- Взаимодействие генов

Содержание
- 2. Взаимодействие генов Ген – последовательность нуклеотидов (участок ДНК), которой может быть приписана определенная функция в организме.
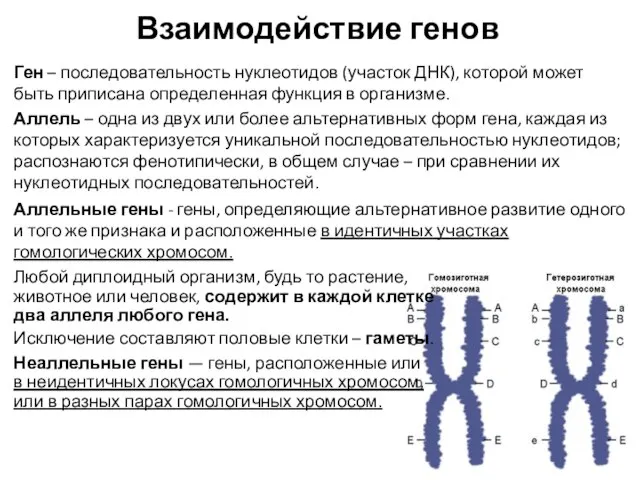
- 3. Взаимодействие аллельных генов Полное доминирование — это вид взаимодействия аллельных генов, при котором фенотип гетерозигот не
- 4. Взаимодействие неаллельных генов Комплементарность — вид взаимодействия неаллельных генов, при котором признак формируется в результате суммарного
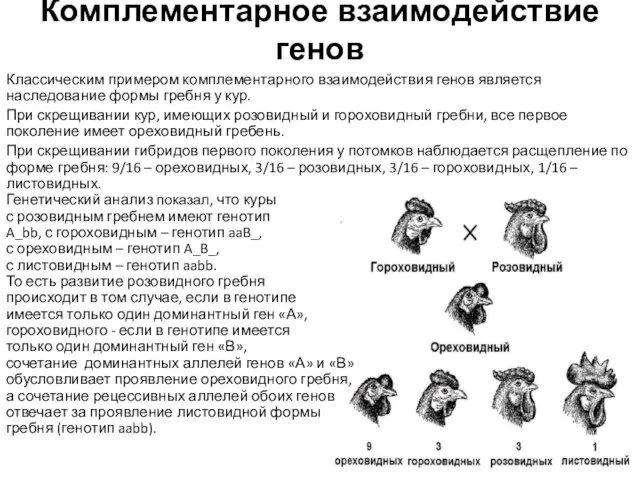
- 5. Комплементарное взаимодействие генов Классическим примером комплементарного взаимодействия генов является наследование формы гребня у кур. При скрещивании
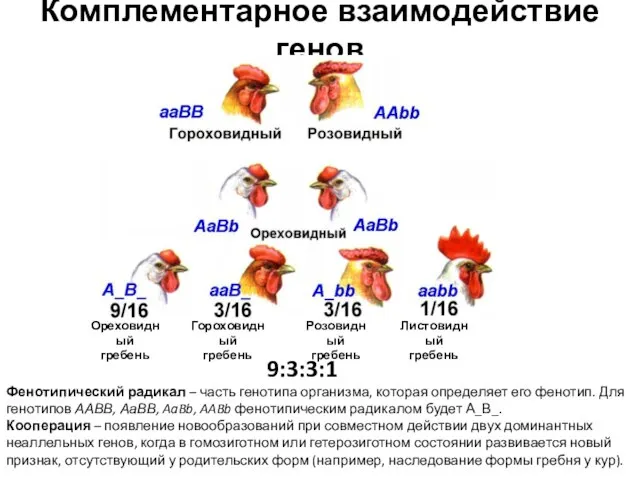
- 6. Комплементарное взаимодействие генов Ореховидный гребень Гороховидный гребень Розовидный гребень Листовидный гребень Фенотипический радикал – часть генотипа
- 7. Комплементарное взаимодействие генов
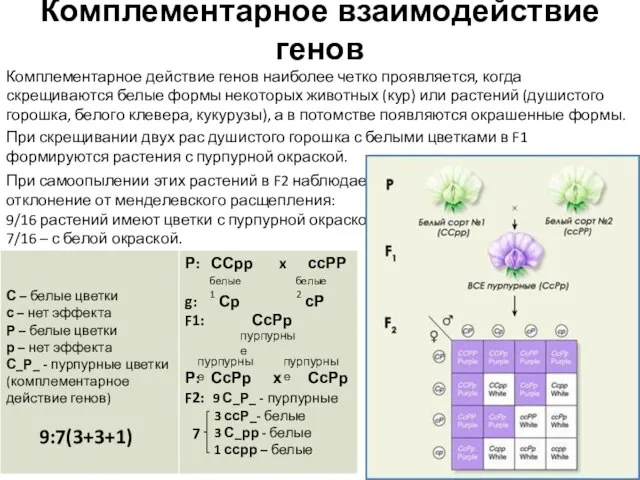
- 8. Комплементарное взаимодействие генов Комплементарное действие генов наиболее четко проявляется, когда скрещиваются белые формы некоторых животных (кур)
- 9. Комплементарное взаимодействие генов темно-синие темно-синие
- 10. Комплементарное взаимодействие генов При скрещивании черной и белой мышей в F1 все потомство будет иметь серую
- 11. Комплементарное взаимодействие генов 9:6(3+3):1
- 12. Комплементарное взаимодействие генов 9:3:3:1 – по отдельности доминантные аллели генов отвечают за «свои признаки», присутствие обоих
- 13. Криптомерия (взаимодействие по типу проявления) Некоторые гены не проявляют своего действия фенотипически до тех пор, пока
- 14. Криптомерия (взаимодействие по типу проявления) Пример: У льна доминантный аллель отвечает за голубую окраску цветков, рецессивный
- 15. Эпистаз (взаимодействие по типу подавления) Эпистаз – тип неаллельного взаимодействия, при котором один ген подавляет фенотипическое
- 16. Доминантный эпистаз (взаимодействие по типу подавления) Примером доминантного эпистаза с расщеп-лением в F2 12:3:1 является взаимодействие
