- Переваривание и всасывание липидов. Транспортные формы липидов в крови. Лекция № 7

Содержание
- 2. План лекции: Особенности строения липидов. Функции липидов. Классификация липидов. Строение ТАГ. Переваривание и всасывание пищевых ТАГ.
- 3. Липиды Липиды – это разнообразная по строению группа органических молекул, имеющих общие свойства – гидрофобностьили амфифильность.
- 5. Функции липидов Участвуют в формировании мембран: глицерофосфолипиды, сфинголипиды, холестерол; Являются предшественниками коферментов: жирорастворимый витамин К; Образуют
- 6. Строение и функции основных классов липидов человека
- 7. Строение и функции основных классов липидов человека
- 8. Строение и функции основных классов липидов человека
- 9. Жирные кислоты
- 11. ТАГ (жиры) являются сложными эфирами жирных кислот и трехатомного спирта глицерола. К 3 гидроксильным группам глицерола
- 12. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА Основные липиды, поступающие с пищей,
- 13. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА Эмульгирование жиров (смешивание жиров с
- 14. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА Действие панкреатической липазы. Белок -
- 15. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА Образование смешанных мицелл. Продукты гидролиза
- 16. ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА Нарушения переваривания жиров. Снижение секреции,
- 18. Переваривание и всасывание пищевых ТАГ Пищевые ТАГ Большие липидные капли Желчные кислоты Эмульгирование Тонкодисперсная эмульсия COR1
- 19. Гидролиз триацилглицеролов
- 20. ХИЛОМИКРОНЫ - ТРАНСПОРТНАЯ ФОРМА ЭКЗОГЕННЫХ ЖИРОВ Липиды, в частности жиры, холестерол и его эфиры не растворяются
- 21. Общая структура липопротеинов плазмы крови (ХМ, ЛПОНП, ЛППП, ЛПНП, ЛПВП) В организме человека липиды представлены большой
- 23. Липопротеины – транспортные формы липидов
- 24. Путь экзогенных жиров и хиломикронов Рецепторы ХМ ост. ХМ незр. ЛПВП апоС-II апоЕ Кровь ЖК +
- 25. β – окисление жирных кислот – специфический путь катаболизма
- 26. Характеристика β–окисления жирных кислот β–окисление жирных кислот происходит только в аэробных условиях. β–окисление жирных кислот –
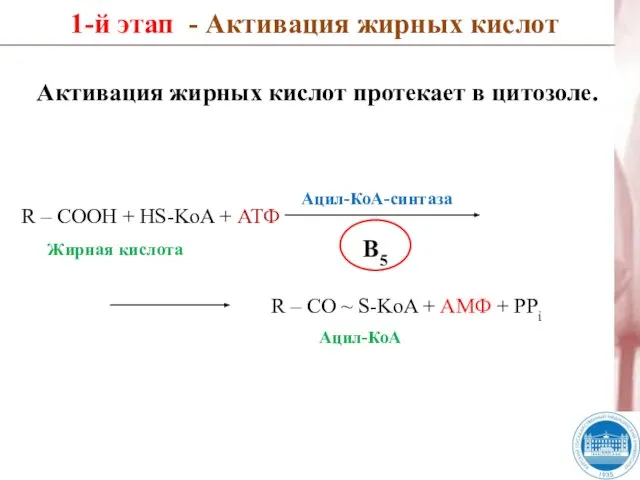
- 27. R – COOH + HS-KoA + АТФ 1-й этап - Активация жирных кислот R – CO
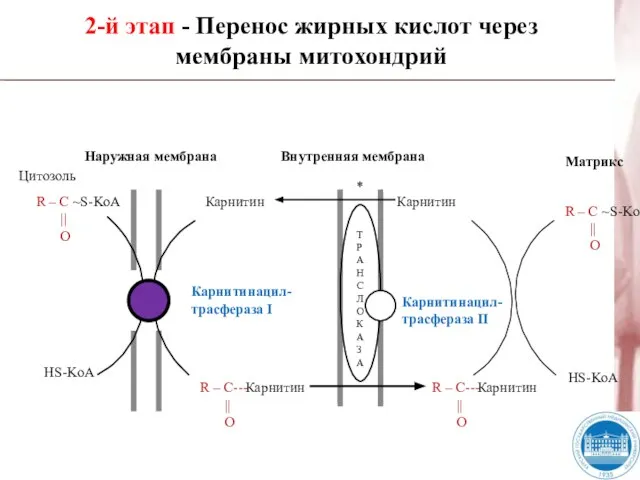
- 28. Наружная мембрана Внутренняя мембрана Цитозоль R – C ~S-KoA || O HS-KoA R – C--- ||
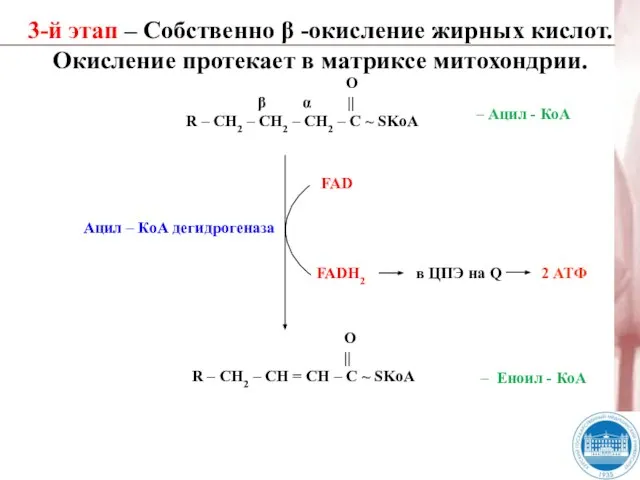
- 29. 3-й этап – Собственно β -окисление жирных кислот. Окисление протекает в матриксе митохондрии. О β α
- 30. 3-й этап – Собственно β -окисление жирных кислот Н2О ОН О | || R – CH2
- 31. 3-й этап – Собственно β -окисление жирных кислот β – Гидроксиацил – КоА дегидрогеназа О О
