- АМИНОКИСЛОТЫ Аминокислоты – соединения, содержащие карбоксильную (COOH) и аминогруппу (NH2).

Содержание
- 2. 1. Классификация 1.1. по положению аминогруппы 25.04.2012
- 3. 1.2. По количеству карбокси- и аминогрупп Моноаминомонокарбоновые кислоты (глицин, аланин, валин, лейцин, изолейцин, серин, треонин, цистеин,
- 4. 1.3 Классификация по встречаемости в белках 25.04.2012 20 классических протеиногенных аминокислот, информация о положении которых в
- 5. 1.4. По пищевой ценности для человека Аминокислоты делятся на заменимые и незаменимые. К незаменимым аминокислотам относят:
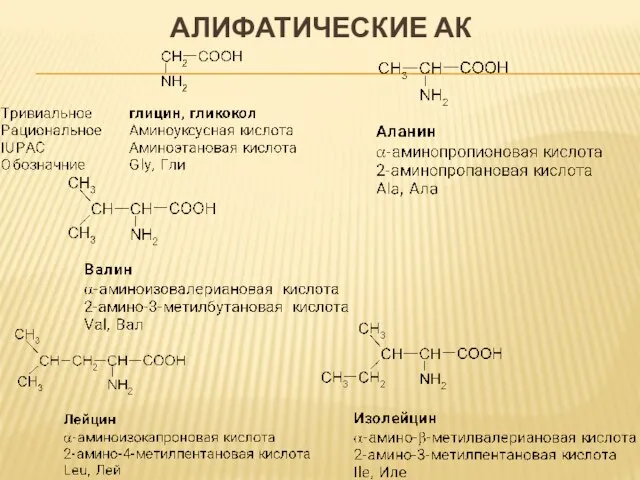
- 6. АЛИФАТИЧЕСКИЕ АК
- 7. СОДЕРЖАЩИЕ ОН-ГРУППУ Ceрин α-амино-β-оксипропионовая кислота 2-амино-3-гидроксипропановая кислота Ser, Сeр Трeонин α-амино-β-оксимасляная кислота 2-амино-3-гидроксибутановая кислота Thr, Трe
- 8. СЕРОСОДЕРЖАЩИЕ АК Цистеин Цистин Мeтионин α-амино-γ-мeтилтиомасляная кислота 2-амино-4-метилсульфанилбутановая кислота (2-амино-4-метилтиобутановая кислота – устаревш.) Met, Мет. Цистeин
- 9. МОНОАМИНОДИКАРБОНОВЫЕ КИСЛОТЫ И ИХ АМИДЫ Аспарагиновая кислота Аминоянтарная кислота Аминобутандиовая кислота Asp, Асп Глутаминовая кислота α-aминоглутаровая
- 10. СОДЕРЖАЩИЕ АМИНОГРУППУ Лизин α,ε-диаминокапроновая кислота 2,6-диаминогексановая кислота Lys, Лиз Аргинин α-амино-δ-гуанидилвалериановая кислота 2-амино-5-[амино(имино)метил]аминопентановая к-та Arg, Арг
- 11. АРОМАТИЧЕСКИЕ АК Фенилаланин α-амино-β-фенилпропионовая к-та 2-амино-3-фенилпропановая к-та Phe, Фен Тирозин α-амино-β-(п-оксифенил)пропионовая к-та 2-амино-3-(4-гидроксифенил)пропановая к-та Tyr, Тир
- 12. ГЕТЕРОЦИКЛИЧЕСКИЕ АК Триптофан α-амино-β-индолилпропионовая к-та 2-амино-3-(1H-индол-3-ил)пропановая к-та Trp, Три Гистидин α-амино-β-имидазолилпропионовая к-та 2-амино-3-(1H-имидазол-4-ил)пропионовая к-та His, Гис
- 13. 2. Номенклатура 2.1. Тривиальная номенклатура в основном используется для широко распространённых аминокислот. 2.2. Рациональная 2.3. IUPAC
- 14. 3. Изомерия 3.1. Структурная изомерия 25.04.2012 лейцин изолейцин аланин β-аланин
- 15. 3.2. Пространственная изомерия 25.04.2012 D-аминокислота L-аминокислота Формулы Фишера Формулы с клиновидными связями
- 16. 4. Физические свойства Аминокислоты, как правило, являются бесцветными кристаллическими соединениями. Большинство из них умеренно растворимы в
- 17. 5. Биологические свойства Из остатков аминокислот построены такие важные соединения как белки, которые участвуют практически во
- 18. うま味 Умами - “мясной вкус” 25.04.2012 Глутаминовая кислота (E620) и её соли (глутамат натрия Е621, глутамат
- 19. Глутаминовая кислота и её соли безопасны J Nutr. 2000 Apr;130(4S Suppl):1049S-52S. The safety evaluation of monosodium
- 20. 6. Химические свойства 6.1. Образование солей 25.04.2012 аланин 2-аминопропионат натрия аланин гидрохлорид аланина
- 21. 25.04.2012 Аминогруппа нейтрализует карбоксильную группу, поэтому АК в твёрдом виде и в растворе при pH =
- 22. 25.04.2012 Сильнокислая среда Почти нейтральная Сильнощелочная среда Для моноаминомонокарбоновых кислот pI ≈ 5-6 pI моноаминодикарбоновых кислот
- 23. 6.2. Реакции по аминогруппе 6.2.1. Ацилирование 25.04.2012 аланин ацетилхлорид N-ацетилаланин
- 24. 25.04.2012 БОК-аминокислота Трет-бутокси-карбоксазид используется для защиты аминогруппы в пептидном синтезе
- 25. 6.2.2. Арилирование 25.04.2012 N-(2,4-динитрофенил)аланин Реакция используется для определения N-концевой аминокислоты в пептидах и белках по Сэнджеру
- 26. 6.2.3. Взаимодействие с карбонильными соединениями 25.04.2012
- 27. 25.04.2012 Данная реакция используется в формольном титровании по Сёренсену: метилольные производные являются гораздо более сильными кислотами
- 28. 6.2.4. Взаимодействие с азотистой кислотой 25.04.2012 α -аминокислота α-гидроксикислота Реакция с азотистой кислотой используется определения аминокислот
- 29. 6.2. Реакции по карбоксильной группе 25.04.2012 Реакция используется для защиты карбоксильной группы в синтезе пептидов
- 30. 6.2.2. Декарбоксилирование 25.04.2012
- 31. 25.04.2012
- 32. Взаимодействие по обеим группам – образование хелатных комплексов 25.04.2012