- 17. Доминантный эпистаз (взаимодействие по типу подавления) Пример: доминантный ген C (вызывает раннее поседение, вследствие чего волос
- 18. Доминантный эпистаз (взаимодействие по типу подавления) Аналогичным образом наследуется окраска плодов тыквы. При скрещивании растения с
- 19. Доминантный эпистаз (взаимодействие по типу подавления) … или окраски свиней. При скрещивании черных и белых свиней
- 20. Доминантный эпистаз (взаимодействие по типу подавления) Расщепление 13 : 3 при доминантном эпистазе наблюдается, когда доминантный
- 21. Рецессивный эпистаз (взаимодействие по типу подавления) В строгом смысле эпистаз всегда доминантный, но также выделяют и
- 22. Рецессивный эпистаз (взаимодействие по типу подавления) При скрещивании черной и белой мышей в F1 все потомство
- 23. Двойной рецессивный эпистаз Примером одновременного комплементарного взаимодействия генов и двойного рецессивного эпистаза является наследование окраски цветков
- 24. Наследование альбизизма Альбинизмом (от латинского albus — белый) называют врожденное отсутствие пигмента меланина, придающего окраску коже,
- 25. (Бомбейский феномен)
- 26. Полимерия Полимерия – формирование признака несколькими парами неаллельных доминантных генов. Чем больше таких генов в генотипе,
- 27. Некумулятивная полимерия 15:1
- 28. Кумулятивная полимерия При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем
- 29. Кумулятивная полимерия
- 30. Кумулятивная полимерия 1 (4А) 4 (3А) 6 (2А) 4 (1А) 1 (0А)
- 31. Кумулятивная полимерия Число возможных комбинаций гамет, образующих зиготы в F2: 4 n Минимальная необходимая совокупность особей
- 33. Скачать презентацию
Слайд 2Взаимодействие генов
Ген – последовательность нуклеотидов (участок ДНК), которой может быть приписана определенная
Взаимодействие генов
Ген – последовательность нуклеотидов (участок ДНК), которой может быть приписана определенная
Слайд 3Взаимодействие аллельных генов
Полное доминирование — это вид взаимодействия аллельных генов, при котором
Взаимодействие аллельных генов
Полное доминирование — это вид взаимодействия аллельных генов, при котором