- 32. 3-й этап – Собственно β -окисление жирных кислот β –Кетоацил– КоА тиолаза HSКоА Следующий цикл β
- 33. Общая формула расчета баланса АТФ при β – окислении жирных кислот с четным числом углеродных атомов
- 34. Обмен жирных кислот с нечетным числом атомов углерода Метил-малонил- КоА-мутаза В12 АТФ АМФ
- 35. Общая формула расчета баланса АТФ при β – окислении жирных кислот с нечетным числом углеродных атомов
- 36. Биосинтез жирных кислот
- 37. Отличия биосинтеза жирных кислот от их окисления Процесс протекает в цитоплазме клетки. Идет с потреблением энергии
- 38. Происхождение субстратов для синтеза жирных кислот и ТАГ Глюкоза Глюкозо – 6 - фосфат Фруктозо– 6
- 39. NADH + H+ NAD+ 1-й этап Перенос ацетильных остатков из митохондрий в цитозоль Цитозоль Митохондрия Глюкоза
- 40. Жирные кислоты синтезируются из ацетил–КоА, который образуется при аэробном окислении глюкозы. Роль переносчика ацетильных групп из
- 41. В цитоплазме ацетил–КоА карбоксилируется и превращается в малонил–КоА – второй субстрат, необходимый для образования жирной кислоты.
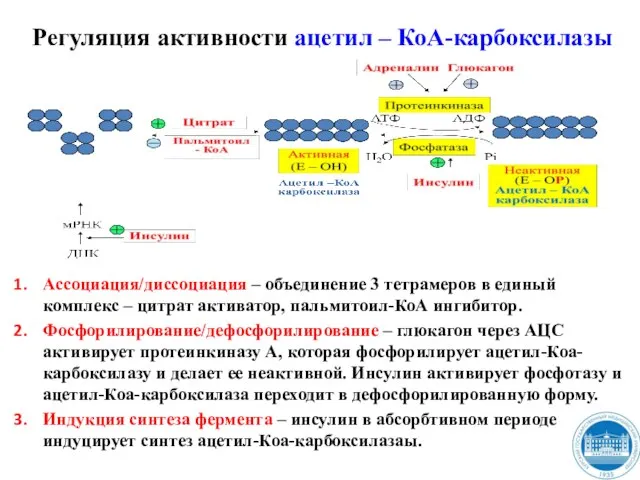
- 42. Регуляция активности ацетил – КоА-карбоксилазы Ассоциация/диссоциация – объединение 3 тетрамеров в единый комплекс – цитрат активатор,
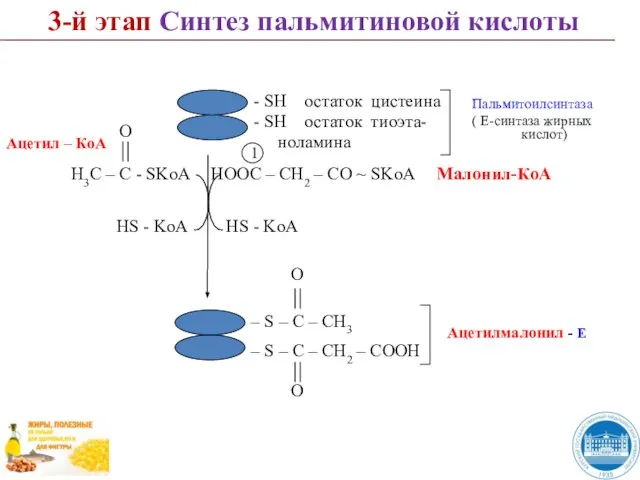
- 43. 3-й этап Синтез пальмитиновой кислоты 1 HS - KoA HOOC – CH2 – CО ~ SKoA
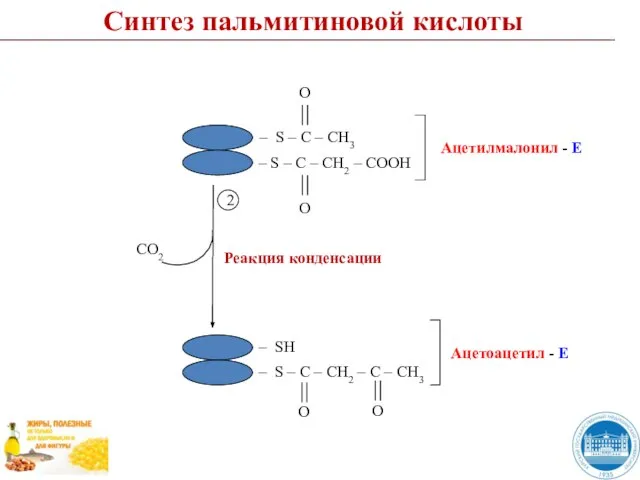
- 44. Синтез пальмитиновой кислоты СО2 Реакция конденсации 2
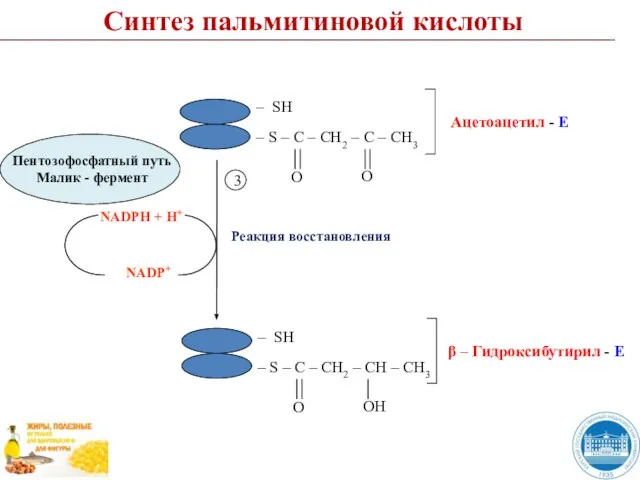
- 45. Синтез пальмитиновой кислоты Реакция восстановления NADPH + H+ NADP+ 3
- 46. Синтез пальмитиновой кислоты H2O Реакция дегидратации 4
- 47. Синтез пальмитиновой кислоты Реакция восстановления 5 NADP+ NADPH + H+
- 48. Синтез пальмитиновой кислоты I цикл 6 НSKoA
- 49. 7 CO2 Синтез пальмитиновой кислоты
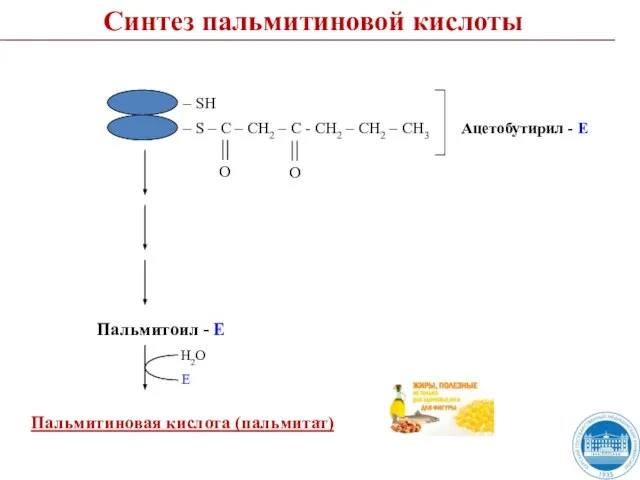
- 50. Пальмитиновая кислота (пальмитат) Пальмитоил - Е Н2О Е Синтез пальмитиновой кислоты
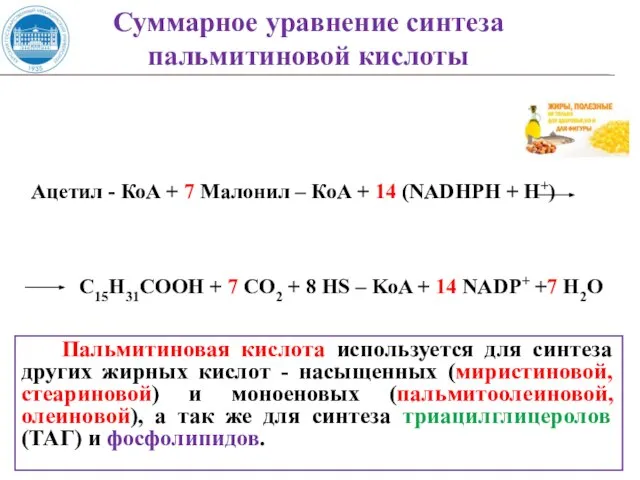
- 51. Ацетил - КоА + 7 Малонил – КоА + 14 (NADHPH + H+) C15H31COOH + 7
- 52. Биосинтез триацилглицеролов
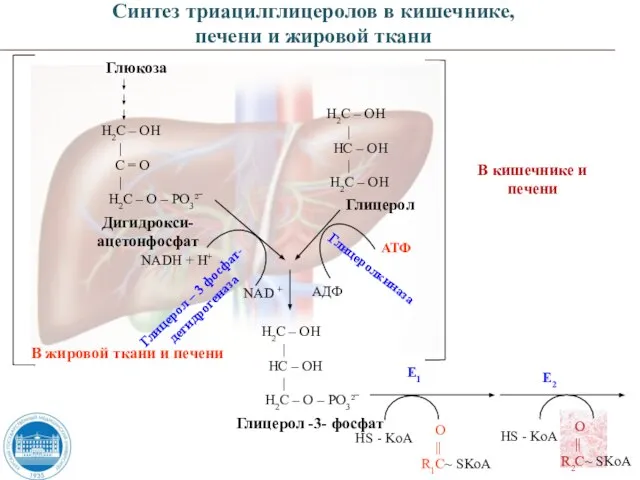
- 53. Глицерол-3-фосфат является субстратом для биосинтеза триацилглицеролов, который образуется из: глюкозы – печень, жировая ткань глицерола –
- 54. Глюкоза Н2С – ОН | С = О | Н2С – О – РО3²ˉ Дигидрокси- ацетонфосфат
- 56. Скачать презентацию
Слайд 2План лекции:
Особенности строения липидов.
Функции липидов.
Классификация липидов.
Строение ТАГ.
Переваривание и всасывание пищевых ТАГ.
Основные транспортные
План лекции:
Особенности строения липидов.
Функции липидов.
Классификация липидов.
Строение ТАГ.
Переваривание и всасывание пищевых ТАГ.
Основные транспортные

