклеозидмоноциклофосфаты. Существуют нуклеотиды, в которых фосфорная кислота одновременно этерифицирует две гидроксильные группы пентозного остатка с образованием
- 53. Нуклеозид-5 ՚ -дифосфаты (НДФ) и нуклеозид-5 ՚ -трифосфаты (НТФ).
- 55. Олиго- и полинуклеотиды Олигонуклеотидами называют соединения, в которых несколько нуклеозидов (до 20) соединены между собой фосфодиэфирными
- 56. Первичная структура нуклеиновых кислот Первичная структура нуклеиновых кислот это нуклеотидный состав и определенная последовательность нуклеотидных
- 57. Первичная структура цепи ДНК и РНК.
- 58. Первичная структура цепи ДНК и РНК
- 59. Вторичная структура нуклеиновых кислот Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы полинуклеотидных цепей. Вторичная
- 60. Вторичная структура ДНК Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями, направленными внутрь спирали.
- 62. Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими ее свойствами: 1) Молекулы
- 63. Вторичная структура РНК В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не
- 64. Вторичная структура РНК Основная роль РНК - непосредственное участие в биосинтезе белка. Известны три вида клеточных
- 65. Химические свойства нуклеиновых кислот ДНК и РНК имеют много общих химических и физических свойств. Так, нуклеиновые
- 66. Ферментативный гидролиз Гидролиз ДНК и РНК также протекает и при участии специфических ферментов - нуклеаз. Эти
- 68. Скачать презентацию
Слайд 2Белки характеризуются несколькими уровнями структурной организации
Первичная структура
Вторичная структура
Надвторичная (супервторичная) структура
Третичная структура
Четвертичная
Белки характеризуются несколькими уровнями структурной организации
Первичная структура
Вторичная структура
Надвторичная (супервторичная) структура
Третичная структура
Четвертичная
Слайд 3Первичная структура
Первичная структура белка – последовательность аминокислотных остатков в молекуле белка,
Первичная структура
Первичная структура белка – последовательность аминокислотных остатков в молекуле белка,

Слайд 4 Пептидная связь очень прочная (так называемая полуторная связь), поэтому полный гидролиз
Пептидная связь очень прочная (так называемая полуторная связь), поэтому полный гидролиз

Слайд 5Процесс установления первичной структуры белка получил название «секвенирование». Первоначально секвенирование проводили что
Процесс установления первичной структуры белка получил название «секвенирование». Первоначально секвенирование проводили что

Слайд 6Вторичная структура
Белковые молекулы содержат множество пептидных связей, поляризованные (-С=ОQ- и
Вторичная структура
Белковые молекулы содержат множество пептидных связей, поляризованные (-С=ОQ- и

Слайд 7В а-спирали (рис. 1-5) каждая NH – группа полипептидного остова образует водородную
В а-спирали (рис. 1-5) каждая NH – группа полипептидного остова образует водородную

Слайд 8 В образовании В-складчатого листа также участвует водородные связи, образованные между –С=О
В образовании В-складчатого листа также участвует водородные связи, образованные между –С=О

Слайд 9 В тех случаях, когда направление полипептидных цепей одинаково, образуются параллельные В-структуры
В тех случаях, когда направление полипептидных цепей одинаково, образуются параллельные В-структуры

Слайд 10 Аминокислота пролин, например, после образования пептидной связи не содержит –NH –
Аминокислота пролин, например, после образования пептидной связи не содержит –NH –

Слайд 11 Присутствие аминокислотных остатков, не способствующих формированию вторичной структуры, объясняет, что во
Присутствие аминокислотных остатков, не способствующих формированию вторичной структуры, объясняет, что во

Слайд 12 Для схематического изображения видов вторичной структуры обычно используют следующие символы:
а-спираль обычно
Для схематического изображения видов вторичной структуры обычно используют следующие символы:
а-спираль обычно

Слайд 13Надвторичная (супервторичная) структура
Следующий за вторичной структурой уровень организации белков получил название
Надвторичная (супервторичная) структура
Следующий за вторичной структурой уровень организации белков получил название

