- Интрондар. Ядролық мРНҚ гендерінің интрондары

Содержание
- 2. Интрондар жануар мен өсімдік геномында көп сипатталған, сонымен қатар олардың вирустарын инфекциялаушылардың геномында жақсы көрсетілген. Интрондарды
- 3. Интрондар мРНҚ, рРНҚ және тРНҚ жеке түрлерін кодтайтын ядролық генде болады; осы органеллалардың мРНҚ, рРНҚ және
- 4. Ядролық мРНҚ гендерінің интрондары Экзондардың өлшемі негізінен 52, 140, 223 және 299 н.ж көлемінде болады. Ядролық
- 6. Сплайсинг сайтында канондық тізбектің болуы осы интронның үнемі жойылатындығын білдірмейді. Кейбір жағдайда сплайсинг сайтының бір немесе
- 7. тРНҚ геніндегі интрондар тРНҚ геніндегі интрон өлшемі 14-тен шамамен 60 нуклеотид аралығында ауытқып тұрады, бірақ олар
- 9. I -ші топ интрондары I –ші топ интрондары кейбір төмен сатыдағы эукариоттардың ядролық рРНҚ генінде болады
- 10. I класс интрондарының құрылысының негізгі ерекшеліктері әрқайсысының ұзындығы шамамен 10 нуклеотидтен тұратын 4 нуклеотидті тізбек арқылы
- 12. I -ші кластың ядролық және митохондриялық интрондарының консервативті тізбегінің ұйымы
- 13. I I топ интрондары I I топ интрондары ашытқының қос митохондриялық генінен табылған. Олар цитохромоксидазаның және
- 14. 2‘,5‘-фосфодиэфирлі байланыстардың бір бірінен белгілі арақашықтықта орналасқан нуклеотидтерінің арасында табылуы
- 15. Автокатализдік сплайсингтің көмегімен кесілген интрондар Самосплайсинг – бұл реттелген процесс, ол фосфодиэфирлік алмасудың тізбекті автокатализдік реакциясынан
- 16. I топ интрондарының самосплайсингі 1. интронға гуанозиннің қосылуы. 2. байланысқан гуанозиннің 3’-гидроксилдік тобының бөлінбеген жұп электрондары
- 17. Ашытқылардың митохондриясының про-мРНҚ генінің I -ші топ кейбір интрондарындағыin vitro сплайсинг Tetrahymena про-рРНҚ сплайсингке қатысатын консервативті
- 20. I-ші топ интрондарының самосплайсингі, Tetrahymena thermophila рРНҚ интроны мысал ретінде
- 21. Интрондағы «ішкі бағытталған тізбек» (ол қанық емес боялған) екі экзон-интронды жүйені жақындастырады
- 22. Кесілген интронның альтернативті молекулаішілік жұптасуы одан олигонуклеотидтер мен циклизацияны алып тастайды.
- 23. II –ші топ интрондарының сплайсингі Сплайсинг механизмінің екі ерекшелігі: 1. II –топ интрондарының сплайсингі үшін нуклеозид-инициатор
- 25. Скачать презентацию
Слайд 2Интрондар жануар мен өсімдік геномында көп сипатталған, сонымен қатар олардың вирустарын инфекциялаушылардың
Интрондар жануар мен өсімдік геномында көп сипатталған, сонымен қатар олардың вирустарын инфекциялаушылардың

Слайд 3Интрондар мРНҚ, рРНҚ және тРНҚ жеке түрлерін кодтайтын ядролық генде болады; осы
Интрондар мРНҚ, рРНҚ және тРНҚ жеке түрлерін кодтайтын ядролық генде болады; осы

Слайд 4Ядролық мРНҚ гендерінің интрондары
Экзондардың өлшемі негізінен 52, 140, 223 және 299 н.ж
Ядролық мРНҚ гендерінің интрондары
Экзондардың өлшемі негізінен 52, 140, 223 және 299 н.ж