- 33. 6.3. Поведение аминокислот при нагревании α-аминокислоты при нагревании дают дикетопиперазины (диоксопиперазины): 25.04.2012 аланин 2,5-диоксо-3,6-диметилпиперазин
- 34. β-аминокислоты при нагревании отщепляют воду (реакция элиминирования), образуя ненасыщенные кислоты: 25.04.2012 β-аминопропионовая пропеновая (акриловая) кислота кислота
- 35. γ- и δ-аминокислоты при нагревании отщепляют воду, циклизуются с образованием лактамов – циклических амидов: 25.04.2012 γ-аминомасляная
- 36. 25.04.2012 δ-аминовалериановая кислота δ-валеролактам
- 37. 7. Получение аминокислот 7.1. Выделение из белков и пептидов Белки гидролизуют в присутствии кислот (6 М
- 38. 7.3. Пребиотический (абиогенный) синтез аминокислот CH4, NH3, H2, H2O, HCN, H2S, CH2O УФ-излучение, электрический разряд, радиация
- 39. Proc Natl Acad Sci U S A. 2011 Apr 5;108(14):5526-31. Epub 2011 Mar 21. Primordial synthesis
- 40. 25.04.2012 http://ru.wikipedia.org/wiki/Миллер,_стенли_ллойд
- 41. Orig Life Evol Biosph. 2011 Apr 12. [Epub ahead of print] Catalytic effects of Murchison Material:
- 42. 7.4. Химические синтезы аминокислот 7.4.1. Аммонолиз галогенкарбоновых кислот 7.4.2. Синтез Штреккера 25.04.2012
- 43. ПЕПТИДЫ И БЕЛКИ Пептиды и белки - продукты поликонденсации аминокислот. Мr пептидов меньше 10000, у белков
- 44. ПЕПТИДЫ И БЕЛКИ Пептиды (πεπτος - сваренный) – первоначально - продукты неполного гидролиза белков Белки –
- 45. ПЕПТИДЫ И БЕЛКИ Остатки АК связаны пептидной связью: Пептидная связь дипептид
- 46. ПЕПТИДЫ И БЕЛКИ Треонин Фенилаланин Тирозин Трипептид- Thr-Phe-Tyr Треонил-Фенилаланил-Тирозин
- 47. Глицил-лизил-тирозин
- 48. Глутамил-аланил-цистеин
- 49. ПЕПТИДНАЯ СВЯЗЬ Пространственное строение 1. Все атомы находятся в одной плоскости 2. Почти всегда атомы водорода
- 50. ПЕПТИДНАЯ СВЯЗЬ Электронное строение НЭП на азоте сопряжена с карбонильной группой (С=О), поэтому связь C—N имеет
- 51. ПЕПТИДЫ И БЕЛКИ Как и аминокислоты, пептиды и белки являются амфотерными соединениями, содержащими и кислотные группы
- 52. АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ ПЕПТИДОВ МЕТОД ЭДМАНА Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК с N-конца и определяют
- 53. МЕТОД ЭДМАНА Фенилтиогидантоиновое производное N-концевой АК Пептид укороченный на 1 АК
- 54. СИНТЕЗ ПЕПТИДОВ И БЕЛКОВ Защита аминогруппы (БОК-защита) Защита карбоксильной группы (образование сложных эфиров) Образование пептидной связи
- 55. Снятие защиты Образование пептидной связи и снятие защиты
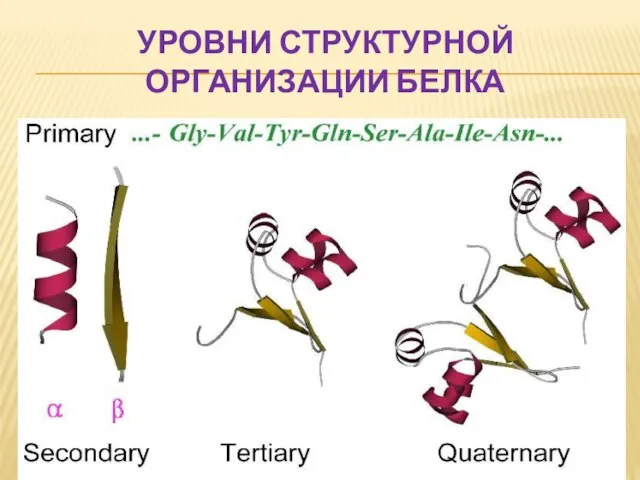
- 56. УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА первичная структура – аминокислотная последовательность вторичная структура – локальные высокоупорядоченные конформации белковой
- 57. УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
- 58. ПЕРВИЧНАЯ СТРУКТУРА первичная структура –последовательность аминокислотных остатков в молекуле белка или пептида. NH2-Tyr-Pro-Lys-Gly-Phe-Tyr-Lys-COOH Первичная структура определяет
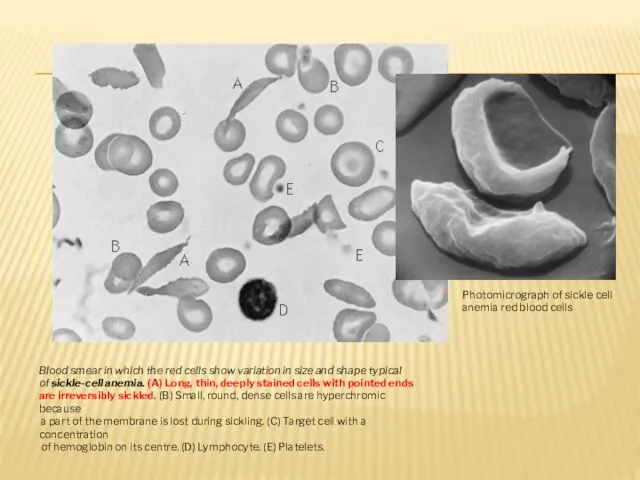
- 59. Blood smear in which the red cells show variation in size and shape typical of sickle-cell
- 60. Количество возможных первичных структур ОГРОМНО! 20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих из 150 остатков
- 61. ВТОРИЧНАЯ СТРУКТУРА Вторичная структура- локальные высокоупорядоченные конформации белковой цепи – спирали и складчатые слои.
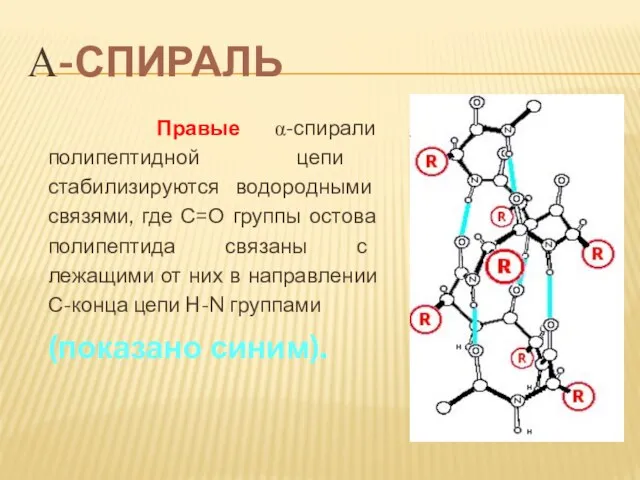
- 62. Α-СПИРАЛЬ Правые α-спирали полипептидной цепи стабилизируются водородными связями, где С=О группы остова полипептида связаны с лежащими
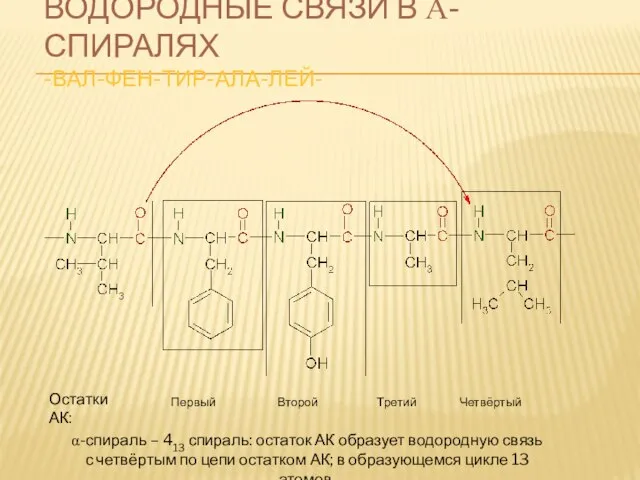
- 63. ВОДОРОДНЫЕ СВЯЗИ В Α-СПИРАЛЯХ -ВАЛ-ФЕН-ТИР-АЛА-ЛЕЙ- Первый Второй Третий Четвёртый Остатки АК: α-спираль – 413 спираль: остаток
- 64. СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ β-структура образуется из нескольких полиипептидных цепей, связанных водородными связями. Она существует в виде
- 65. СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ ВОДОРОДНЫЕ СВЯЗИ Тре-Ала-Гли-Сер-Асп Вал-Фен-Тир-Ала-Лей
- 66. ОБОЗНАЧЕНИЕ ВТОРИЧНОЙ СТРУКТУРЫ Укладка α-спиралей и β-структуры с образованием глобулы