Слайд 14 Надвторичная структура представляет промежуточный этап на пути формирования третичной структуры всей
Надвторичная структура представляет промежуточный этап на пути формирования третичной структуры всей

Слайд 15даже если ее вырезать из полипептидной цепи данного белка. Во многих случаях
даже если ее вырезать из полипептидной цепи данного белка. Во многих случаях

Слайд 16Третичная структура белка
Под третичной структурой понимают пространственное расположение всех атомов белковой
Третичная структура белка
Под третичной структурой понимают пространственное расположение всех атомов белковой

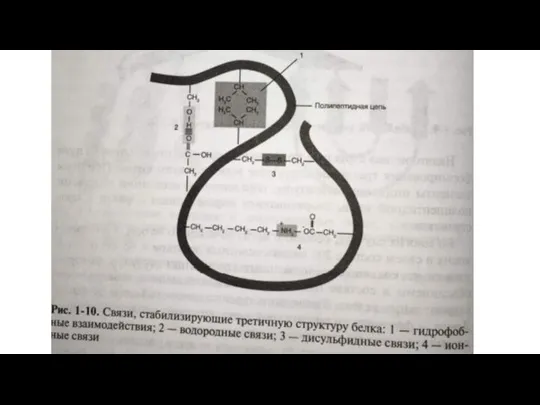
Слайд 18 Третичную структуру стабилизируют ковалентными дисульфидными связями (называют так же дисульфидными мостиками).
Третичную структуру стабилизируют ковалентными дисульфидными связями (называют так же дисульфидными мостиками).

Слайд 19Ионные связи, возникающие в результате электростатического притяжения между противоположно заряженными функциональными группами
Ионные связи, возникающие в результате электростатического притяжения между противоположно заряженными функциональными группами

Слайд 20 В зависимости от соотношения регулярных элементов вторичной структуры все типы пространственной
В зависимости от соотношения регулярных элементов вторичной структуры все типы пространственной

Слайд 21Четвертичная структура
Под четвертичной структурой понимают укладку взаимодействующих между субъединиц в функционально
Четвертичная структура
Под четвертичной структурой понимают укладку взаимодействующих между субъединиц в функционально

Слайд 22Они и формируют области межсубъединичных контактов, которыми, как правило, и ограничивается взаимодействие
Они и формируют области межсубъединичных контактов, которыми, как правило, и ограничивается взаимодействие

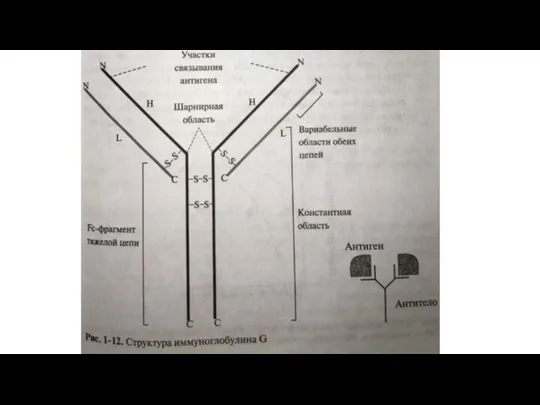
Слайд 23 Несмотря на самостоятельность субъединиц в четвертичной структуре олигомерного белка, все они
Несмотря на самостоятельность субъединиц в четвертичной структуре олигомерного белка, все они

Слайд 25Формирование метаболических и транспортных туннелей. В уже упоминаемой синтезе жирных кислот промежуточные
Формирование метаболических и транспортных туннелей. В уже упоминаемой синтезе жирных кислот промежуточные

