- Жануарлар клеткалық культурасын алу және өсіру түрлері

Содержание
- 2. Жоспары: І. Кіріспе Әдіс тарихы ІІ. Негізгі бөлім 1. Клеткаларды культивирлеу 2. Культураға клетканы енгізу 3.
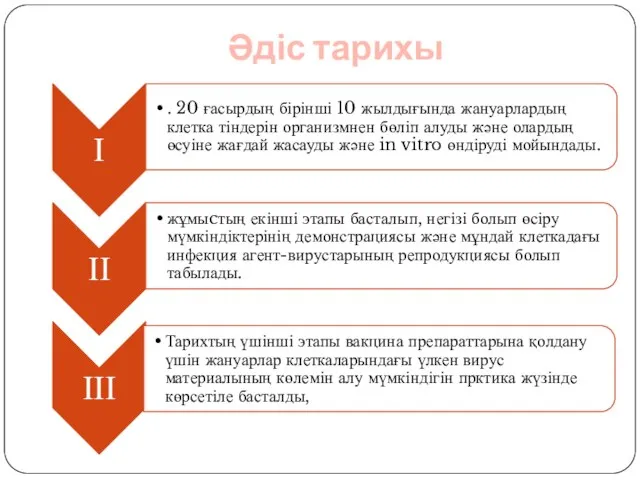
- 3. Әдіс тарихы
- 4. ІІІ этап Үлкен вирус материалының көлемін алу мүмкіндігін прктика жүзінде көрсетіле басталды, және жойылуы: 1) клеткаға
- 5. Культурада жануар клеткаларының өсу және бөліну қасиетін көрсету үшін, әдістер қатарын меңгеру қажет етілді. 1. Экзогенді
- 6. Осы әдістемелердің разработкасының ғылыми негізін, жануар және өсімдік тектес тірі организмдердің құрылымдық элементі ретінде клетка құрайды.
- 7. Одан кейінірек 1885 жылы У. Рукс организмнен тыс тірі организмдерді сақтайһу мүмкіншілігін практика жүзінде көрсетті. Ол
- 8. 1913ж Алексис Каррель эмбрион экстрактысымен байытылған қан плазмасын қолданды. Бұл әдістеме үлкен жетістікті қамтамасыз етті. Рид
- 9. Каррель хирург болғандықтан және асептика жөнінде білгендіктен жануарлардың клеткаларын in vitro культивирлеуге үлкен үлесін тигізді. Каррель
- 11. Культураға клетканы енгізу Экспериментальды жұмыстың мақсаты мен тапсырмаларына қатысты жануар клеткаларының 2 бағытта культивирленуін шығаруға болады:
- 12. Клеткалар культуралары құрылымдық мекемелерден алыстатылған гистиотипикалық архитектураға тән қасиетін биохимиялық белгілерін жоғалтады және олар арнайы жағдайлардың
- 13. Клеткалар түрінің культураға кірген тізімі жетерліктей көп. Олар: адамның қосалқы тін элементі (фибробластар), скелет тіндері (сүйек
- 15. Клеткалар популяциясы үнемі гомогенді болмайды және фиксирленген фенотипі бар. Культураға енгізуге қандай ті алуға болады, ересек
- 16. Жаңа бөлінген культуралар пассирлеу мен субкуьтурлеубасталғанға дейін біріншілік культура деген атқа ие болады. Біріншілік культуралар клеткалары
- 17. Іn vitro жағдайында культивирленетін клеткалар сипаттамасы Бір типтегі клеткалар ұстап тұру үшін тіндердеәрекеттеседі және бөліну жылдамдығын
- 18. Адгезияда субстрат көпвалентті антидене ретінде әсер етеді. Ал көп түзілген белгілерді «адгезивті дақ» деп атайды. Бұл
- 19. Қалыпты клеткалар бөлінуді тоқтатса, бұл құбылыс тығыздыққа байланысты пролиферацияның тоқтауымен түсіндіріледі.Егер мұндай моноқабат атбақшада клеткадан бос
- 21. Өсу факторы әдетте концентрациясы 10-10 М болатын ортада болады. Өсу факторы бір фибробласта 105 рецепторлы өсу
- 22. Өсу факторы мен қоректік орта бәсекелестігі клетка культурасындағы бөлу жылдамдығына әсер ететін жалғыз фактор емес. Субстрат
- 23. Бірақ та субстрат аймағымен байланысқа түсе сала, клетка распластай алмайтындай бұл аймақ өте кішкентай болса да
- 24. Қоректік орта Тіннен немесе ағзадан клеткаларды алып оларды культураға орналастырған соң, культуральды орта клеткалары in vivo
- 25. Адам және жануарлардың клетка культурасы сұйық (қоректік орта) газообразды (газ концентрациясы) және қатты (субстрат беті) фазада,
- 26. Қоректік ортаны дайындау үшін Эрл мен Хенкстің тұ ерітіндісі қолданылады. Бұл ерітінділер фосфаттұзды буфер Дубленко мен
- 27. Мысалы, Эрл ерітіндісі үшін осмолярлықтың есептік шамасы. 310,6 мосмоль/кг шындығында 283 клетка көбеюінен шығатын pН диапазоны
- 28. Жануарлар клеткасы культурасын жүргізу үшін стандарттық орта. Игл орталары МЕМ (minimal essetial medium) және ВМЕ (basal
- 29. Исков IMDM – Дульбекко ортасы модификациясы. алмастырылмайтын амиқышқылдары, биотин, В12 витамині, натрий селениті қосылады. Ортаға HEPES
- 31. Скачать презентацию
Слайд 2Жоспары:
І. Кіріспе
Әдіс тарихы
ІІ. Негізгі бөлім
1. Клеткаларды культивирлеу
2. Культураға клетканы енгізу
3. Клеткалардың шығуы
4.
Жоспары:
І. Кіріспе
Әдіс тарихы
ІІ. Негізгі бөлім
1. Клеткаларды культивирлеу
2. Культураға клетканы енгізу
3. Клеткалардың шығуы
4.