Слайд 4Взаимодействие неаллельных генов
Комплементарность — вид взаимодействия неаллельных генов, при котором признак формируется
Взаимодействие неаллельных генов
Комплементарность — вид взаимодействия неаллельных генов, при котором признак формируется

Слайд 5Комплементарное взаимодействие генов
Классическим примером комплементарного взаимодействия генов является наследование формы гребня у
Комплементарное взаимодействие генов
Классическим примером комплементарного взаимодействия генов является наследование формы гребня у
Слайд 6Комплементарное взаимодействие генов
Ореховидный
гребень
Гороховидный
гребень
Розовидный
гребень
Листовидный
гребень
Фенотипический радикал – часть генотипа организма, которая определяет его
Комплементарное взаимодействие генов
Ореховидный
гребень
Гороховидный
гребень
Розовидный
гребень
Листовидный
гребень
Фенотипический радикал – часть генотипа организма, которая определяет его
Слайд 7Комплементарное взаимодействие генов
Комплементарное взаимодействие генов

Слайд 8Комплементарное взаимодействие генов
Комплементарное действие генов наиболее четко проявляется, когда скрещиваются белые формы
Комплементарное взаимодействие генов
Комплементарное действие генов наиболее четко проявляется, когда скрещиваются белые формы
Слайд 9Комплементарное взаимодействие генов
темно-синие
темно-синие
Комплементарное взаимодействие генов
темно-синие
темно-синие

Слайд 10Комплементарное взаимодействие генов
При скрещивании черной и белой мышей в F1 все потомство
Комплементарное взаимодействие генов
При скрещивании черной и белой мышей в F1 все потомство
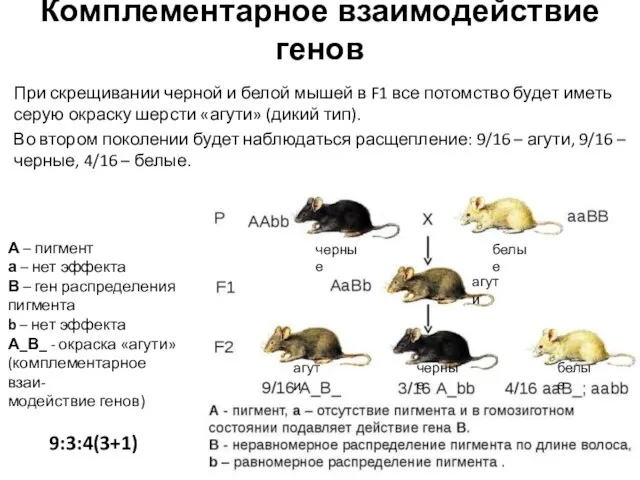
Слайд 11Комплементарное взаимодействие генов
9:6(3+3):1
Комплементарное взаимодействие генов
9:6(3+3):1
Слайд 12Комплементарное взаимодействие генов
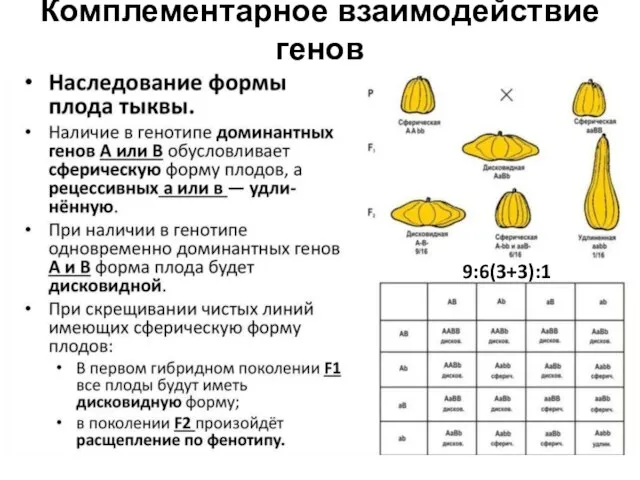
9:3:3:1 – по отдельности доминантные аллели генов отвечают за «свои
Комплементарное взаимодействие генов
9:3:3:1 – по отдельности доминантные аллели генов отвечают за «свои

Слайд 13Криптомерия
(взаимодействие по типу проявления)
Некоторые гены не проявляют своего действия фенотипически до тех
Криптомерия
(взаимодействие по типу проявления)
Некоторые гены не проявляют своего действия фенотипически до тех

Слайд 14Криптомерия
(взаимодействие по типу проявления)
Пример: У льна доминантный аллель отвечает за голубую окраску
Криптомерия
(взаимодействие по типу проявления)
Пример: У льна доминантный аллель отвечает за голубую окраску

Слайд 15Эпистаз
(взаимодействие по типу подавления)
Эпистаз – тип неаллельного взаимодействия, при котором один ген подавляет
Эпистаз
(взаимодействие по типу подавления)
Эпистаз – тип неаллельного взаимодействия, при котором один ген подавляет