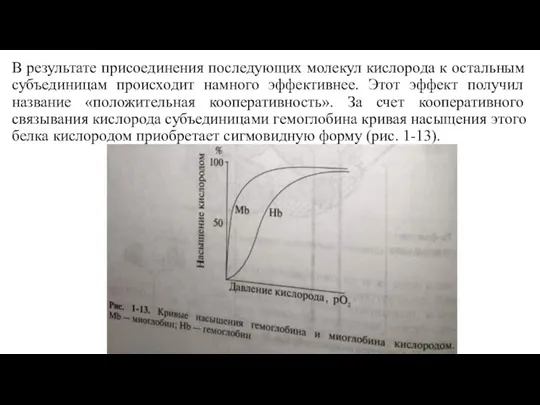
Слайд 26В результате присоединения последующих молекул кислорода к остальным субъединицам происходит намного эффективнее.
В результате присоединения последующих молекул кислорода к остальным субъединицам происходит намного эффективнее.
Слайд 27 Субъединицы гемоглобина имеют третичную структуру, очень схожую с миоглобином – кислородпереносящим
Субъединицы гемоглобина имеют третичную структуру, очень схожую с миоглобином – кислородпереносящим

Слайд 28Надмолекулярные комплексы
Четвертичная структура имеет ряд преимуществ, которые были рассмотрены в предыдущем
Надмолекулярные комплексы
Четвертичная структура имеет ряд преимуществ, которые были рассмотрены в предыдущем

Слайд 29Денатурация и ренатурация белка
Денатурация – нарушение третичной и вторичной структур белка,
Денатурация и ренатурация белка
Денатурация – нарушение третичной и вторичной структур белка,

Слайд 30последними, вызывают нарушение системы водородных связей. В результате этого происходит разупорядочивание структуры
последними, вызывают нарушение системы водородных связей. В результате этого происходит разупорядочивание структуры

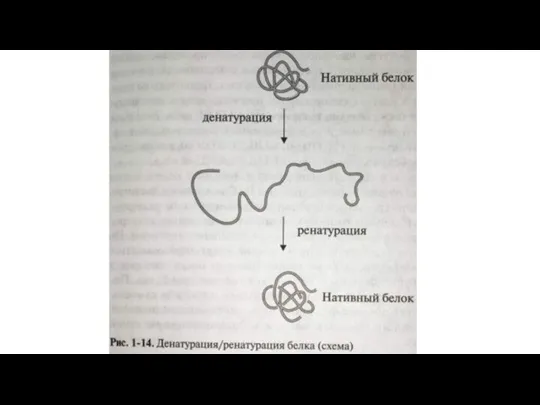
Слайд 32 Наиболее демонстративным приемом успешной ренатурации считают рефолдинг in vitro фермента панкреатической
Наиболее демонстративным приемом успешной ренатурации считают рефолдинг in vitro фермента панкреатической

Слайд 33Молекулярные шапероны
Шапероны – специализированные внутриклеточные белки, ответственные за быстрое нахождение правильной
Молекулярные шапероны
Шапероны – специализированные внутриклеточные белки, ответственные за быстрое нахождение правильной

Слайд 34По мере усложнения структурной организации образовываться несколько промежуточных форм сворачиваемого белка, которые
По мере усложнения структурной организации образовываться несколько промежуточных форм сворачиваемого белка, которые

Слайд 35 Прионы – белки, которые могут находиться в одной из двух различных
Прионы – белки, которые могут находиться в одной из двух различных

Слайд 36Нуклеиновые кислоты
Нуклеиновые кислоты - это водорастворимые высокомолекулярные соединения, макромолекулы которых состоят из
Нуклеиновые кислоты
Нуклеиновые кислоты - это водорастворимые высокомолекулярные соединения, макромолекулы которых состоят из

Слайд 37 Фрагмент макромолекулы нуклеиновой кислоты моносахарид-основание называется нуклеозидом, а фрагмент моносахарид-основание-фосфат - нуклеотидом.
Фрагмент макромолекулы нуклеиновой кислоты моносахарид-основание называется нуклеозидом, а фрагмент моносахарид-основание-фосфат - нуклеотидом.