Слайд 3Әдіс тарихы
Әдіс тарихы
Слайд 4ІІІ этап
Үлкен вирус материалының көлемін алу мүмкіндігін прктика жүзінде көрсетіле басталды, және
ІІІ этап
Үлкен вирус материалының көлемін алу мүмкіндігін прктика жүзінде көрсетіле басталды, және

Слайд 5 Культурада жануар клеткаларының өсу және бөліну қасиетін көрсету үшін, әдістер қатарын
Культурада жануар клеткаларының өсу және бөліну қасиетін көрсету үшін, әдістер қатарын

Слайд 6 Осы әдістемелердің разработкасының ғылыми негізін, жануар және өсімдік тектес тірі организмдердің
Осы әдістемелердің разработкасының ғылыми негізін, жануар және өсімдік тектес тірі организмдердің

Слайд 7 Одан кейінірек 1885 жылы У. Рукс организмнен тыс тірі организмдерді сақтайһу
Одан кейінірек 1885 жылы У. Рукс организмнен тыс тірі организмдерді сақтайһу

Слайд 81913ж Алексис Каррель эмбрион экстрактысымен байытылған қан плазмасын қолданды. Бұл әдістеме үлкен
1913ж Алексис Каррель эмбрион экстрактысымен байытылған қан плазмасын қолданды. Бұл әдістеме үлкен

Слайд 9 Каррель хирург болғандықтан және асептика жөнінде білгендіктен жануарлардың клеткаларын in vitro
Каррель хирург болғандықтан және асептика жөнінде білгендіктен жануарлардың клеткаларын in vitro


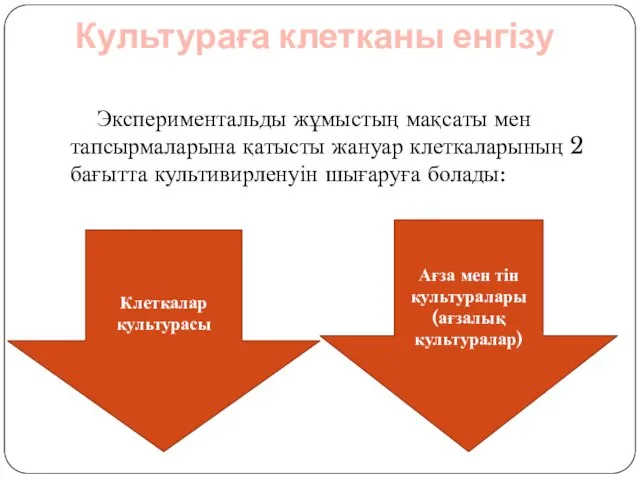
Слайд 11Культураға клетканы енгізу
Экспериментальды жұмыстың мақсаты мен тапсырмаларына қатысты жануар клеткаларының 2
Культураға клетканы енгізу
Экспериментальды жұмыстың мақсаты мен тапсырмаларына қатысты жануар клеткаларының 2
Слайд 12 Клеткалар культуралары құрылымдық мекемелерден алыстатылған гистиотипикалық архитектураға тән қасиетін биохимиялық белгілерін
Клеткалар культуралары құрылымдық мекемелерден алыстатылған гистиотипикалық архитектураға тән қасиетін биохимиялық белгілерін

