- Механизмы клеточной смерти

Содержание
- 2. Апоптоз Регуляция клеточной пролиферации и дифференцировки, поддержание клеточного равновесия, элиминация дефектных клеток и клеток, достигших стадии
- 3. Основной закон клеточной кинетики состоит в том, что в единицу времени рождается и умирает одно и
- 4. Морфологические признаки апоптоза Морфологическими признаками апоптоза являются: конденсация хроматина, фрагментация ядра, буллезное (пузырчатое) выпячивание цитоплазматической мембраны.
- 5. Последовательность ультраструктурных изменений в процессе некроза и апоптоза
- 6. Морфологические признаки апоптоза Клетки разрушаются до апоптозных телец различного размера, сохраняющих целостность своей мембраны. В некоторых
- 7. Биологическая роль апоптоза Апоптоз играет биологически полезную роль в элиминации тех клеток, необходимость в которых отпала
- 8. Молекулярные механизмы апоптоза
- 9. Молекулярные механизмы апоптоза Выделяют 2 типа сигнальных путей, вызывающих апоптоз – повреждение ДНК, радиация и другие
- 10. Регуляция апоптоза Множество генов участвуют в регуляции апоптоза. Ведущую роль в осуществлении апоптоза играют ферменты -
- 11. Пролиферация и дифференцировка стволовых и коммитированных клеток-предшественников в нормальном кроветворении способна осуществляться только в условиях их
- 12. Ростовые факторы действуют на зрелые клетки, обеспечивая их выживание и нормальное функционирование. Воздействие ростовых факторов происходит
- 13. Некроз При некрозе гибель клеток происходит в результате физической или химической травмы, ишемии, гипертермии и других
- 15. Скачать презентацию
Слайд 2Апоптоз
Регуляция клеточной пролиферации и дифференцировки, поддержание клеточного равновесия, элиминация дефектных клеток и
Апоптоз
Регуляция клеточной пролиферации и дифференцировки, поддержание клеточного равновесия, элиминация дефектных клеток и

Слайд 3Основной закон клеточной кинетики состоит в том, что в единицу времени рождается
Основной закон клеточной кинетики состоит в том, что в единицу времени рождается

Слайд 4Морфологические признаки апоптоза
Морфологическими признаками апоптоза являются:
конденсация хроматина,
фрагментация ядра,
буллезное (пузырчатое)
Морфологические признаки апоптоза
Морфологическими признаками апоптоза являются:
конденсация хроматина,
фрагментация ядра,
буллезное (пузырчатое)

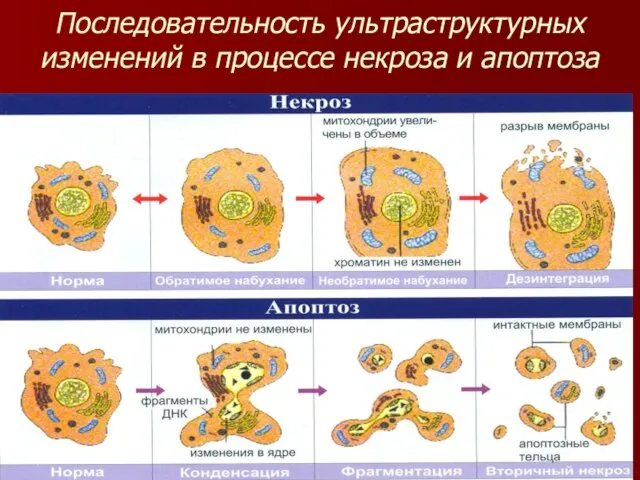
Слайд 5Последовательность ультраструктурных изменений в процессе некроза и апоптоза
Последовательность ультраструктурных изменений в процессе некроза и апоптоза
Слайд 6Морфологические признаки апоптоза
Клетки разрушаются до апоптозных телец различного размера, сохраняющих целостность своей
Морфологические признаки апоптоза
Клетки разрушаются до апоптозных телец различного размера, сохраняющих целостность своей

Слайд 7Биологическая роль апоптоза
Апоптоз играет биологически полезную роль в элиминации тех клеток, необходимость
Биологическая роль апоптоза
Апоптоз играет биологически полезную роль в элиминации тех клеток, необходимость