Слайд 38Компоненты нуклеиновых кислот
Азотистые основания.
В состав нуклеиновых кислот входят гетероциклические основания пиримидинового
Компоненты нуклеиновых кислот
Азотистые основания.
В состав нуклеиновых кислот входят гетероциклические основания пиримидинового

Слайд 39Углеводы
Углеводным компонентом нуклеиновых кислот являются пентозы: D-рибоза или 2-дезокси-D-рибоза. Обе пентозы всегда
Углеводы
Углеводным компонентом нуклеиновых кислот являются пентозы: D-рибоза или 2-дезокси-D-рибоза. Обе пентозы всегда

Слайд 40Фосфорная кислота
Остатки фосфорной кислоты (H3PO4) входят в состав нуклеотидов и этерифицируют D-рибозу
Фосфорная кислота
Остатки фосфорной кислоты (H3PO4) входят в состав нуклеотидов и этерифицируют D-рибозу

Слайд 41 Нуклеозиды
Нуклеозиды состоят из гетероциклических оснований, связанных с моносахаридом N-гликозидной связью. В
Нуклеозиды
Нуклеозиды состоят из гетероциклических оснований, связанных с моносахаридом N-гликозидной связью. В

Слайд 42 В зависимости от углеводного остатка нуклеозиды делятся на рибонуклеозиды и дезоксирибонуклеозиды. Номенклатурные
В зависимости от углеводного остатка нуклеозиды делятся на рибонуклеозиды и дезоксирибонуклеозиды. Номенклатурные

Слайд 43Нуклеозиды ДНК
Нуклеозиды ДНК
Слайд 45Нуклеозиды РНК.
Нуклеозиды РНК.


Слайд 47 Нуклеозиды значительно лучше растворимы в воде, чем исходные азотистые основания. Подобно всем
Нуклеозиды значительно лучше растворимы в воде, чем исходные азотистые основания. Подобно всем

Слайд 48Нуклеотиды
Нуклеотиды

Слайд 49Мононуклеотиды
Мононуклеотиды - это повторяющиеся мономерные единицы
олигонуклеотидов и полинуклеотидов.
Нуклеозидмонофосфаты.
Нуклеозидмонофосфаты содержат
Мононуклеотиды
Мононуклеотиды - это повторяющиеся мономерные единицы
олигонуклеотидов и полинуклеотидов.
Нуклеозидмонофосфаты.
Нуклеозидмонофосфаты содержат


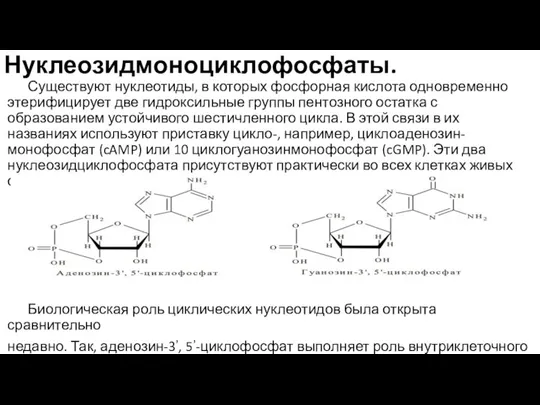
Слайд 52Нуклеозидмоноциклофосфаты.
Существуют нуклеотиды, в которых фосфорная кислота одновременно этерифицирует две гидроксильные группы пентозного
Нуклеозидмоноциклофосфаты.
Существуют нуклеотиды, в которых фосфорная кислота одновременно этерифицирует две гидроксильные группы пентозного
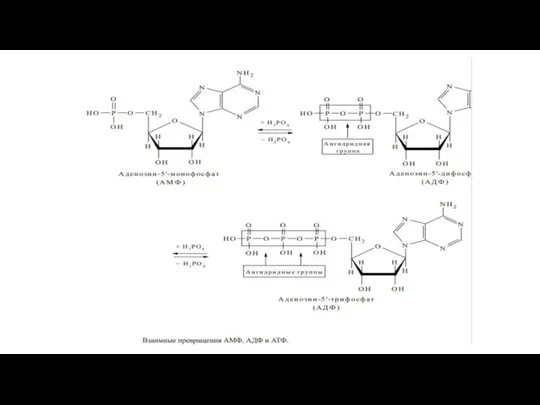
Слайд 53Нуклеозид-5 ՚ -дифосфаты (НДФ) и нуклеозид-5 ՚ -трифосфаты (НТФ).
Нуклеозид-5 ՚ -дифосфаты (НДФ) и нуклеозид-5 ՚ -трифосфаты (НТФ).