Слайд 3Липиды
Липиды – это разнообразная по строению группа органических молекул, имеющих общие
Липиды
Липиды – это разнообразная по строению группа органических молекул, имеющих общие
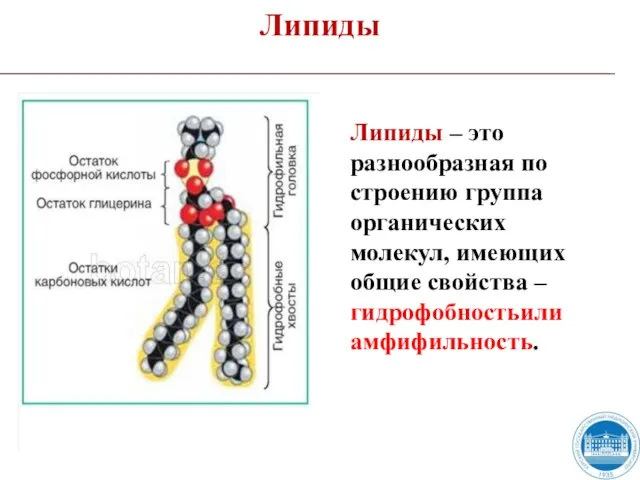
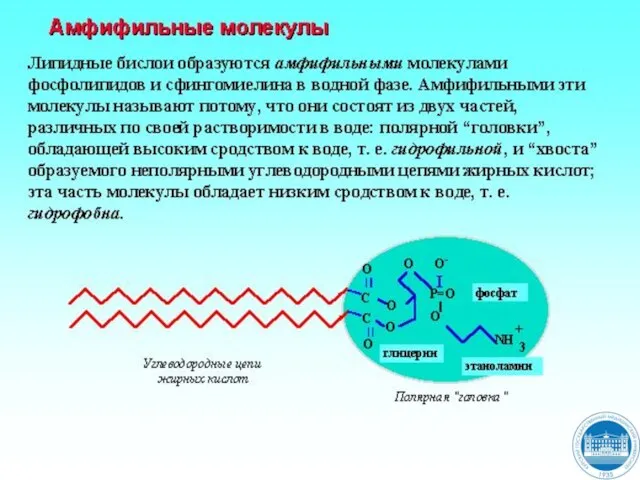
Слайд 5Функции липидов
Участвуют в формировании мембран: глицерофосфолипиды, сфинголипиды, холестерол;
Являются предшественниками коферментов: жирорастворимый витамин
Функции липидов
Участвуют в формировании мембран: глицерофосфолипиды, сфинголипиды, холестерол;
Являются предшественниками коферментов: жирорастворимый витамин

Слайд 6Строение и функции основных классов липидов человека
Строение и функции основных классов липидов человека
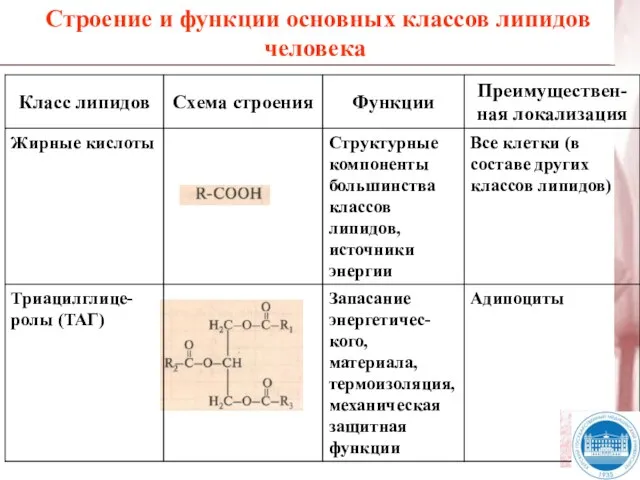
Слайд 7Строение и функции основных классов липидов человека
Строение и функции основных классов липидов человека