- 67. ТРЕТИЧНАЯ СТРУКТУРА третичная структура – форма белковой молекулы; трёхмерная структура белка. Укладка нерегулярных областей и α
- 68. ВЗАИМОДЕЙСТВИЯ ОСТАТКОВ АК В ТРЕТИЧНОЙ СТРУКТУРЕ Остатки АК в белковой глобуле взаимодействуют за счёт: Гидрофобных взаимодействий
- 70. ДИСУЛЬФИДНЫЕ СВЯЗИ В ЦИСТИНЕ
- 71. ДИСУЛЬФИДНЫЕ СВЯЗИ Структура лизоцима. Дисульфидные связи показаны пунктиром
- 72. ИОННЫЕ СВЯЗИ Асп Лиз
- 73. ТРЕТИЧНАЯ СТРУКТУРА Сериновая протеаза - трипсин Родопсин
- 74. ТРЕТИЧНАЯ СТРУКТУРА ацилтрансфераза пиктатлиаза С
- 75. ТРЕТИЧНАЯ СТРУКТУРА α и β цепи гемоглобина лошади
- 76. ТРЕТИЧНАЯ СТРУКТУРА Порин
- 77. Фолдинг – сворачивание белковой цепи с образованием нативной структуры.
- 78. Неправильный фолдинг может привести к различным болезням, например к губчатым энцефалопатиям: Синдром Крейцфельда-Якоба Новый вариант Крейцфельда-Якоба
- 79. ДОМЕНЫ Домены – глобулярные области в пределах одной белковой молекулы Домены соединены шарнирным участком Доменная структура
- 80. ЧЕТВЕРТИЧНАЯ СТРУКТУРА Четвертичная структура- агрегат нескольких белковых молекул образующих одну структуру Взаимодействия: ионные, водородные, гидрофобные, ковалентные
- 81. ДИСУЛЬФИДНЫЕ СВЯЗИ Дисульфидные связи в иммуноглобулине G
- 82. ЧЕТВЕРТИЧНАЯ СТРУКТУРА Пример четвертичной структуры - вирус табачной мозаики: 2130 одинаковых молекул белка расположены вокруг РНК
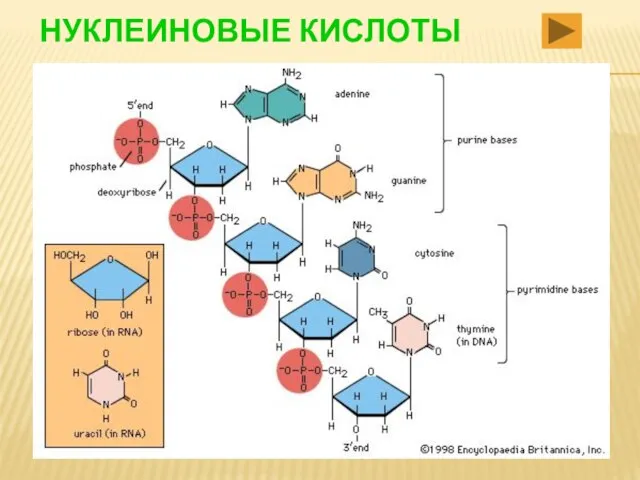
- 83. НУКЛЕИНОВЫЕ КИСЛОТЫ НК- это полинуклеотиды, т.е. биополимеры, мономерами которых являются нуклеотиды. Нуклеотиды-фосфорные эфиры нуклеозидов Нуклеозиды –
- 84. НУКЛЕИНОВЫЕ КИСЛОТЫ
- 85. НУКЛЕИНОВЫЕ КИСЛОТЫ РНК ДНК нуклеотиды нуклеозиды пентоза Нукл. Осн. Гидролиз
- 86. НУКЛЕИНОВЫЕ ОСНОВАНИЯ ПУРИНОВЫЕ ОСНОВАНИЯ Аденин Гуанин лактам лактим
- 87. НУКЛЕИНОВЫЕ ОСНОВАНИЯ ПИРИМИДИНОВЫЕ ОСНОВАНИЯ Урацил Тимин Цитозин
- 88. НУКЛЕОЗИДЫ Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой) Уридин
- 89. НУКЛЕОЗИДЫ Надо иметь ввиду, что можно и иногда надо писать нуклеиновое основание развёрнутым на 180 градусов.
- 90. НУКЛЕОЗИДЫ Гликозидная связь
- 91. НУКЛЕОЗИДЫ Цитидин Дезоксицитидин
- 92. НУКЛЕОЗИДЫ Тимидин
- 93. НУКЛЕОЗИДЫ Аденозин Дезоксиаденозин
- 94. НУКЛЕОЗИДЫ Аденозин Дезоксиаденозин Аденозин и дезоксиаденозин можно изобразить и так:
- 95. НУКЛЕОЗИДЫ Гуанозин Дезоксигуанозин
- 96. НУКЛЕОЗИДЫ Гуанозин Дезоксигуанозин Гуанозин и дезоксигуанозин можно изобразить и так:
- 97. ГИДРОЛИЗ НУКЛЕОЗИДОВ аденозин рибоза аденин
- 98. НУКЛЕОТИДЫ Нуклеотиды - фосфорные эфиры нуклеозидов гликозидная связь сложноэфирная связь
- 99. НУКЛЕОТИДЫ Аденозин-5’-фосфат Аденозинмонофосфат (АМФ) 5’-адениловая кислота Гуанозин-5’-фосфат Гуанозинмонофосфат (ГМФ) 5’-гуаниловая кислота
- 100. НУКЛЕОТИДЫ Уридин-5’-фосфат Уридинмонофосфат (УМФ) 5’-уридиловая кислота Цитидин-5’-фосфат Цитидинмонофосфат (ЦМФ) 5’-цитидиловая кислота
- 101. НУКЛЕОТИДЫ
- 102. НУКЛЕОТИДЫ Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам. У других нуклеотидов в сокращённом названии указывают
- 103. НУКЛЕОТИДЫ цAMФ
- 104. Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль вторичного посредника некоторых гормонов (глюкагона
- 105. ГИДРОЛИЗ НУКЛЕОТИДОВ АМФ аденозин рибоза аденин
- 106. НУКЛЕОТИДЫ - КОФЕРМЕНТЫ АМФ АДФ АТФ Ангидридные связи Сложноэфирная связь АТФ – универсальный макроэрг
- 107. Образование АТФ происходит 1. В результате субстратного фосфорилирования (гликолиз, одна реакция ЦТК) 2. Окислительное фосфорилирование (в
- 108. АТФ КАК КОФЕРМЕНТ АТФ АДФ АТФ АДФ АТФ АДФ ГМФ ГДФ ГТФ ГМФ гуанозин
- 109. ОБРАЗОВАНИЕ АМИНОАЦИЛАДЕНИЛАТА АТФ аминокислота аминоациладенилат Аминоацил-тРНК (в синтез белка)
- 110. НИКОТИНАМИДДИНУКЛЕОТИД (НАД+) КАК КОФЕРМЕНТ НАД+ НАДН
- 111. ПЕРВИЧНАЯ СТРУКТУРА НК Первичная структура НК - последовательность нуклеотидов 5’- конец ТГАЦТААГТАЦЦ 3’-конец (ф-конец) (OH-конец)
- 112. ПЕРВИЧНАЯ СТРУКТУРА ДНК Т Г А
- 113. ПЕРВИЧНАЯ СТРУКТУРА РНК У Г А
- 114. ВТОРИЧНАЯ СТРУКТУРА Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953) Две цепи антипараллельны друг
- 115. ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ КОМПЛЕМЕНТАРНЫМИ ОСНОВАНИЯМИ В ДНК Г Ц А Т
- 117. Скачать презентацию