Слайд 16Доминантный эпистаз
(взаимодействие по типу подавления)
Примером доминантного эпистаза с расщеп-лением в F2 12:3:1
Доминантный эпистаз
(взаимодействие по типу подавления)
Примером доминантного эпистаза с расщеп-лением в F2 12:3:1

Слайд 17Доминантный эпистаз
(взаимодействие по типу подавления)
Пример: доминантный ген C (вызывает раннее поседение, вследствие
Доминантный эпистаз
(взаимодействие по типу подавления)
Пример: доминантный ген C (вызывает раннее поседение, вследствие

Слайд 18Доминантный эпистаз
(взаимодействие по типу подавления)
Аналогичным образом наследуется окраска плодов тыквы. При скрещивании
Доминантный эпистаз
(взаимодействие по типу подавления)
Аналогичным образом наследуется окраска плодов тыквы. При скрещивании
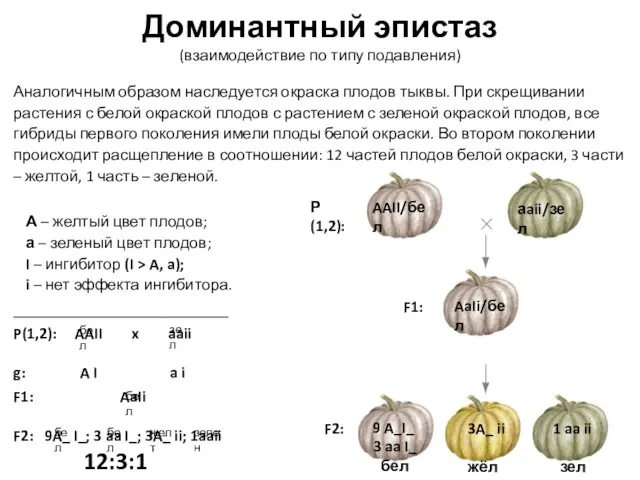
Слайд 19Доминантный эпистаз
(взаимодействие по типу подавления)
… или окраски свиней.
При скрещивании черных и белых
Доминантный эпистаз
(взаимодействие по типу подавления)
… или окраски свиней.
При скрещивании черных и белых

Слайд 20Доминантный эпистаз
(взаимодействие по типу подавления)
Расщепление 13 : 3 при доминантном эпистазе наблюдается,
Доминантный эпистаз
(взаимодействие по типу подавления)
Расщепление 13 : 3 при доминантном эпистазе наблюдается,

Слайд 21Рецессивный эпистаз
(взаимодействие по типу подавления)
В строгом смысле эпистаз всегда доминантный, но также
Рецессивный эпистаз
(взаимодействие по типу подавления)
В строгом смысле эпистаз всегда доминантный, но также

Слайд 22Рецессивный эпистаз
(взаимодействие по типу подавления)
При скрещивании черной и белой мышей в F1
Рецессивный эпистаз
(взаимодействие по типу подавления)
При скрещивании черной и белой мышей в F1

Слайд 23Двойной рецессивный эпистаз
Примером одновременного комплементарного взаимодействия генов и двойного рецессивного эпистаза является
Двойной рецессивный эпистаз
Примером одновременного комплементарного взаимодействия генов и двойного рецессивного эпистаза является
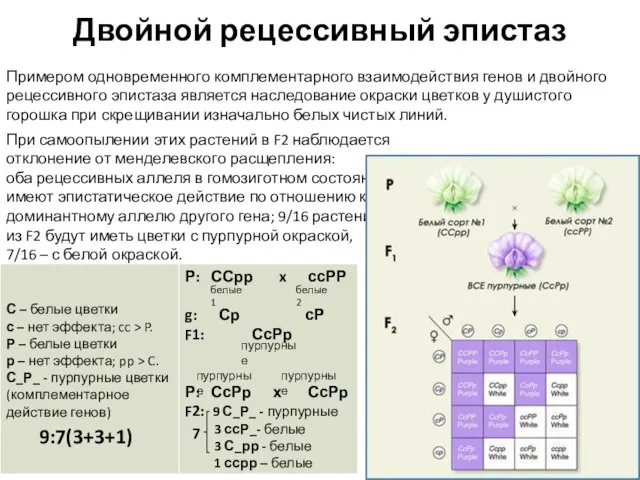
Слайд 24Наследование альбизизма
Альбинизмом (от латинского albus — белый) называют врожденное отсутствие пигмента меланина,
Наследование альбизизма
Альбинизмом (от латинского albus — белый) называют врожденное отсутствие пигмента меланина,

Слайд 25(Бомбейский феномен)
(Бомбейский феномен)