Слайд 8Строение и функции основных классов липидов человека
Строение и функции основных классов липидов человека
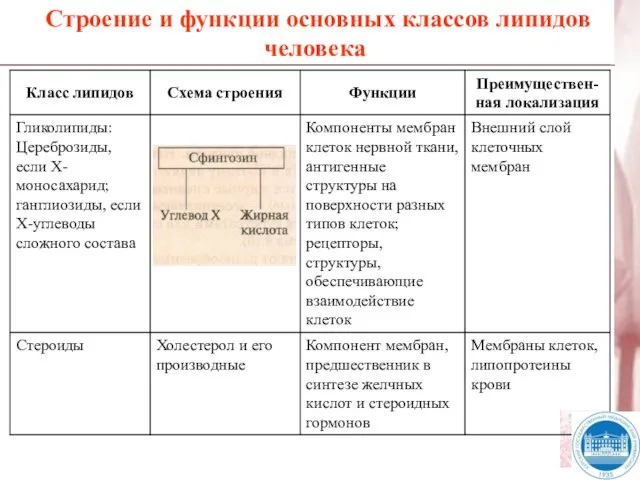
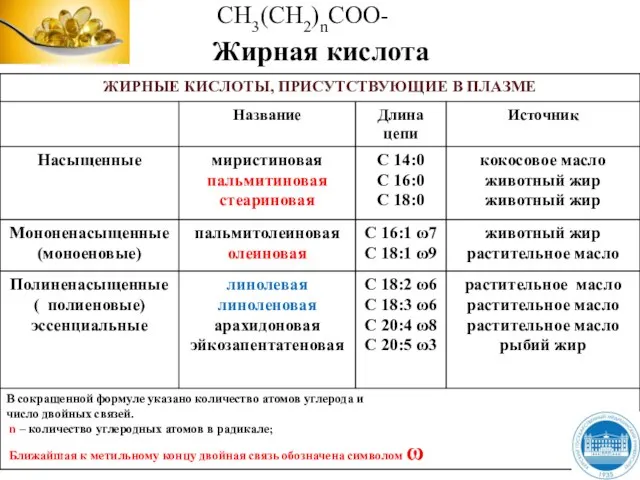
Слайд 9Жирные кислоты
Жирные кислоты
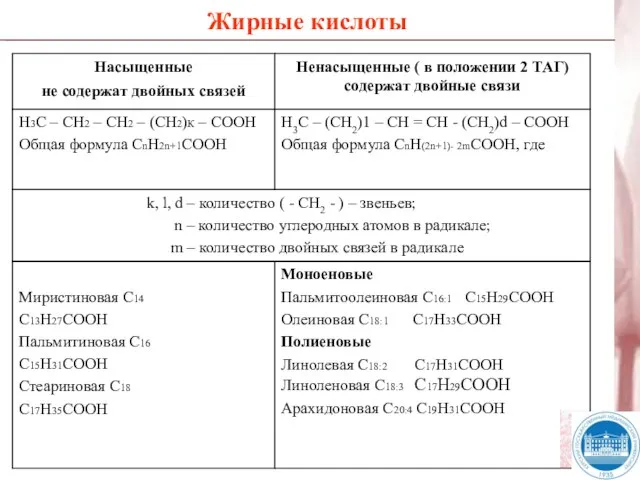
Слайд 11ТАГ (жиры) являются сложными эфирами жирных кислот и трехатомного спирта глицерола. К
ТАГ (жиры) являются сложными эфирами жирных кислот и трехатомного спирта глицерола. К

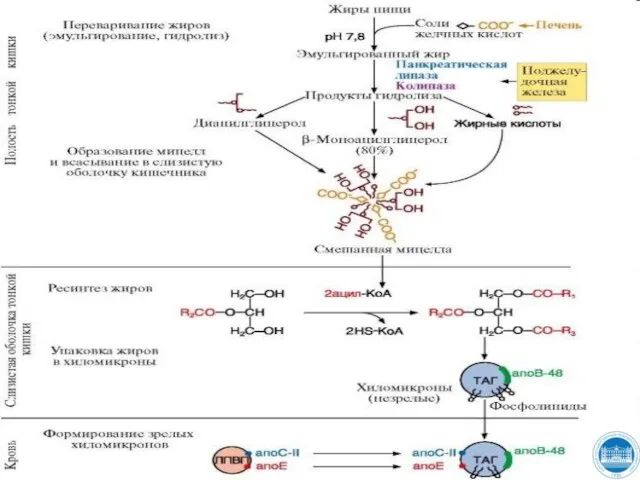
Слайд 12ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Основные липиды,
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Основные липиды,

Слайд 13ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Эмульгирование жиров
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Эмульгирование жиров

Слайд 14ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Действие панкреатической
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Действие панкреатической

Слайд 15ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Образование смешанных
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Образование смешанных

Слайд 16ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Нарушения переваривания
ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ЖИРОВ. РЕСИНТЕЗ ЖИРОВ В КЛЕТКАХ СЛИЗИСТОЙ ОБОЛОЧКИ КИШЕЧНИКА
Нарушения переваривания

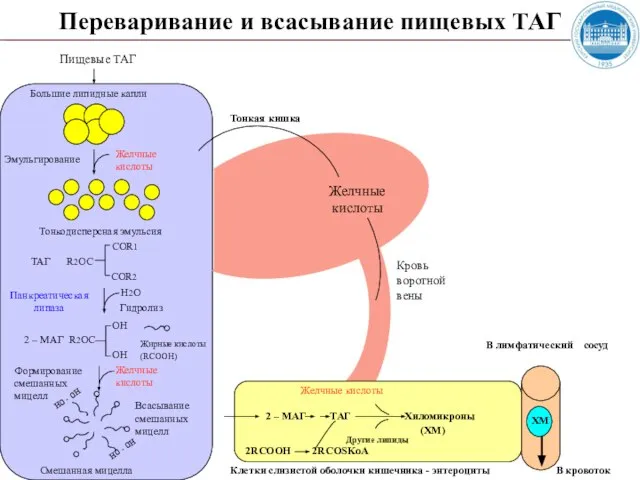
Слайд 18 Переваривание и всасывание пищевых ТАГ
Пищевые ТАГ
Большие липидные капли
Желчные кислоты
Эмульгирование
Тонкодисперсная
Переваривание и всасывание пищевых ТАГ
Пищевые ТАГ
Большие липидные капли
Желчные кислоты
Эмульгирование
Тонкодисперсная
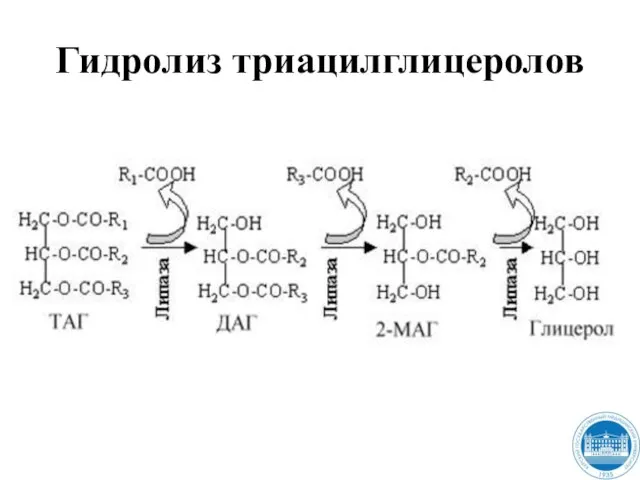
Слайд 19Гидролиз триацилглицеролов
Гидролиз триацилглицеролов
Слайд 20ХИЛОМИКРОНЫ - ТРАНСПОРТНАЯ ФОРМА ЭКЗОГЕННЫХ ЖИРОВ
Липиды, в частности жиры, холестерол и его
ХИЛОМИКРОНЫ - ТРАНСПОРТНАЯ ФОРМА ЭКЗОГЕННЫХ ЖИРОВ
Липиды, в частности жиры, холестерол и его