Слайд 31.2. По количеству карбокси- и аминогрупп
Моноаминомонокарбоновые кислоты (глицин, аланин, валин, лейцин, изолейцин,
1.2. По количеству карбокси- и аминогрупп
Моноаминомонокарбоновые кислоты (глицин, аланин, валин, лейцин, изолейцин,

Слайд 41.3 Классификация по встречаемости в белках
25.04.2012
20 классических протеиногенных аминокислот, информация о
1.3 Классификация по встречаемости в белках
25.04.2012
20 классических протеиногенных аминокислот, информация о

Слайд 51.4. По пищевой ценности для человека
Аминокислоты делятся на
заменимые и незаменимые.
К незаменимым
1.4. По пищевой ценности для человека
Аминокислоты делятся на
заменимые и незаменимые.
К незаменимым

Слайд 6АЛИФАТИЧЕСКИЕ АК
АЛИФАТИЧЕСКИЕ АК
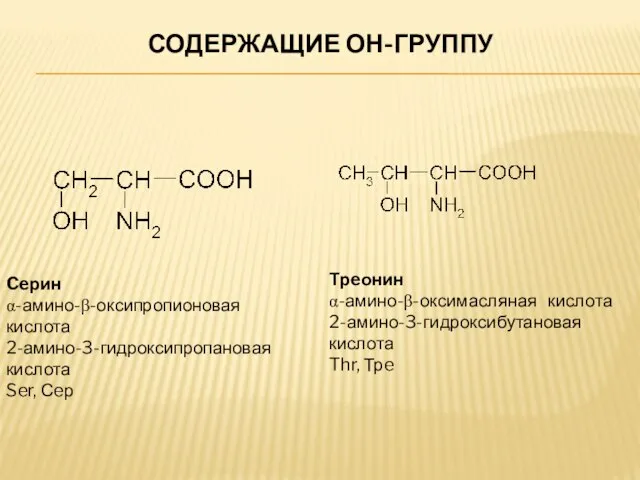
Слайд 7СОДЕРЖАЩИЕ ОН-ГРУППУ
Ceрин
α-амино-β-оксипропионовая кислота
2-амино-3-гидроксипропановая кислота
Ser, Сeр
Трeонин
α-амино-β-оксимасляная кислота
2-амино-3-гидроксибутановая кислота
Thr, Трe
СОДЕРЖАЩИЕ ОН-ГРУППУ
Ceрин
α-амино-β-оксипропионовая кислота
2-амино-3-гидроксипропановая кислота
Ser, Сeр
Трeонин
α-амино-β-оксимасляная кислота
2-амино-3-гидроксибутановая кислота
Thr, Трe
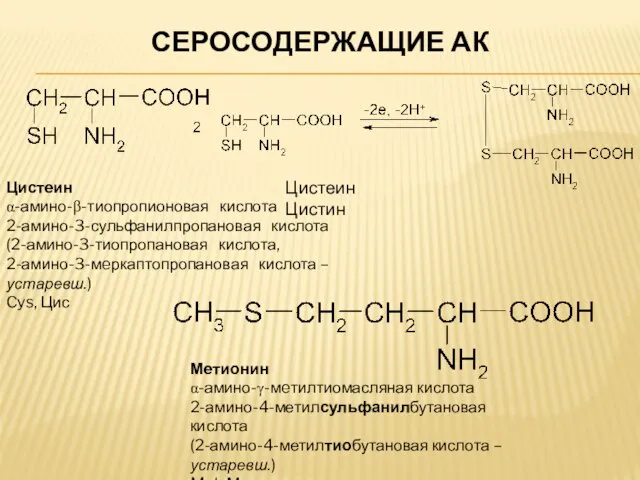
Слайд 8СЕРОСОДЕРЖАЩИЕ АК
Цистеин Цистин
Мeтионин
α-амино-γ-мeтилтиомасляная кислота
2-амино-4-метилсульфанилбутановая кислота
(2-амино-4-метилтиобутановая кислота – устаревш.)
Met, Мет.
Цистeин
α-амино-β-тиопропионовая кислота
2-амино-3-сульфанилпропановая кислота
(2-амино-3-тиопропановая кислота,
СЕРОСОДЕРЖАЩИЕ АК
Цистеин Цистин
Мeтионин
α-амино-γ-мeтилтиомасляная кислота
2-амино-4-метилсульфанилбутановая кислота
(2-амино-4-метилтиобутановая кислота – устаревш.)
Met, Мет.
Цистeин
α-амино-β-тиопропионовая кислота
2-амино-3-сульфанилпропановая кислота
(2-амино-3-тиопропановая кислота,
Слайд 9МОНОАМИНОДИКАРБОНОВЫЕ КИСЛОТЫ И ИХ АМИДЫ
Аспарагиновая кислота
Аминоянтарная кислота
Аминобутандиовая кислота
Asp, Асп
Глутаминовая кислота
α-aминоглутаровая кислота
2-аминопентандиовая кислота
Glu,
МОНОАМИНОДИКАРБОНОВЫЕ КИСЛОТЫ И ИХ АМИДЫ
Аспарагиновая кислота
Аминоянтарная кислота
Аминобутандиовая кислота
Asp, Асп
Глутаминовая кислота
α-aминоглутаровая кислота
2-аминопентандиовая кислота
Glu,

Слайд 10СОДЕРЖАЩИЕ АМИНОГРУППУ
Лизин
α,ε-диаминокапроновая кислота
2,6-диаминогексановая кислота
Lys, Лиз
Аргинин
α-амино-δ-гуанидилвалериановая кислота
2-амино-5-[амино(имино)метил]аминопентановая к-та
Arg, Арг
СОДЕРЖАЩИЕ АМИНОГРУППУ
Лизин
α,ε-диаминокапроновая кислота
2,6-диаминогексановая кислота
Lys, Лиз
Аргинин
α-амино-δ-гуанидилвалериановая кислота
2-амино-5-[амино(имино)метил]аминопентановая к-та
Arg, Арг
![СОДЕРЖАЩИЕ АМИНОГРУППУ Лизин α,ε-диаминокапроновая кислота 2,6-диаминогексановая кислота Lys, Лиз Аргинин α-амино-δ-гуанидилвалериановая кислота 2-амино-5-[амино(имино)метил]аминопентановая к-та Arg, Арг](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/426661/slide-9.jpg)
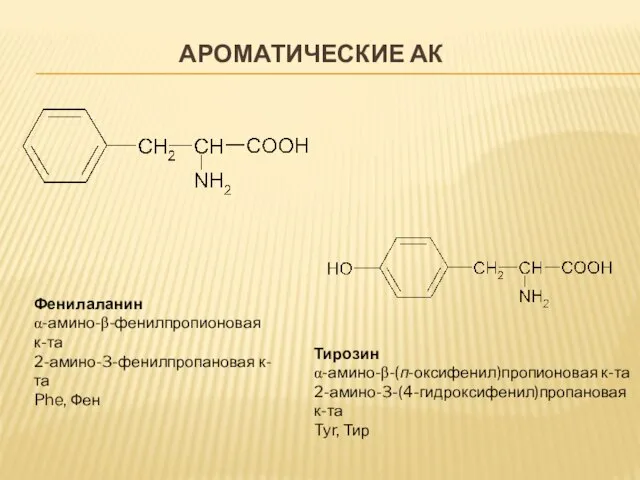
Слайд 11АРОМАТИЧЕСКИЕ АК
Фенилаланин
α-амино-β-фенилпропионовая к-та
2-амино-3-фенилпропановая к-та
Phe, Фен
Тирозин
α-амино-β-(п-оксифенил)пропионовая к-та
2-амино-3-(4-гидроксифенил)пропановая к-та
Tyr, Тир
АРОМАТИЧЕСКИЕ АК
Фенилаланин
α-амино-β-фенилпропионовая к-та
2-амино-3-фенилпропановая к-та
Phe, Фен
Тирозин
α-амино-β-(п-оксифенил)пропионовая к-та
2-амино-3-(4-гидроксифенил)пропановая к-та
Tyr, Тир
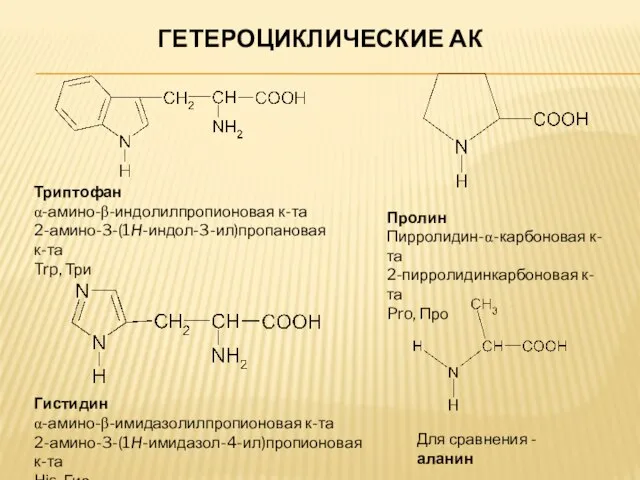
Слайд 12ГЕТЕРОЦИКЛИЧЕСКИЕ АК
Триптофан
α-амино-β-индолилпропионовая к-та
2-амино-3-(1H-индол-3-ил)пропановая к-та
Trp, Три
Гистидин
α-амино-β-имидазолилпропионовая к-та
2-амино-3-(1H-имидазол-4-ил)пропионовая к-та
His, Гис
Пролин
Пирролидин-α-карбоновая к-та
2-пирролидинкарбоновая к-та
Pro, Про
Для сравнения
ГЕТЕРОЦИКЛИЧЕСКИЕ АК
Триптофан
α-амино-β-индолилпропионовая к-та
2-амино-3-(1H-индол-3-ил)пропановая к-та
Trp, Три
Гистидин
α-амино-β-имидазолилпропионовая к-та
2-амино-3-(1H-имидазол-4-ил)пропионовая к-та
His, Гис
Пролин
Пирролидин-α-карбоновая к-та
2-пирролидинкарбоновая к-та
Pro, Про
Для сравнения
Слайд 132. Номенклатура
2.1. Тривиальная номенклатура в основном используется для широко распространённых аминокислот.
2.2. Рациональная
2. Номенклатура
2.1. Тривиальная номенклатура в основном используется для широко распространённых аминокислот.
2.2. Рациональная