Слайд 6Сплайсинг сайтында канондық тізбектің болуы осы интронның үнемі жойылатындығын білдірмейді.
Кейбір жағдайда
Сплайсинг сайтында канондық тізбектің болуы осы интронның үнемі жойылатындығын білдірмейді.
Кейбір жағдайда

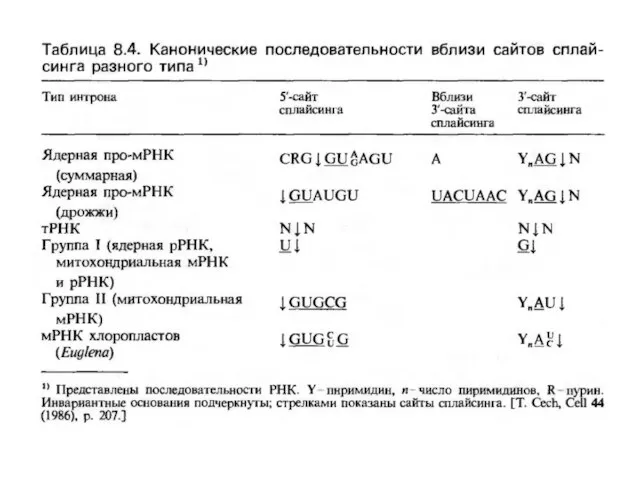
Слайд 7тРНҚ геніндегі интрондар
тРНҚ геніндегі интрон өлшемі 14-тен шамамен 60 нуклеотид аралығында ауытқып
тРНҚ геніндегі интрондар
тРНҚ геніндегі интрон өлшемі 14-тен шамамен 60 нуклеотид аралығында ауытқып

Слайд 9I -ші топ интрондары
I –ші топ интрондары кейбір төмен сатыдағы эукариоттардың ядролық
I -ші топ интрондары
I –ші топ интрондары кейбір төмен сатыдағы эукариоттардың ядролық

Слайд 10I класс интрондарының құрылысының негізгі ерекшеліктері әрқайсысының ұзындығы шамамен 10 нуклеотидтен тұратын
I класс интрондарының құрылысының негізгі ерекшеліктері әрқайсысының ұзындығы шамамен 10 нуклеотидтен тұратын


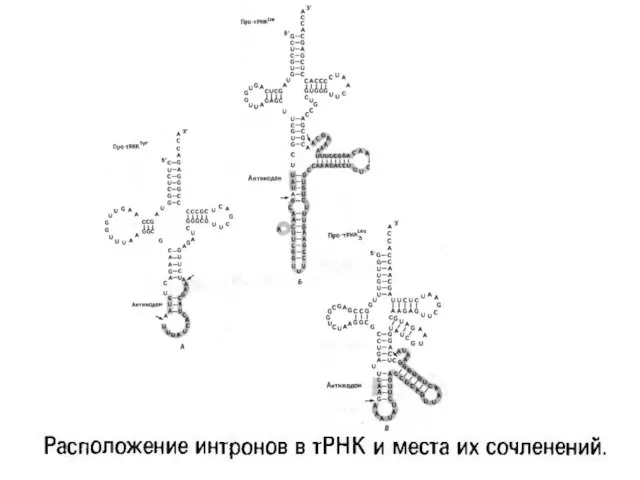
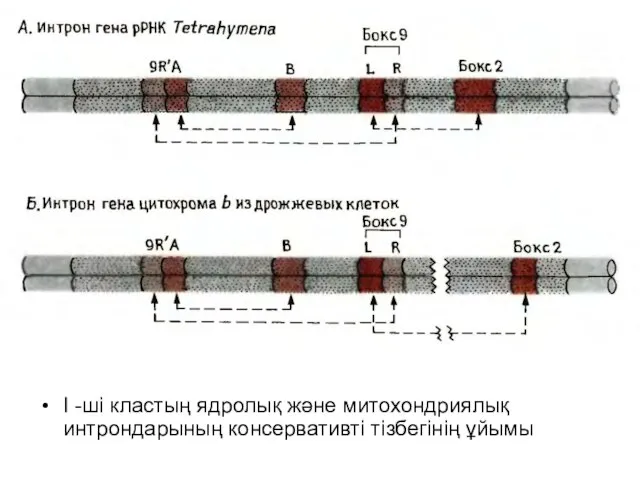
Слайд 12I -ші кластың ядролық және митохондриялық интрондарының консервативті тізбегінің ұйымы
I -ші кластың ядролық және митохондриялық интрондарының консервативті тізбегінің ұйымы
Слайд 13I I топ интрондары
I I топ интрондары ашытқының қос митохондриялық генінен табылған.
I I топ интрондары
I I топ интрондары ашытқының қос митохондриялық генінен табылған.