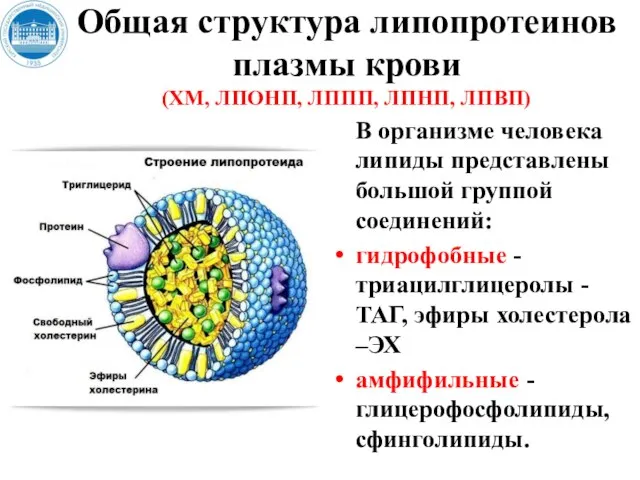
Слайд 21Общая структура липопротеинов плазмы крови
(ХМ, ЛПОНП, ЛППП, ЛПНП, ЛПВП)
В организме человека липиды
Общая структура липопротеинов плазмы крови
(ХМ, ЛПОНП, ЛППП, ЛПНП, ЛПВП)
В организме человека липиды
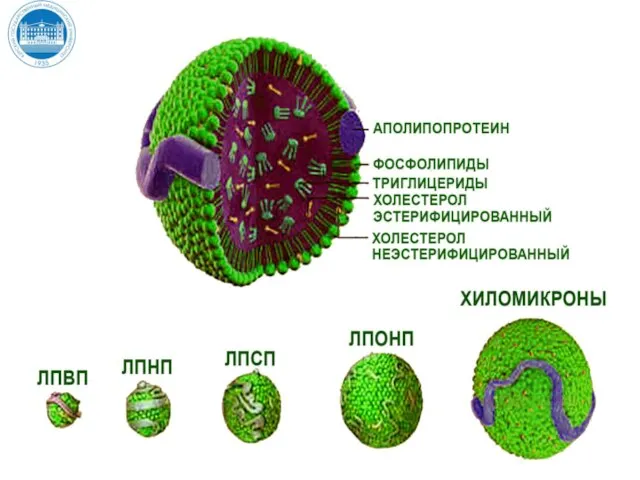
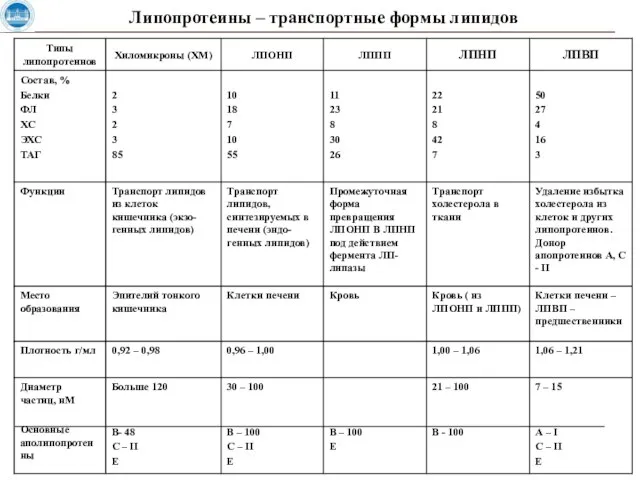
Слайд 23Липопротеины – транспортные формы липидов
Липопротеины – транспортные формы липидов
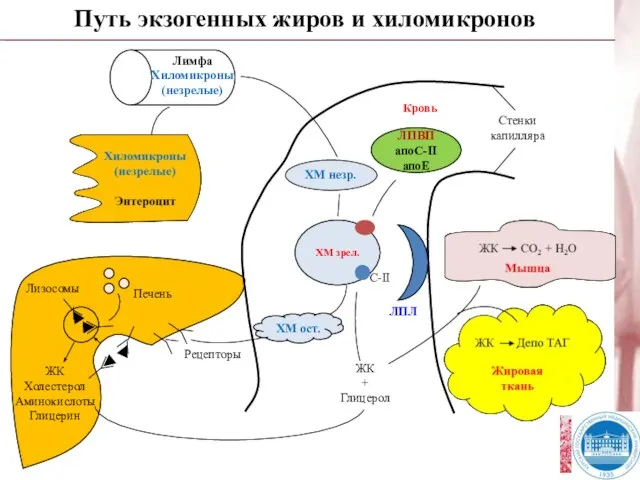
Слайд 24Путь экзогенных жиров и хиломикронов
Рецепторы
ХМ ост.
ХМ незр.
ЛПВП
апоС-II
апоЕ
Кровь
ЖК
+
Глицерол
Стенки
капилляра
Путь экзогенных жиров и хиломикронов
Рецепторы
ХМ ост.
ХМ незр.
ЛПВП
апоС-II
апоЕ
Кровь
ЖК
+
Глицерол
Стенки
капилляра
Слайд 25β – окисление жирных кислот – специфический путь катаболизма
β – окисление жирных кислот – специфический путь катаболизма

Слайд 26Характеристика β–окисления жирных кислот
β–окисление жирных кислот происходит только в аэробных условиях.
β–окисление жирных
Характеристика β–окисления жирных кислот
β–окисление жирных кислот происходит только в аэробных условиях.
β–окисление жирных