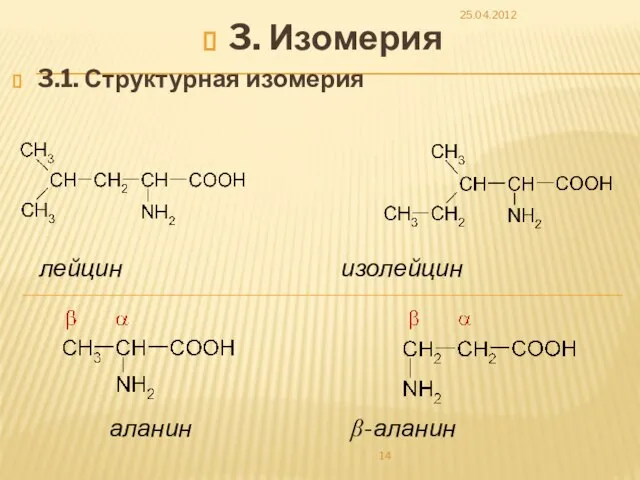
Слайд 143. Изомерия
3.1. Структурная изомерия
25.04.2012
лейцин изолейцин
аланин β-аланин
3. Изомерия
3.1. Структурная изомерия
25.04.2012
лейцин изолейцин
аланин β-аланин
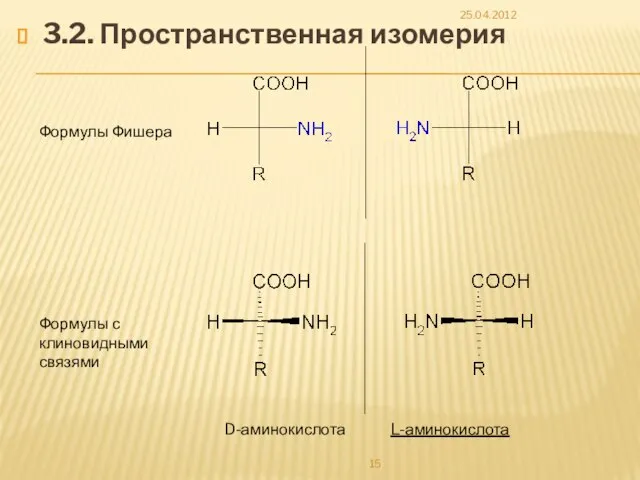
Слайд 153.2. Пространственная изомерия
25.04.2012
D-аминокислота L-аминокислота
Формулы Фишера
Формулы с
клиновидными
связями
3.2. Пространственная изомерия
25.04.2012
D-аминокислота L-аминокислота
Формулы Фишера
Формулы с
клиновидными
связями
Слайд 164. Физические свойства
Аминокислоты, как правило, являются бесцветными кристаллическими соединениями. Большинство из них
4. Физические свойства
Аминокислоты, как правило, являются бесцветными кристаллическими соединениями. Большинство из них

Слайд 175. Биологические свойства
Из остатков аминокислот построены такие важные соединения как белки, которые
5. Биологические свойства
Из остатков аминокислот построены такие важные соединения как белки, которые

Слайд 18うま味
Умами - “мясной вкус”
25.04.2012
Глутаминовая кислота (E620) и её соли (глутамат натрия Е621,
うま味
Умами - “мясной вкус”
25.04.2012
Глутаминовая кислота (E620) и её соли (глутамат натрия Е621,

Слайд 19Глутаминовая кислота и её соли безопасны
J Nutr. 2000 Apr;130(4S Suppl):1049S-52S.
The safety evaluation
Глутаминовая кислота и её соли безопасны
J Nutr. 2000 Apr;130(4S Suppl):1049S-52S.
The safety evaluation

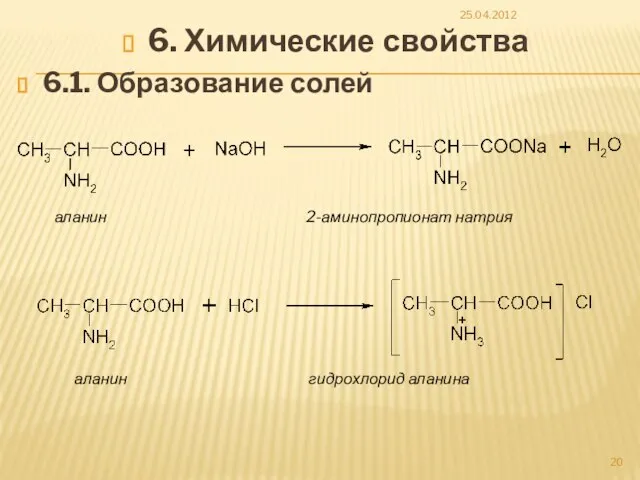
Слайд 206. Химические свойства
6.1. Образование солей
25.04.2012
аланин 2-аминопропионат натрия
аланин гидрохлорид аланина
6. Химические свойства
6.1. Образование солей
25.04.2012
аланин 2-аминопропионат натрия
аланин гидрохлорид аланина
Слайд 2125.04.2012
Аминогруппа нейтрализует карбоксильную группу,
поэтому АК в твёрдом виде и в растворе
25.04.2012
Аминогруппа нейтрализует карбоксильную группу,
поэтому АК в твёрдом виде и в растворе

Слайд 2225.04.2012
Сильнокислая среда Почти нейтральная Сильнощелочная среда
Для моноаминомонокарбоновых кислот pI ≈ 5-6
pI моноаминодикарбоновых
25.04.2012
Сильнокислая среда Почти нейтральная Сильнощелочная среда
Для моноаминомонокарбоновых кислот pI ≈ 5-6
pI моноаминодикарбоновых