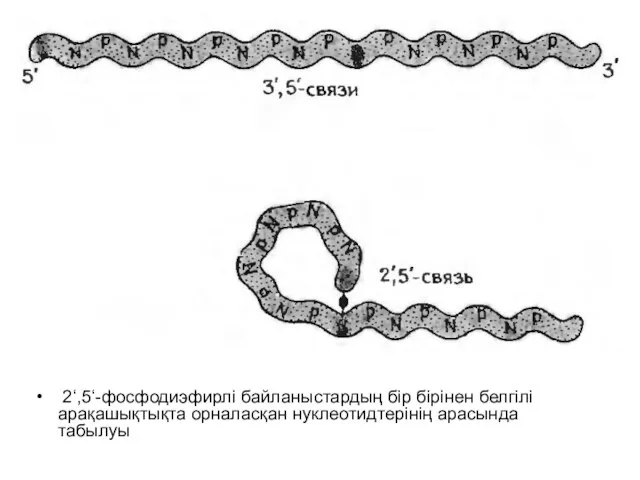
Слайд 14 2‘,5‘-фосфодиэфирлі байланыстардың бір бірінен белгілі арақашықтықта орналасқан нуклеотидтерінің арасында табылуы
2‘,5‘-фосфодиэфирлі байланыстардың бір бірінен белгілі арақашықтықта орналасқан нуклеотидтерінің арасында табылуы
Слайд 15Автокатализдік сплайсингтің көмегімен кесілген интрондар
Самосплайсинг – бұл реттелген процесс, ол фосфодиэфирлік алмасудың
Автокатализдік сплайсингтің көмегімен кесілген интрондар
Самосплайсинг – бұл реттелген процесс, ол фосфодиэфирлік алмасудың

Слайд 16I топ интрондарының самосплайсингі
1. интронға гуанозиннің қосылуы.
2. байланысқан гуанозиннің 3’-гидроксилдік тобының бөлінбеген
I топ интрондарының самосплайсингі
1. интронға гуанозиннің қосылуы.
2. байланысқан гуанозиннің 3’-гидроксилдік тобының бөлінбеген

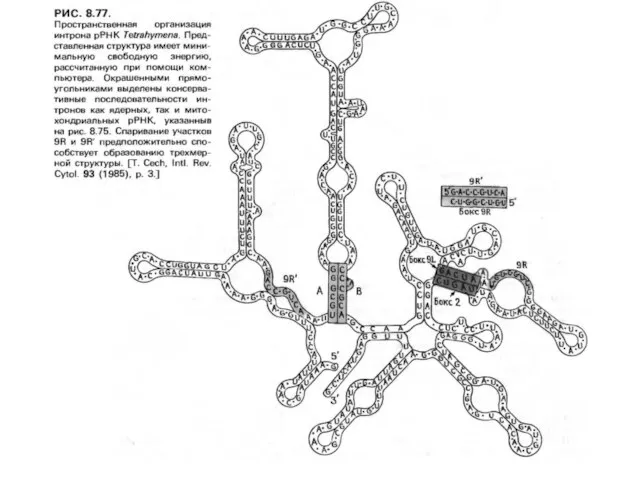
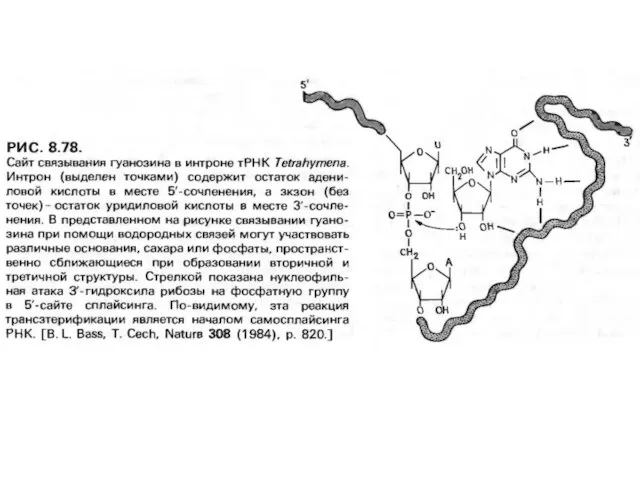
Слайд 17Ашытқылардың митохондриясының про-мРНҚ генінің I -ші топ кейбір интрондарындағыin vitro сплайсинг Tetrahymena
Ашытқылардың митохондриясының про-мРНҚ генінің I -ші топ кейбір интрондарындағыin vitro сплайсинг Tetrahymena