Слайд 8
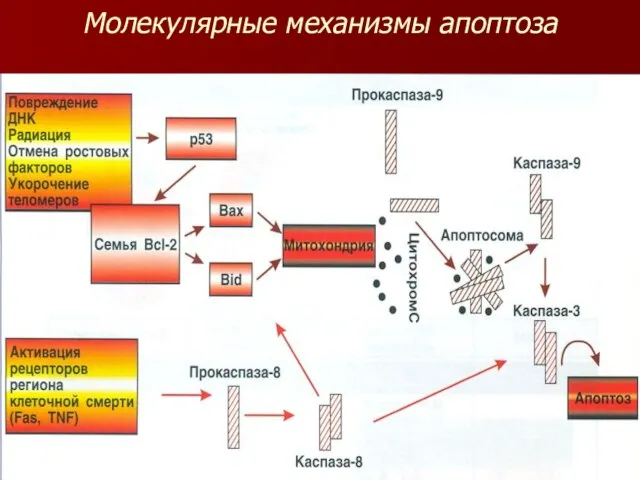
Молекулярные механизмы апоптоза
Молекулярные механизмы апоптоза
Слайд 9Молекулярные механизмы апоптоза
Выделяют 2 типа сигнальных путей, вызывающих апоптоз – повреждение ДНК,
Молекулярные механизмы апоптоза
Выделяют 2 типа сигнальных путей, вызывающих апоптоз – повреждение ДНК,

Слайд 10Регуляция апоптоза
Множество генов участвуют в регуляции апоптоза.
Ведущую роль в осуществлении апоптоза
Регуляция апоптоза
Множество генов участвуют в регуляции апоптоза.
Ведущую роль в осуществлении апоптоза

Слайд 11Пролиферация и дифференцировка стволовых и коммитированных клеток-предшественников в нормальном кроветворении способна осуществляться
Пролиферация и дифференцировка стволовых и коммитированных клеток-предшественников в нормальном кроветворении способна осуществляться

Слайд 12Ростовые факторы действуют на зрелые клетки, обеспечивая их выживание и нормальное функционирование.
Ростовые факторы действуют на зрелые клетки, обеспечивая их выживание и нормальное функционирование.

Слайд 13Некроз
При некрозе гибель клеток происходит в результате физической или химической травмы, ишемии,
Некроз
При некрозе гибель клеток происходит в результате физической или химической травмы, ишемии,

 Миссия Компании: «Мы делаем здоровый образ жизни» Миссия Компании – «Мы делаем здоровый бизнес» Millenium 2010 D4X My UnitDose ® - ПРОДУКТ БУДУЩЕГО. - презентация_
Миссия Компании: «Мы делаем здоровый образ жизни» Миссия Компании – «Мы делаем здоровый бизнес» Millenium 2010 D4X My UnitDose ® - ПРОДУКТ БУДУЩЕГО. - презентация_ Stabilometry
Stabilometry Травматизм, виды травм и их профилактика
Травматизм, виды травм и их профилактика Limfoscyntygrafia
Limfoscyntygrafia Закаливание и здоровье
Закаливание и здоровье Опиоидные аналгетики
Опиоидные аналгетики Стрептококки
Стрептококки Sour advice. LL - lemon’s lifehachs
Sour advice. LL - lemon’s lifehachs Желчно-каменная болезнь
Желчно-каменная болезнь Реабилитация и абилитация детей раннего возраста с нарушениями слуха
Реабилитация и абилитация детей раннего возраста с нарушениями слуха Фармакотерапия легочной гипертензии
Фармакотерапия легочной гипертензии Функциональная анатомия спинномозговых нервов
Функциональная анатомия спинномозговых нервов Гипертонические препараты
Гипертонические препараты Профилактика вирусных заболеваний
Профилактика вирусных заболеваний День работников Скорой помощи!
День работников Скорой помощи! Newtone. Профілактика простудних захворювань
Newtone. Профілактика простудних захворювань У меня родился ребенок!
У меня родился ребенок! История трансплантации легких. Хирургическая анатомия легких
История трансплантации легких. Хирургическая анатомия легких Правильное питание
Правильное питание Нервное перенапряжение. Причины, симптомы и способы разгрузки
Нервное перенапряжение. Причины, симптомы и способы разгрузки Зависимость от употребления энергетических напитков
Зависимость от употребления энергетических напитков Основы травматологии (шов сухожилия)
Основы травматологии (шов сухожилия) Выздоравливаем скорее! Рекомендации по реабилитации после ОРВИ
Выздоравливаем скорее! Рекомендации по реабилитации после ОРВИ ИКСИ-технология
ИКСИ-технология Коринебактерии
Коринебактерии Как дожить до офтальмолога
Как дожить до офтальмолога Поведение в родах
Поведение в родах Инфицированный аборт
Инфицированный аборт