Слайд 13Клеткалар түрінің культураға кірген тізімі жетерліктей көп. Олар:
адамның қосалқы тін элементі
Клеткалар түрінің культураға кірген тізімі жетерліктей көп. Олар:
адамның қосалқы тін элементі

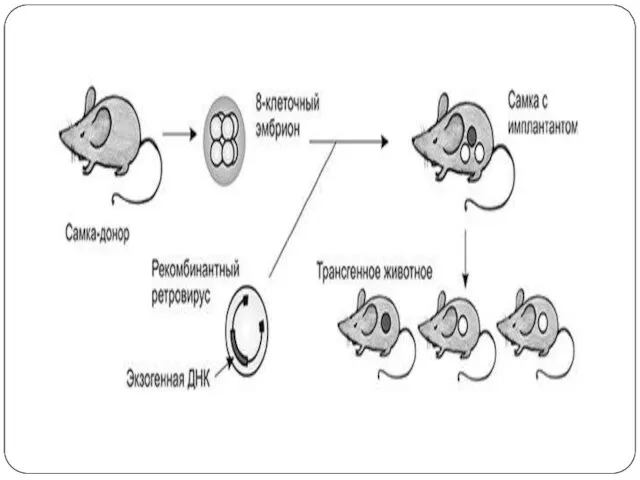
Слайд 15Клеткалар популяциясы үнемі гомогенді болмайды және фиксирленген фенотипі бар. Культураға енгізуге қандай
Клеткалар популяциясы үнемі гомогенді болмайды және фиксирленген фенотипі бар. Культураға енгізуге қандай

Слайд 16Жаңа бөлінген культуралар пассирлеу мен субкуьтурлеубасталғанға дейін біріншілік культура деген атқа ие
Жаңа бөлінген культуралар пассирлеу мен субкуьтурлеубасталғанға дейін біріншілік культура деген атқа ие

Слайд 17Іn vitro жағдайында культивирленетін клеткалар сипаттамасы
Бір типтегі клеткалар ұстап тұру үшін
Іn vitro жағдайында культивирленетін клеткалар сипаттамасы
Бір типтегі клеткалар ұстап тұру үшін

Слайд 18Адгезияда субстрат көпвалентті антидене ретінде әсер етеді. Ал көп түзілген белгілерді «адгезивті
Адгезияда субстрат көпвалентті антидене ретінде әсер етеді. Ал көп түзілген белгілерді «адгезивті

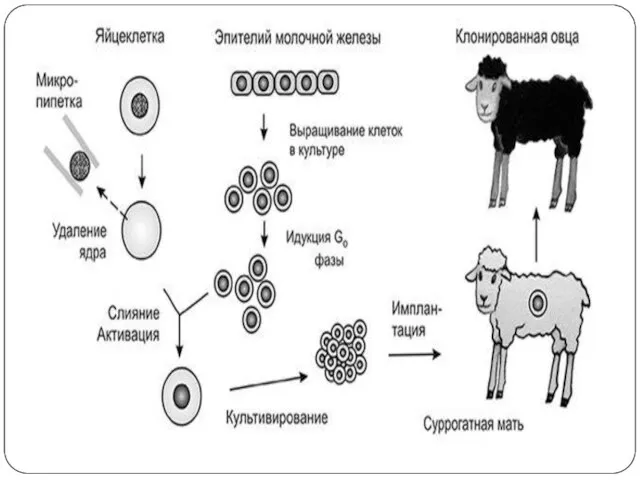
Слайд 19Қалыпты клеткалар бөлінуді тоқтатса, бұл құбылыс тығыздыққа байланысты пролиферацияның тоқтауымен түсіндіріледі.Егер мұндай
Қалыпты клеткалар бөлінуді тоқтатса, бұл құбылыс тығыздыққа байланысты пролиферацияның тоқтауымен түсіндіріледі.Егер мұндай

Слайд 21Өсу факторы әдетте концентрациясы 10-10 М болатын ортада болады. Өсу факторы бір
Өсу факторы әдетте концентрациясы 10-10 М болатын ортада болады. Өсу факторы бір

Слайд 22Өсу факторы мен қоректік орта бәсекелестігі клетка культурасындағы бөлу жылдамдығына әсер ететін
Өсу факторы мен қоректік орта бәсекелестігі клетка культурасындағы бөлу жылдамдығына әсер ететін