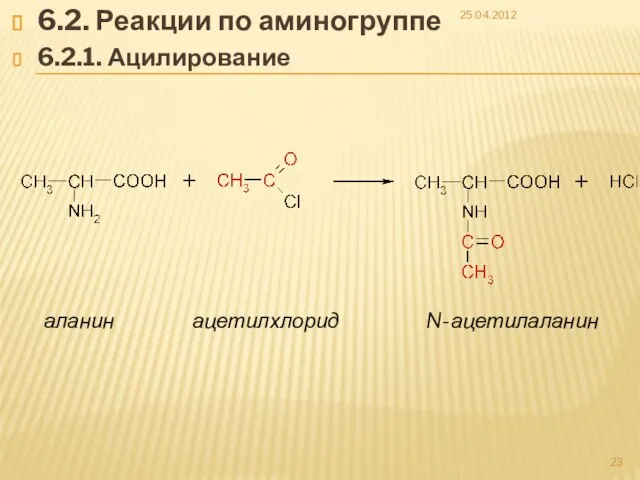
Слайд 236.2. Реакции по аминогруппе
6.2.1. Ацилирование
25.04.2012
аланин ацетилхлорид N-ацетилаланин
6.2. Реакции по аминогруппе
6.2.1. Ацилирование
25.04.2012
аланин ацетилхлорид N-ацетилаланин
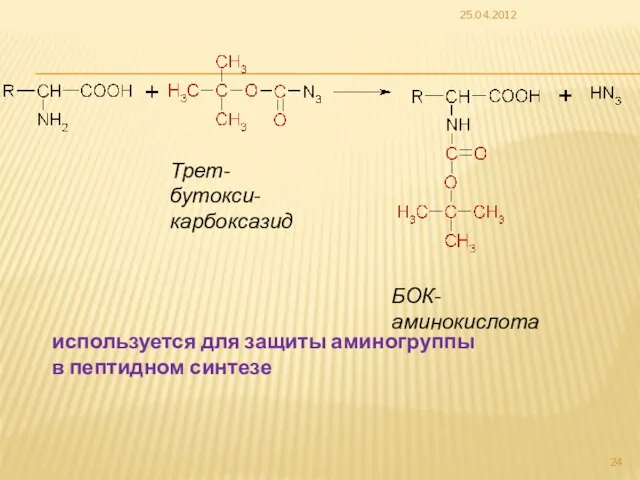
Слайд 2425.04.2012
БОК-аминокислота
Трет-бутокси-карбоксазид
используется для защиты аминогруппы
в пептидном синтезе
25.04.2012
БОК-аминокислота
Трет-бутокси-карбоксазид
используется для защиты аминогруппы
в пептидном синтезе
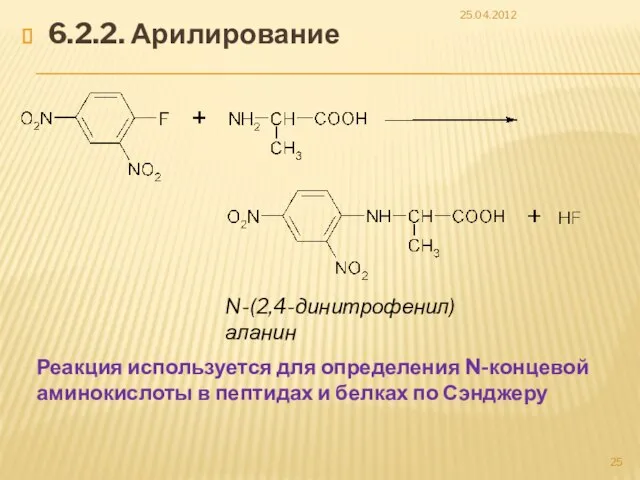
Слайд 256.2.2. Арилирование
25.04.2012
N-(2,4-динитрофенил)аланин
Реакция используется для определения N-концевой аминокислоты в пептидах и белках по
6.2.2. Арилирование
25.04.2012
N-(2,4-динитрофенил)аланин
Реакция используется для определения N-концевой аминокислоты в пептидах и белках по
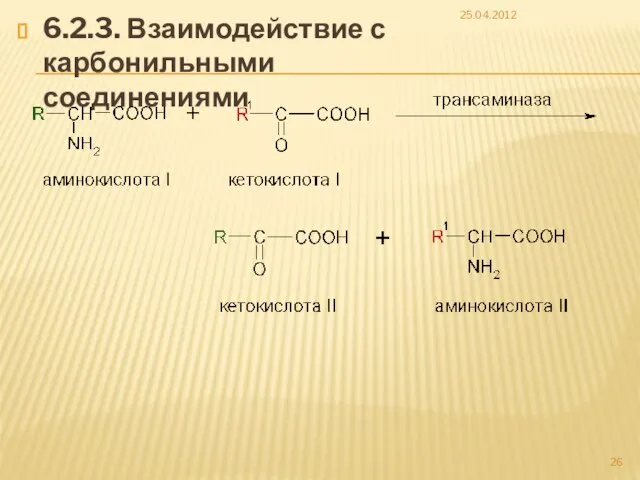
Слайд 266.2.3. Взаимодействие с карбонильными соединениями
25.04.2012
6.2.3. Взаимодействие с карбонильными соединениями
25.04.2012
Слайд 2725.04.2012
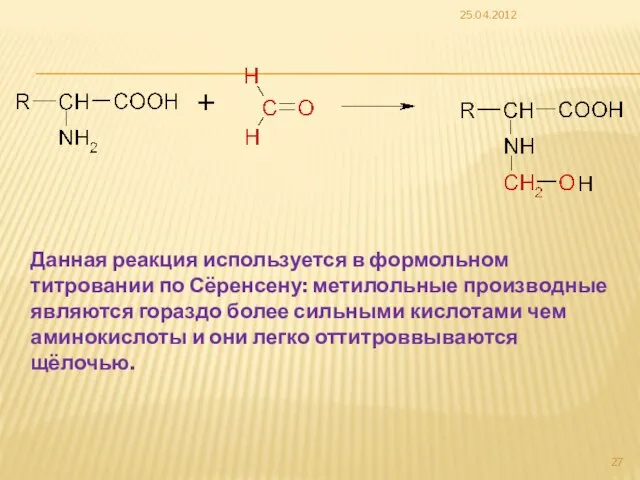
Данная реакция используется в формольном титровании по Сёренсену: метилольные производные являются гораздо
25.04.2012
Данная реакция используется в формольном титровании по Сёренсену: метилольные производные являются гораздо
Слайд 286.2.4. Взаимодействие с азотистой кислотой
25.04.2012
α -аминокислота α-гидроксикислота
Реакция с азотистой кислотой используется определения
6.2.4. Взаимодействие с азотистой кислотой
25.04.2012
α -аминокислота α-гидроксикислота
Реакция с азотистой кислотой используется определения

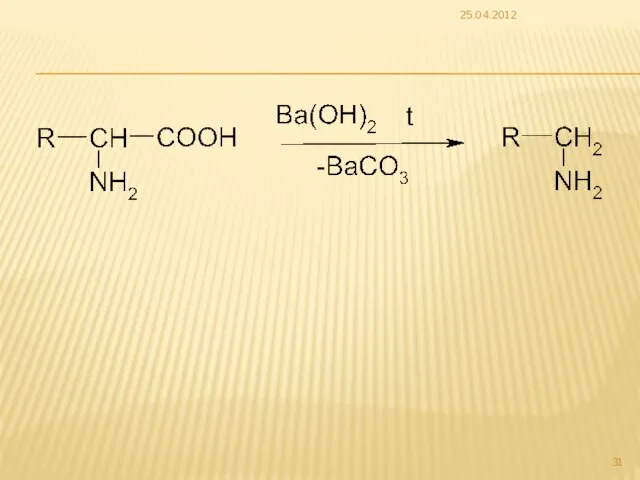
Слайд 296.2. Реакции по карбоксильной группе
25.04.2012
Реакция используется для защиты карбоксильной группы в синтезе
6.2. Реакции по карбоксильной группе
25.04.2012
Реакция используется для защиты карбоксильной группы в синтезе