Слайд 26Полимерия
Полимерия – формирование признака несколькими парами неаллельных доминантных генов. Чем больше таких
Полимерия
Полимерия – формирование признака несколькими парами неаллельных доминантных генов. Чем больше таких

Слайд 27Некумулятивная полимерия
15:1
Некумулятивная полимерия
15:1

Слайд 28Кумулятивная полимерия
При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия
Кумулятивная полимерия
При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия

Слайд 29Кумулятивная полимерия
Кумулятивная полимерия

Слайд 30Кумулятивная полимерия
1 (4А)
4 (3А)
6 (2А)
4 (1А)
1 (0А)
Кумулятивная полимерия
1 (4А)
4 (3А)
6 (2А)
4 (1А)
1 (0А)

Слайд 31Кумулятивная полимерия
Число возможных комбинаций гамет, образующих зиготы в F2: 4 n
Минимальная необходимая
Кумулятивная полимерия
Число возможных комбинаций гамет, образующих зиготы в F2: 4 n
Минимальная необходимая

 Значение давления для живых организмов. Измерение артериального и атмосферного давления
Значение давления для живых организмов. Измерение артериального и атмосферного давления Закономірності мінливості
Закономірності мінливості Птицы. Специализация
Птицы. Специализация Класс Насекомые. 7 Класс
Класс Насекомые. 7 Класс Строение клетки клеточная оболочка, плазматическая мембрана, цитоплазма, ядро
Строение клетки клеточная оболочка, плазматическая мембрана, цитоплазма, ядро Общие принципы физиологии пищеварения
Общие принципы физиологии пищеварения Влияние спелого яблока на зелёные помидоры
Влияние спелого яблока на зелёные помидоры Zheludochno-kishechny_trakt_Prezentatsia_1
Zheludochno-kishechny_trakt_Prezentatsia_1 Коалы
Коалы Игра В мире животных
Игра В мире животных Ферменты
Ферменты Царство Грибы
Царство Грибы Микробы
Микробы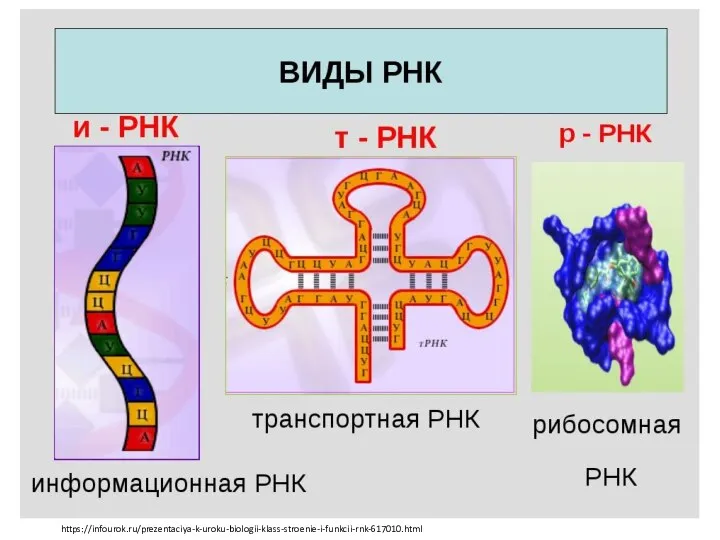 ДНК и РНК
ДНК и РНК Tkáně, základ orgánů
Tkáně, základ orgánů Гаметогенез. Оогенез. Сперматогенез
Гаметогенез. Оогенез. Сперматогенез Дыхательная система. Спланхнология
Дыхательная система. Спланхнология Жизнь в воде
Жизнь в воде Презентация на тему ГЕНЕТИКА ПОЛА
Презентация на тему ГЕНЕТИКА ПОЛА  Царство Грибы
Царство Грибы Кровеносная система
Кровеносная система Растения-обереги. Комнатные растения – обереги
Растения-обереги. Комнатные растения – обереги Координация и регуляция. Рефлекторная природа поведения. Условные и безусловные рефлексы
Координация и регуляция. Рефлекторная природа поведения. Условные и безусловные рефлексы Биомолекулы: ДНК
Биомолекулы: ДНК Загадки Лесовичка
Загадки Лесовичка Презентация на тему Что такое систематика
Презентация на тему Что такое систематика  Учение В.И. Вернадского о биосфере. Эволюция биосферы
Учение В.И. Вернадского о биосфере. Эволюция биосферы Венечный круг кровообращения
Венечный круг кровообращения