Слайд 23Бірақ та субстрат аймағымен байланысқа түсе сала, клетка распластай алмайтындай бұл аймақ
Бірақ та субстрат аймағымен байланысқа түсе сала, клетка распластай алмайтындай бұл аймақ

Слайд 24 Қоректік орта
Тіннен немесе ағзадан клеткаларды алып оларды культураға орналастырған соң, культуральды
Қоректік орта
Тіннен немесе ағзадан клеткаларды алып оларды культураға орналастырған соң, культуральды

Слайд 25Адам және жануарлардың клетка культурасы сұйық (қоректік орта) газообразды (газ концентрациясы) және
Адам және жануарлардың клетка культурасы сұйық (қоректік орта) газообразды (газ концентрациясы) және

Слайд 26Қоректік ортаны дайындау үшін Эрл мен Хенкстің тұ ерітіндісі қолданылады. Бұл ерітінділер
Қоректік ортаны дайындау үшін Эрл мен Хенкстің тұ ерітіндісі қолданылады. Бұл ерітінділер

Слайд 27Мысалы, Эрл ерітіндісі үшін осмолярлықтың есептік шамасы. 310,6 мосмоль/кг шындығында 283 клетка
Мысалы, Эрл ерітіндісі үшін осмолярлықтың есептік шамасы. 310,6 мосмоль/кг шындығында 283 клетка

Слайд 28Жануарлар клеткасы культурасын жүргізу үшін стандарттық орта.
Игл орталары МЕМ (minimal essetial medium)
Жануарлар клеткасы культурасын жүргізу үшін стандарттық орта.
Игл орталары МЕМ (minimal essetial medium)

Слайд 29Исков IMDM – Дульбекко ортасы модификациясы. алмастырылмайтын амиқышқылдары, биотин, В12 витамині, натрий
Исков IMDM – Дульбекко ортасы модификациясы. алмастырылмайтын амиқышқылдары, биотин, В12 витамині, натрий

 Морфофункциональная характеристика отделов скелета и аппарата их движения. Тема 7
Морфофункциональная характеристика отделов скелета и аппарата их движения. Тема 7 Ревматическая лихорадка на Северном Кавказе: вчера, сегодня, завтра
Ревматическая лихорадка на Северном Кавказе: вчера, сегодня, завтра Депрескрайбинг. Практика снижения дозы и отмены лекарственных средств
Депрескрайбинг. Практика снижения дозы и отмены лекарственных средств Витамины
Витамины Емізіктің жарақаттануы және лактостаз
Емізіктің жарақаттануы және лактостаз Синдром Эдвардса
Синдром Эдвардса Цереброваскулярные болезни
Цереброваскулярные болезни Остановка сердца
Остановка сердца Метрология и лабораторная медицина
Метрология и лабораторная медицина Направление медицинского туризма внутри и вне РФ
Направление медицинского туризма внутри и вне РФ Приспособительные реакции и компенсация утраченных функций организмов
Приспособительные реакции и компенсация утраченных функций организмов Золотые нити в косметологии
Золотые нити в косметологии Интеллект и интеллектуальная недостаточность. Признаки
Интеллект и интеллектуальная недостаточность. Признаки Biocad в цифрах
Biocad в цифрах Эндодонтия. Лекция № 2
Эндодонтия. Лекция № 2 Правда о СПИДе
Правда о СПИДе Лечебная физкультура при бронхиальной астме
Лечебная физкультура при бронхиальной астме Топографическая анатомия брюшной полости, операции на печени и желчных путях
Топографическая анатомия брюшной полости, операции на печени и желчных путях Сестринский уход. Документация к сестринскому уходу
Сестринский уход. Документация к сестринскому уходу Назальная цитология у детей с бронхиальной астмой
Назальная цитология у детей с бронхиальной астмой Алгоритм локализации области фиброаденом на маммограмме
Алгоритм локализации области фиброаденом на маммограмме Школа здоровья для беременных. Занятие 4. Возможные проблемы
Школа здоровья для беременных. Занятие 4. Возможные проблемы Препараты применяемые в детской гастроэнтрологии
Препараты применяемые в детской гастроэнтрологии Химический состав шоколада
Химический состав шоколада Опухоли грудной полости у детей
Опухоли грудной полости у детей Целиакия
Целиакия Регенерация и старение
Регенерация и старение Цистит. Виды цистита. Лечение цистита
Цистит. Виды цистита. Лечение цистита