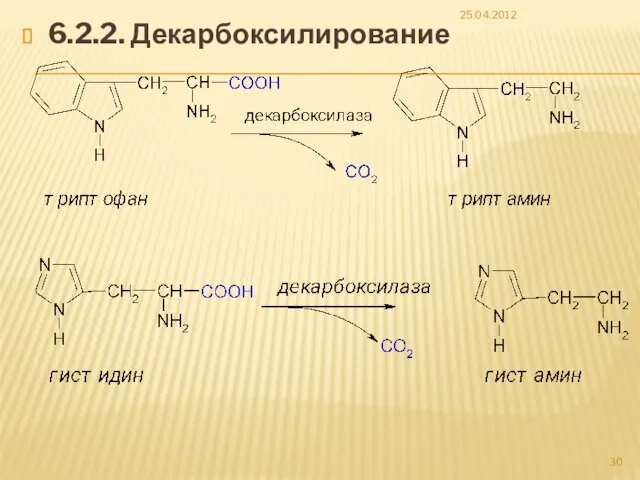
Слайд 306.2.2. Декарбоксилирование
25.04.2012
6.2.2. Декарбоксилирование
25.04.2012
Слайд 3125.04.2012
25.04.2012
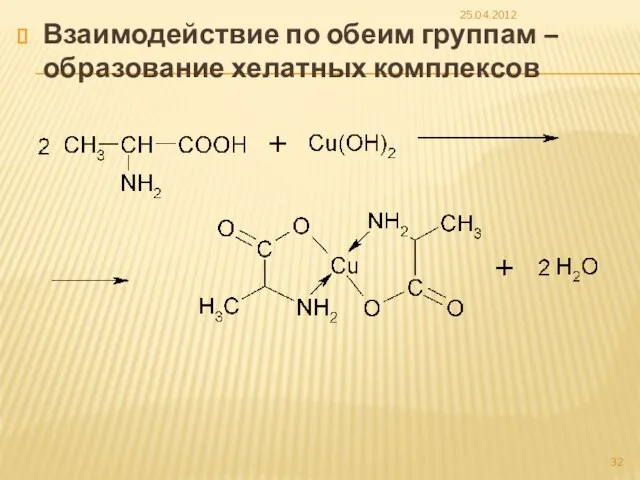
Слайд 32Взаимодействие по обеим группам – образование хелатных комплексов
25.04.2012
Взаимодействие по обеим группам – образование хелатных комплексов
25.04.2012
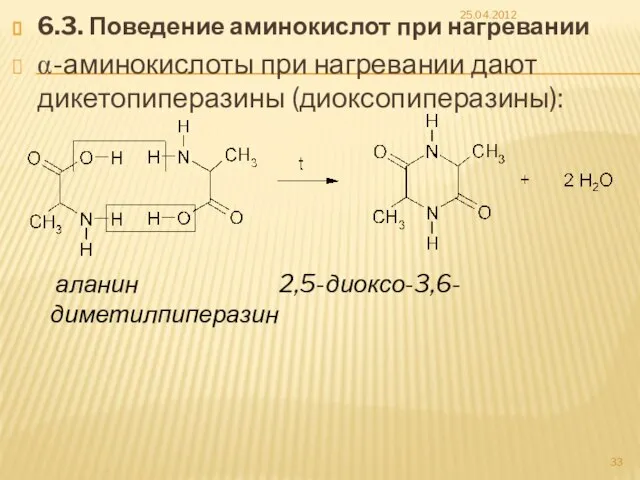
Слайд 336.3. Поведение аминокислот при нагревании
α-аминокислоты при нагревании дают дикетопиперазины (диоксопиперазины):
25.04.2012
аланин 2,5-диоксо-3,6-диметилпиперазин
6.3. Поведение аминокислот при нагревании
α-аминокислоты при нагревании дают дикетопиперазины (диоксопиперазины):
25.04.2012
аланин 2,5-диоксо-3,6-диметилпиперазин
Слайд 34β-аминокислоты при нагревании отщепляют воду (реакция элиминирования), образуя ненасыщенные кислоты:
25.04.2012
β-аминопропионовая пропеновая (акриловая)
β-аминокислоты при нагревании отщепляют воду (реакция элиминирования), образуя ненасыщенные кислоты:
25.04.2012
β-аминопропионовая пропеновая (акриловая)

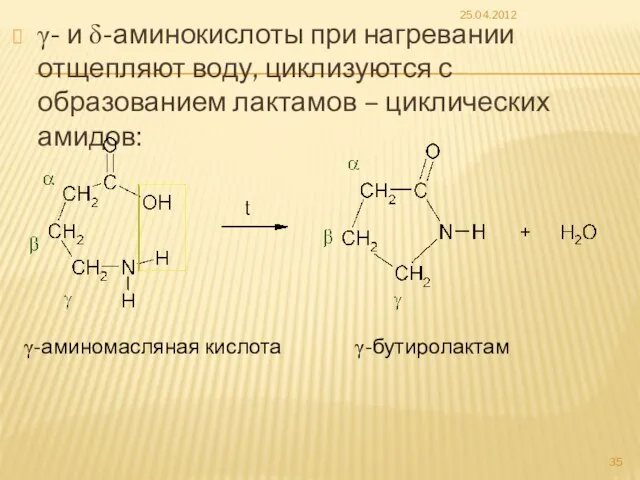
Слайд 35γ- и δ-аминокислоты при нагревании отщепляют воду, циклизуются с образованием лактамов –
γ- и δ-аминокислоты при нагревании отщепляют воду, циклизуются с образованием лактамов –
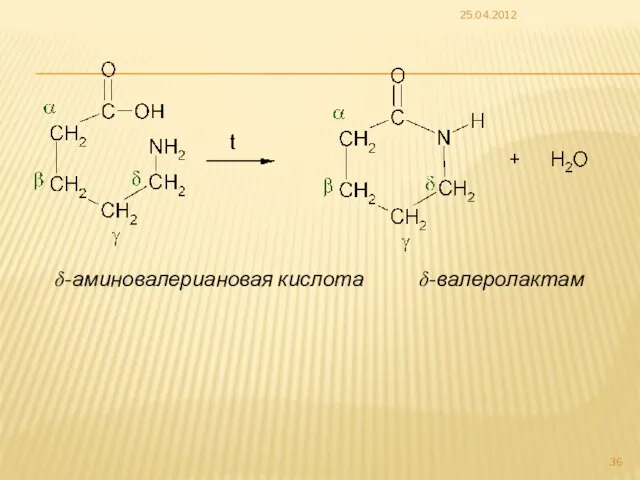
Слайд 3625.04.2012
δ-аминовалериановая кислота δ-валеролактам
25.04.2012
δ-аминовалериановая кислота δ-валеролактам
Слайд 377. Получение аминокислот
7.1. Выделение из белков и пептидов
Белки гидролизуют в присутствии кислот
7. Получение аминокислот
7.1. Выделение из белков и пептидов
Белки гидролизуют в присутствии кислот

Слайд 387.3. Пребиотический (абиогенный) синтез аминокислот
CH4, NH3, H2, H2O, HCN, H2S, CH2O
УФ-излучение, электрический
7.3. Пребиотический (абиогенный) синтез аминокислот
CH4, NH3, H2, H2O, HCN, H2S, CH2O
УФ-излучение, электрический

Слайд 39Proc Natl Acad Sci U S A. 2011 Apr 5;108(14):5526-31. Epub 2011
Proc Natl Acad Sci U S A. 2011 Apr 5;108(14):5526-31. Epub 2011

Слайд 4025.04.2012
http://ru.wikipedia.org/wiki/Миллер,_стенли_ллойд
25.04.2012
http://ru.wikipedia.org/wiki/Миллер,_стенли_ллойд
Слайд 41Orig Life Evol Biosph. 2011 Apr 12. [Epub ahead of print]
Catalytic effects
Orig Life Evol Biosph. 2011 Apr 12. [Epub ahead of print]
Catalytic effects
![Orig Life Evol Biosph. 2011 Apr 12. [Epub ahead of print] Catalytic](/_ipx/f_webp&q_80&fit_contain&s_1440x1080/imagesDir/jpg/426661/slide-40.jpg)
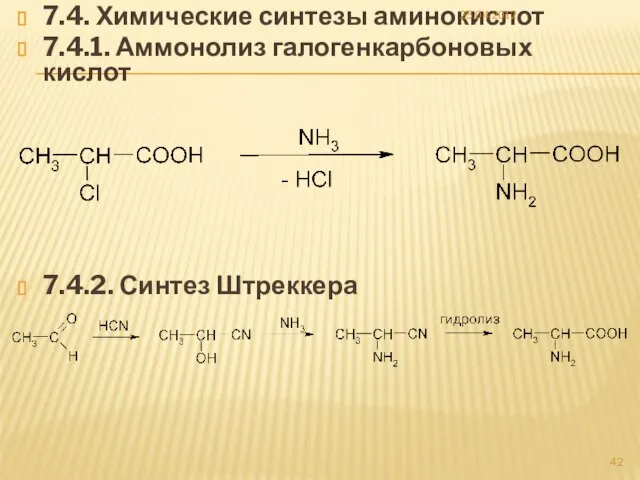
Слайд 427.4. Химические синтезы аминокислот
7.4.1. Аммонолиз галогенкарбоновых кислот
7.4.2. Синтез Штреккера
25.04.2012
7.4. Химические синтезы аминокислот
7.4.1. Аммонолиз галогенкарбоновых кислот
7.4.2. Синтез Штреккера
25.04.2012
Слайд 43ПЕПТИДЫ И БЕЛКИ
Пептиды и белки - продукты поликонденсации аминокислот.
Мr пептидов меньше 10000,
ПЕПТИДЫ И БЕЛКИ
Пептиды и белки - продукты поликонденсации аминокислот.
Мr пептидов меньше 10000,