Слайд 27R – COOH + HS-KoA + АТФ
1-й этап - Активация жирных
R – COOH + HS-KoA + АТФ
1-й этап - Активация жирных
Слайд 28Наружная мембрана
Внутренняя мембрана
Цитозоль
R – C ~S-KoA
||
O
HS-KoA
R – C---
||
O
R
Наружная мембрана
Внутренняя мембрана
Цитозоль
R – C ~S-KoA
||
O
HS-KoA
R – C---
||
O
R
Слайд 293-й этап – Собственно β -окисление жирных кислот. Окисление протекает в матриксе
3-й этап – Собственно β -окисление жирных кислот. Окисление протекает в матриксе
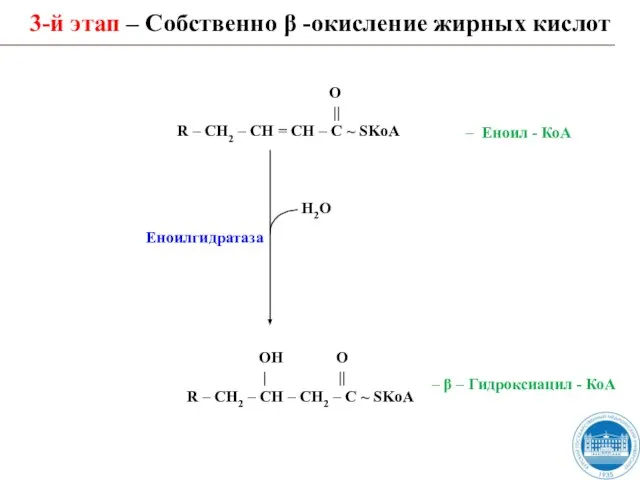
Слайд 303-й этап – Собственно β -окисление жирных кислот
Н2О
ОН О
| ||
3-й этап – Собственно β -окисление жирных кислот
Н2О
ОН О
| ||
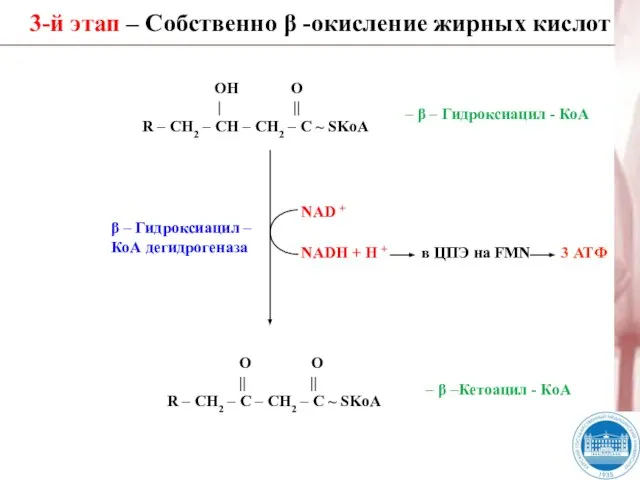
Слайд 313-й этап – Собственно β -окисление жирных кислот
β – Гидроксиацил –
КоА дегидрогеназа
3-й этап – Собственно β -окисление жирных кислот
β – Гидроксиацил –
КоА дегидрогеназа
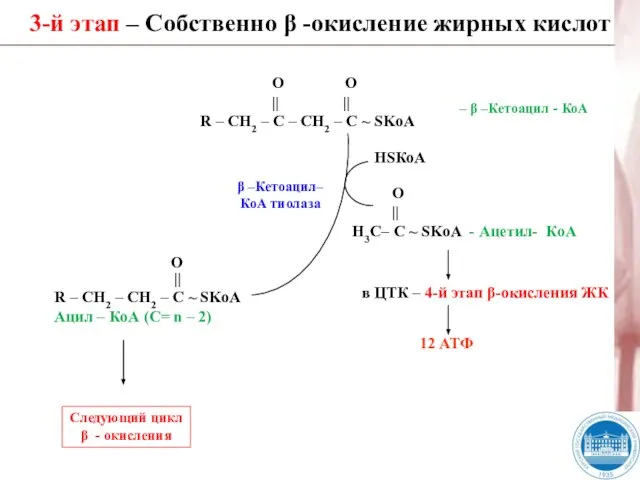
Слайд 323-й этап – Собственно β -окисление жирных кислот
β –Кетоацил–
КоА тиолаза
HSКоА
Следующий цикл
β
3-й этап – Собственно β -окисление жирных кислот
β –Кетоацил–
КоА тиолаза
HSКоА
Следующий цикл
β
Слайд 33Общая формула расчета баланса АТФ при β – окислении жирных кислот с
Общая формула расчета баланса АТФ при β – окислении жирных кислот с

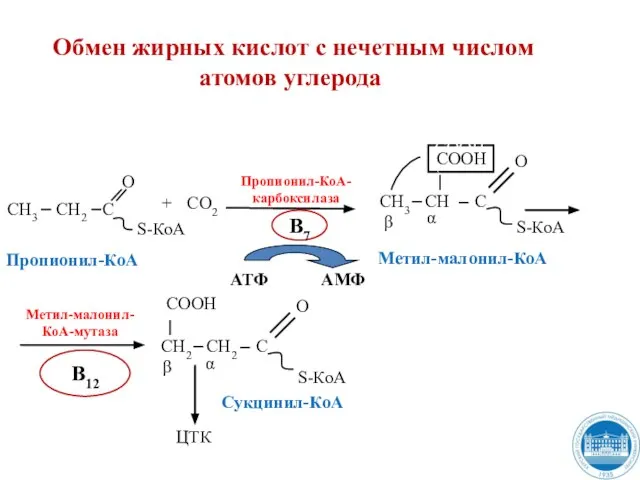
Слайд 34 Обмен жирных кислот с нечетным числом
атомов углерода
Метил-малонил-
КоА-мутаза
В12
АТФ
АМФ
Обмен жирных кислот с нечетным числом
атомов углерода
Метил-малонил-
КоА-мутаза
В12
АТФ
АМФ
Слайд 35Общая формула расчета баланса АТФ при
β – окислении жирных кислот с
Общая формула расчета баланса АТФ при β – окислении жирных кислот с

Слайд 36Биосинтез жирных кислот
Биосинтез жирных кислот

Слайд 37Отличия биосинтеза
жирных кислот от их окисления
Процесс протекает в цитоплазме клетки.
Идет с
Отличия биосинтеза
жирных кислот от их окисления
Процесс протекает в цитоплазме клетки.
Идет с