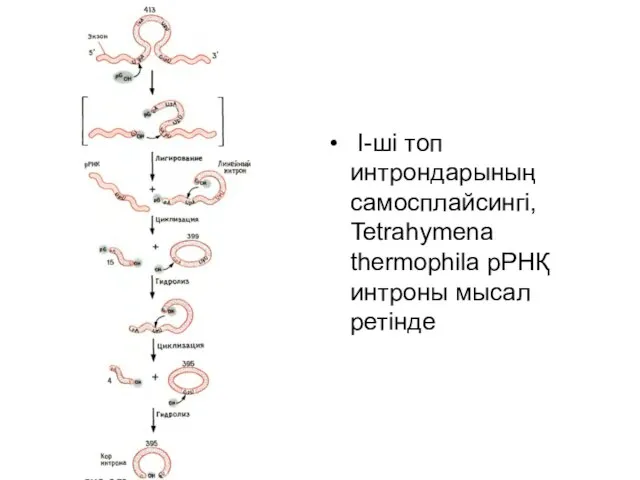
Слайд 20 I-ші топ интрондарының самосплайсингі, Tetrahymena thermophila рРНҚ интроны мысал ретінде
I-ші топ интрондарының самосплайсингі, Tetrahymena thermophila рРНҚ интроны мысал ретінде
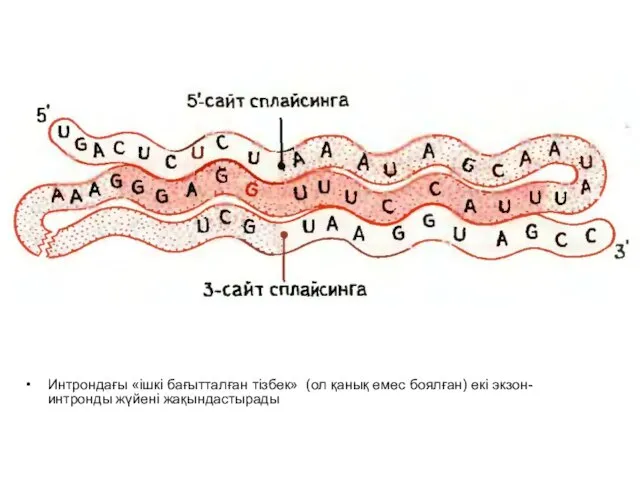
Слайд 21Интрондағы «ішкі бағытталған тізбек» (ол қанық емес боялған) екі экзон-интронды жүйені жақындастырады
Интрондағы «ішкі бағытталған тізбек» (ол қанық емес боялған) екі экзон-интронды жүйені жақындастырады
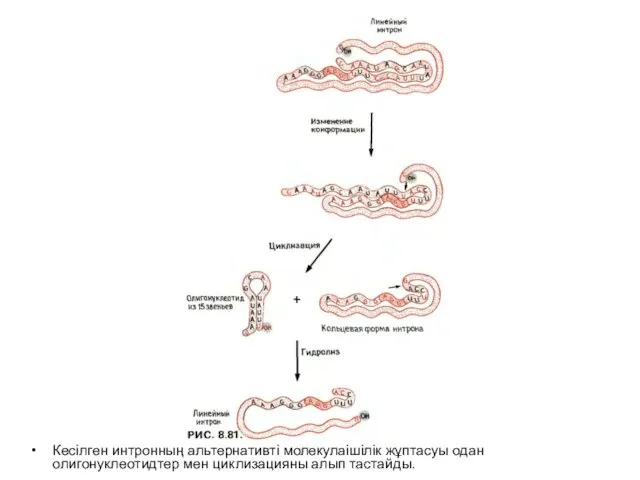
Слайд 22Кесілген интронның альтернативті молекулаішілік жұптасуы одан олигонуклеотидтер мен циклизацияны алып тастайды.
Кесілген интронның альтернативті молекулаішілік жұптасуы одан олигонуклеотидтер мен циклизацияны алып тастайды.
Слайд 23II –ші топ интрондарының сплайсингі
Сплайсинг механизмінің екі ерекшелігі:
1. II –топ интрондарының сплайсингі
II –ші топ интрондарының сплайсингі
Сплайсинг механизмінің екі ерекшелігі:
1. II –топ интрондарының сплайсингі