Слайд 55Олиго- и полинуклеотиды
Олигонуклеотидами называют соединения, в которых несколько нуклеозидов (до 20)
Олиго- и полинуклеотиды
Олигонуклеотидами называют соединения, в которых несколько нуклеозидов (до 20)

Слайд 56Первичная структура нуклеиновых кислот
Первичная структура нуклеиновых кислот это нуклеотидный состав и
Первичная структура нуклеиновых кислот
Первичная структура нуклеиновых кислот это нуклеотидный состав и

Слайд 57Первичная структура цепи ДНК и РНК.
Первичная структура цепи ДНК и РНК.

Слайд 58Первичная структура цепи ДНК и РНК
Первичная структура цепи ДНК и РНК

Слайд 59Вторичная структура нуклеиновых кислот
Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы
Вторичная структура нуклеиновых кислот
Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы

Слайд 60Вторичная структура ДНК
Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями,
Вторичная структура ДНК
Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями,


Слайд 62 Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими
Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими

Слайд 63Вторичная структура РНК
В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной
Вторичная структура РНК
В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной

Слайд 64Вторичная структура РНК
Основная роль РНК - непосредственное участие в биосинтезе белка.
Известны
Вторичная структура РНК
Основная роль РНК - непосредственное участие в биосинтезе белка.
Известны

Слайд 65Химические свойства нуклеиновых кислот
ДНК и РНК имеют много общих химических и физических
Химические свойства нуклеиновых кислот
ДНК и РНК имеют много общих химических и физических

Слайд 66Ферментативный гидролиз
Гидролиз ДНК и РНК также протекает и при участии специфических ферментов
Ферментативный гидролиз
Гидролиз ДНК и РНК также протекает и при участии специфических ферментов

 Презентация по биологии на тему _Водоросли_ (7 класс)
Презентация по биологии на тему _Водоросли_ (7 класс) Железы внутренней секреции и их функции. 8 класс
Железы внутренней секреции и их функции. 8 класс Презентация на тему СЕКРЕТЫ ДОЛГОЛЕТИЯ
Презентация на тему СЕКРЕТЫ ДОЛГОЛЕТИЯ  Интересные факты о человеческом теле
Интересные факты о человеческом теле Луговое разнотравье. Экологическая викторина
Луговое разнотравье. Экологическая викторина Функции кожи
Функции кожи Наследственная изменчивость
Наследственная изменчивость Презентация на тему Гельминты человека
Презентация на тему Гельминты человека  Ребусы о растениях
Ребусы о растениях Көпфакторлы дисперсиялық талдау. Көпфакторлы дисперсиялық талдаудың бірфакторлы талдаудан негізгі айырмашылығы
Көпфакторлы дисперсиялық талдау. Көпфакторлы дисперсиялық талдаудың бірфакторлы талдаудан негізгі айырмашылығы Состав и функции белков
Состав и функции белков Вид, критерии вида
Вид, критерии вида Різноманітність птахів
Різноманітність птахів Растения России
Растения России Голосеменные хвойные растения
Голосеменные хвойные растения Презентация на тему ТЕЛА И ВЕЩЕСТВА
Презентация на тему ТЕЛА И ВЕЩЕСТВА  Патология системы внешнего дыхания
Патология системы внешнего дыхания Фотосинтез и его значение в природе и жизни человека
Фотосинтез и его значение в природе и жизни человека Прирождённые разрушители
Прирождённые разрушители Законы Менделя
Законы Менделя Тип Плоские черви
Тип Плоские черви Строение и работа скелетных мышц
Строение и работа скелетных мышц Генетические доказательства эволюции
Генетические доказательства эволюции Лютиковые. Живокость высокая
Лютиковые. Живокость высокая Побег. Строение и значение побега
Побег. Строение и значение побега Основные группы микроорганизмов
Основные группы микроорганизмов Опорно-двигательная, нервная, система размножения и органы выделения животных. 6 класс
Опорно-двигательная, нервная, система размножения и органы выделения животных. 6 класс Птичий двор. Самые быстрые птицы мира
Птичий двор. Самые быстрые птицы мира