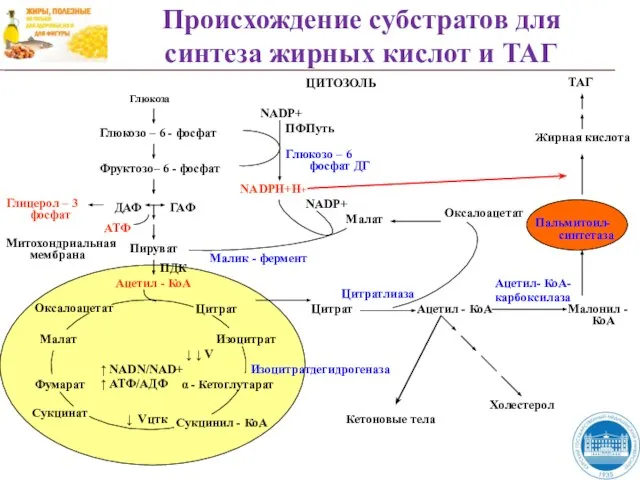
Слайд 38Происхождение субстратов для синтеза жирных кислот и ТАГ
Глюкоза
Глюкозо – 6 - фосфат
Фруктозо–
Происхождение субстратов для синтеза жирных кислот и ТАГ
Глюкоза
Глюкозо – 6 - фосфат
Фруктозо–
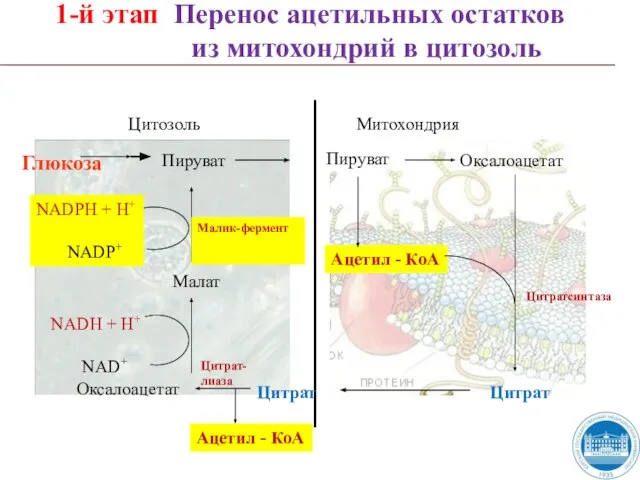
Слайд 39NADH + H+
NAD+
1-й этап Перенос ацетильных остатков
из митохондрий в цитозоль
NADH + H+
NAD+
1-й этап Перенос ацетильных остатков из митохондрий в цитозоль
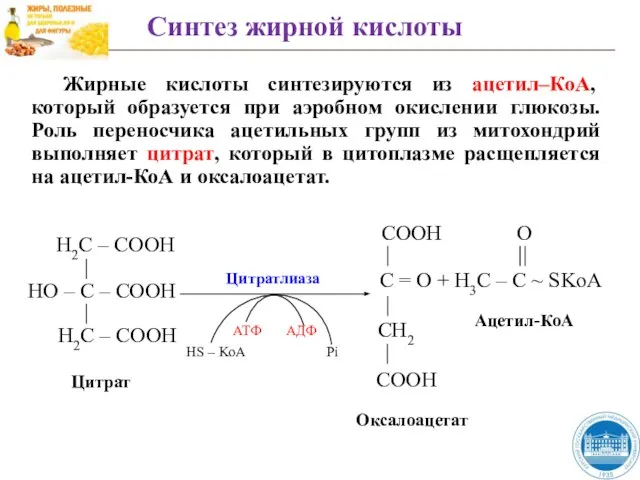
Слайд 40Жирные кислоты синтезируются из ацетил–КоА, который образуется при аэробном окислении глюкозы. Роль
Жирные кислоты синтезируются из ацетил–КоА, который образуется при аэробном окислении глюкозы. Роль
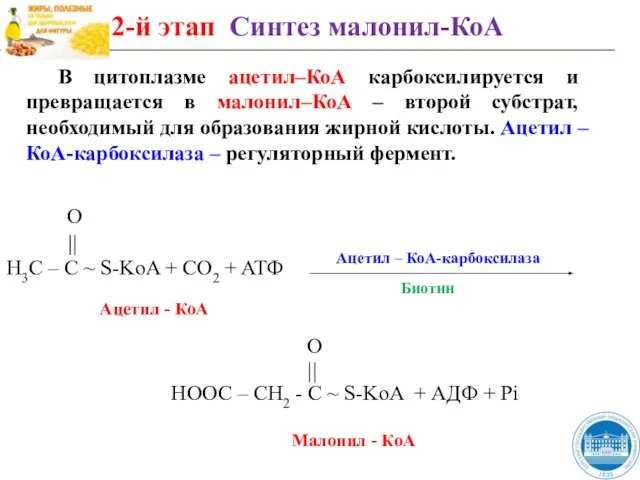
Слайд 41В цитоплазме ацетил–КоА карбоксилируется и превращается в малонил–КоА – второй субстрат, необходимый
В цитоплазме ацетил–КоА карбоксилируется и превращается в малонил–КоА – второй субстрат, необходимый
Слайд 42Регуляция активности ацетил – КоА-карбоксилазы
Ассоциация/диссоциация – объединение 3 тетрамеров в единый комплекс
Регуляция активности ацетил – КоА-карбоксилазы
Ассоциация/диссоциация – объединение 3 тетрамеров в единый комплекс
Слайд 433-й этап Синтез пальмитиновой кислоты
1
HS - KoA
HOOC – CH2 – CО ~
3-й этап Синтез пальмитиновой кислоты
1
HS - KoA
HOOC – CH2 – CО ~
Слайд 44Синтез пальмитиновой кислоты
СО2
Реакция конденсации
2
Синтез пальмитиновой кислоты
СО2
Реакция конденсации
2
Слайд 45Синтез пальмитиновой кислоты
Реакция восстановления
NADPH + H+
NADP+
3
Синтез пальмитиновой кислоты
Реакция восстановления
NADPH + H+
NADP+
3
Слайд 46Синтез пальмитиновой кислоты
H2O
Реакция дегидратации
4
Синтез пальмитиновой кислоты
H2O
Реакция дегидратации
4
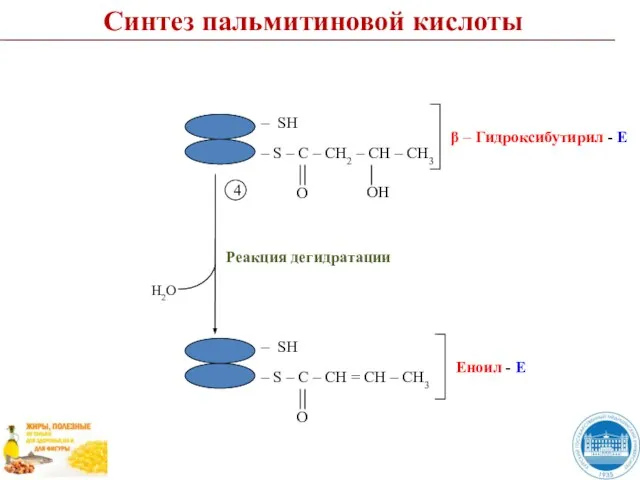
Слайд 47Синтез пальмитиновой кислоты
Реакция восстановления
5
NADP+
NADPH + H+
Синтез пальмитиновой кислоты
Реакция восстановления
5
NADP+
NADPH + H+
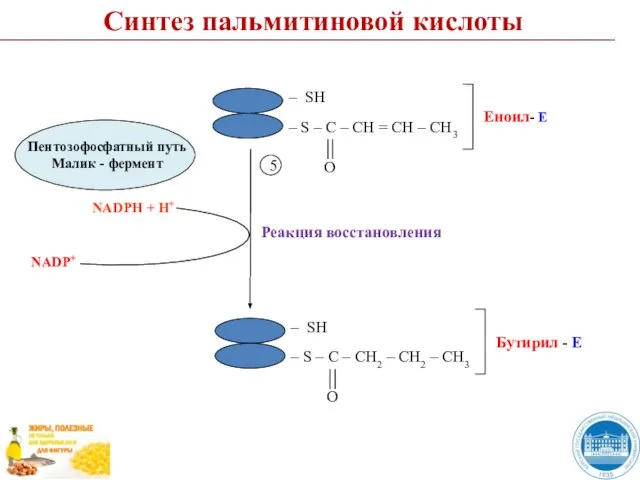
Слайд 48Синтез пальмитиновой кислоты
I цикл
6
НSKoA
Синтез пальмитиновой кислоты
I цикл
6
НSKoA
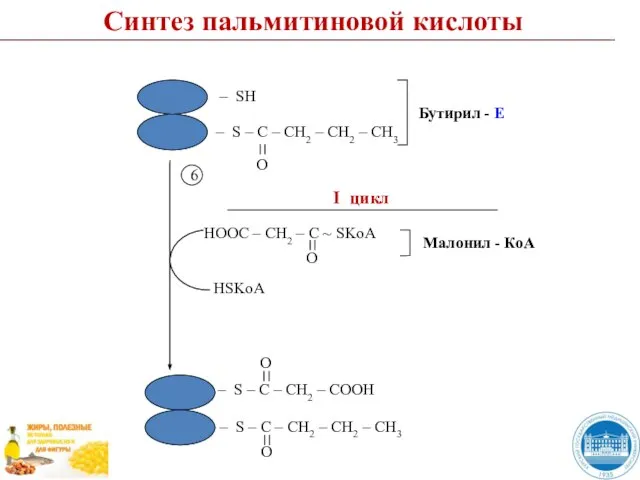
Слайд 497
CO2
Синтез пальмитиновой кислоты
7
CO2
Синтез пальмитиновой кислоты
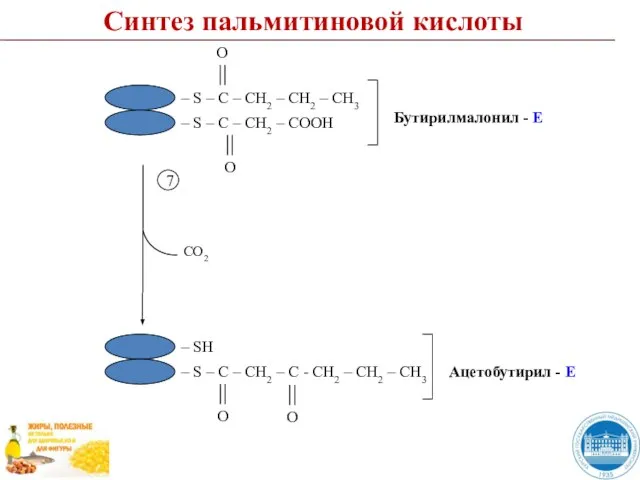
Слайд 50Пальмитиновая кислота (пальмитат)
Пальмитоил - Е
Н2О
Е
Синтез пальмитиновой кислоты
Пальмитиновая кислота (пальмитат)
Пальмитоил - Е
Н2О
Е
Синтез пальмитиновой кислоты
Слайд 51Ацетил - КоА + 7 Малонил – КоА + 14 (NADHPH +
Ацетил - КоА + 7 Малонил – КоА + 14 (NADHPH +
Слайд 52Биосинтез триацилглицеролов
Биосинтез триацилглицеролов