 Роль медсестры гастроэнтерологического отделения стационара в лечении заболеваний желудочно- кишечного тракта
Роль медсестры гастроэнтерологического отделения стационара в лечении заболеваний желудочно- кишечного тракта Пневмония 2021 Манищенкова
Пневмония 2021 Манищенкова Лечение гиперурикемии
Лечение гиперурикемии Профилактика инфекционных болезней плотоядных животных
Профилактика инфекционных болезней плотоядных животных Медицинское освидетельствование при первоначальной постановке на воинский учет
Медицинское освидетельствование при первоначальной постановке на воинский учет Трихинеллез животных
Трихинеллез животных Репродуктивные органы
Репродуктивные органы Черепно-мозговая травма. Протокол действий
Черепно-мозговая травма. Протокол действий Опорно-двигательный аппарат
Опорно-двигательный аппарат Концепция нейропсихологического синдрома и симптома
Концепция нейропсихологического синдрома и симптома Шизофрения. Продуктивная симптоматика
Шизофрения. Продуктивная симптоматика Нейровоспаление. Микроглия
Нейровоспаление. Микроглия Анэспум_ВОПРОСЫ-ОТВЕТЫ
Анэспум_ВОПРОСЫ-ОТВЕТЫ Операции при свищах и опухолях околоушной слюнной железы
Операции при свищах и опухолях околоушной слюнной железы История развития психопатологии в зарубежный странах
История развития психопатологии в зарубежный странах Род Corynebacterium
Род Corynebacterium Чума м,ясоїдних
Чума м,ясоїдних Брюшной тиф. Шигеллёз. Сестринское дело
Брюшной тиф. Шигеллёз. Сестринское дело профилактика 1 лекция
профилактика 1 лекция Общие положения о щитовидной железе. Лекция 5
Общие положения о щитовидной железе. Лекция 5 Особенноси ухода за больными с заболеваниями дыхательной системы
Особенноси ухода за больными с заболеваниями дыхательной системы Профилактика туберкулеза. Лекция №11
Профилактика туберкулеза. Лекция №11 Айырша бездің топографиялық анатомиясы. Балалардағы ерекшеліктері
Айырша бездің топографиялық анатомиясы. Балалардағы ерекшеліктері Экстренная реанимационная помощь
Экстренная реанимационная помощь Особенности клинических исследований в кардиологии
Особенности клинических исследований в кардиологии Помповая инсулинотерапия. Технические аспекты
Помповая инсулинотерапия. Технические аспекты дезінфекція2
дезінфекція2 Что такое СПИД
Что такое СПИД