Слайд 53Глицерол-3-фосфат является субстратом для биосинтеза триацилглицеролов, который образуется из:
глюкозы – печень, жировая
Глицерол-3-фосфат является субстратом для биосинтеза триацилглицеролов, который образуется из:
глюкозы – печень, жировая

Слайд 54Глюкоза
Н2С – ОН
|
С = О
|
Н2С – О –
Глюкоза
Н2С – ОН
|
С = О
|
Н2С – О –
 Карбоновые кислоты
Карбоновые кислоты Окислительно-восстановительные реакции. Степень окисления
Окислительно-восстановительные реакции. Степень окисления Магний и его сплавы
Магний и его сплавы Сырьё для каменного литья и производства минеральной ваты
Сырьё для каменного литья и производства минеральной ваты Железо
Железо Углерод и его свойства
Углерод и его свойства Неметаллы. Агрегатные состояния неметаллов
Неметаллы. Агрегатные состояния неметаллов Физическая химия. Вводная лекция
Физическая химия. Вводная лекция Ионные уравнения
Ионные уравнения Основания. Классификация
Основания. Классификация Алкены. Строение
Алкены. Строение Синтез индолов по Неницеску
Синтез индолов по Неницеску Улеводы Моносахариды
Улеводы Моносахариды Простые и сложные вещества. Основные классы неорганических веществ
Простые и сложные вещества. Основные классы неорганических веществ Элементоорганические мономеры (соединения)
Элементоорганические мономеры (соединения) Синтетические высокомолекулярные органические вещества
Синтетические высокомолекулярные органические вещества Аммиак. Азот
Аммиак. Азот Химия в еде
Химия в еде Кислоты, их классификация и свойства
Кислоты, их классификация и свойства Коррозия металла
Коррозия металла Соединения галогенов
Соединения галогенов Химическая связь
Химическая связь Алюминий
Алюминий Презентация на тему Химическая связь (8 класс)
Презентация на тему Химическая связь (8 класс)  Металлы
Металлы Закономерности протеканий химических реакций
Закономерности протеканий химических реакций Биосинтез фенольных соединений в растениях
Биосинтез фенольных соединений в растениях Технологические особенности никеля и никелевых сплавов
Технологические особенности никеля и никелевых сплавов