Слайд 44ПЕПТИДЫ И БЕЛКИ
Пептиды (πεπτος - сваренный) – первоначально - продукты неполного гидролиза
ПЕПТИДЫ И БЕЛКИ
Пептиды (πεπτος - сваренный) – первоначально - продукты неполного гидролиза

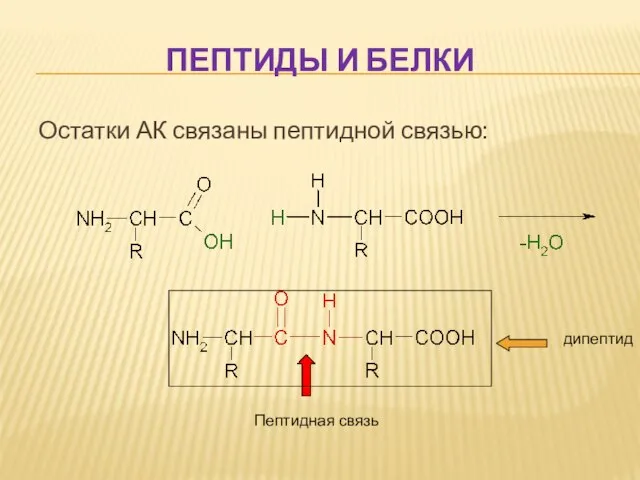
Слайд 45ПЕПТИДЫ И БЕЛКИ
Остатки АК связаны пептидной связью:
Пептидная связь
дипептид
ПЕПТИДЫ И БЕЛКИ
Остатки АК связаны пептидной связью:
Пептидная связь
дипептид
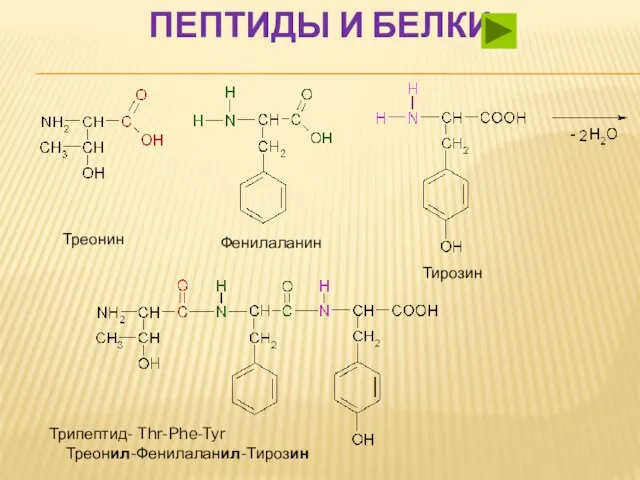
Слайд 46ПЕПТИДЫ И БЕЛКИ
Треонин
Фенилаланин
Тирозин
Трипептид- Thr-Phe-Tyr
Треонил-Фенилаланил-Тирозин
ПЕПТИДЫ И БЕЛКИ
Треонин
Фенилаланин
Тирозин
Трипептид- Thr-Phe-Tyr
Треонил-Фенилаланил-Тирозин
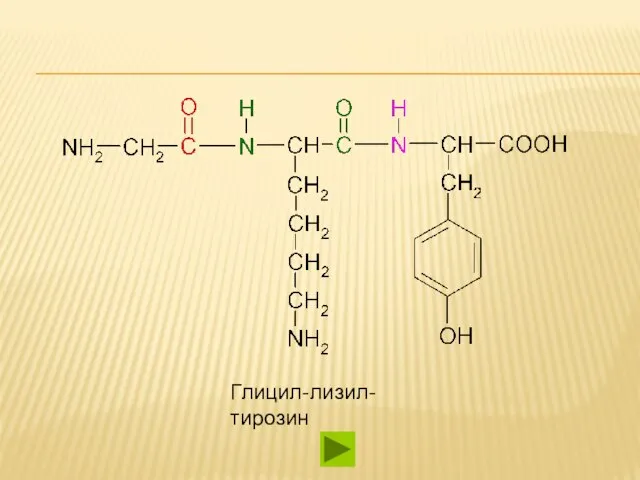
Слайд 47Глицил-лизил-тирозин
Глицил-лизил-тирозин
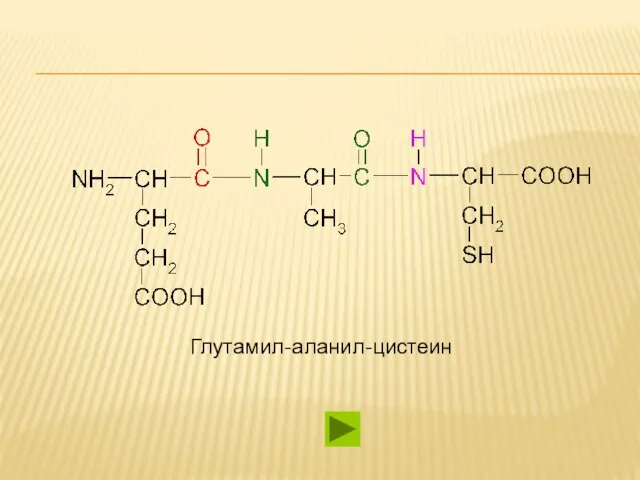
Слайд 48Глутамил-аланил-цистеин
Глутамил-аланил-цистеин
Слайд 49ПЕПТИДНАЯ СВЯЗЬ
Пространственное строение
1. Все атомы находятся в одной плоскости
2. Почти всегда атомы
ПЕПТИДНАЯ СВЯЗЬ
Пространственное строение
1. Все атомы находятся в одной плоскости
2. Почти всегда атомы

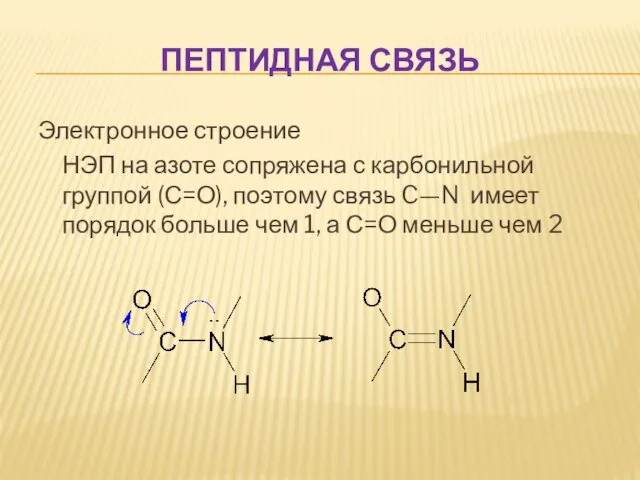
Слайд 50ПЕПТИДНАЯ СВЯЗЬ
Электронное строение
НЭП на азоте сопряжена с карбонильной группой (С=О), поэтому связь
ПЕПТИДНАЯ СВЯЗЬ
Электронное строение
НЭП на азоте сопряжена с карбонильной группой (С=О), поэтому связь
Слайд 51ПЕПТИДЫ И БЕЛКИ
Как и аминокислоты, пептиды и белки являются амфотерными соединениями, содержащими
ПЕПТИДЫ И БЕЛКИ
Как и аминокислоты, пептиды и белки являются амфотерными соединениями, содержащими

Слайд 52АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ ПЕПТИДОВ
МЕТОД ЭДМАНА
Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК с N-конца
АНАЛИЗ ПЕРВИЧНОЙ СТРУКТУРЫ ПЕПТИДОВ
МЕТОД ЭДМАНА
Используя фенилизотиоцианат (Ph-NCS) последовательно отщепляют АК с N-конца

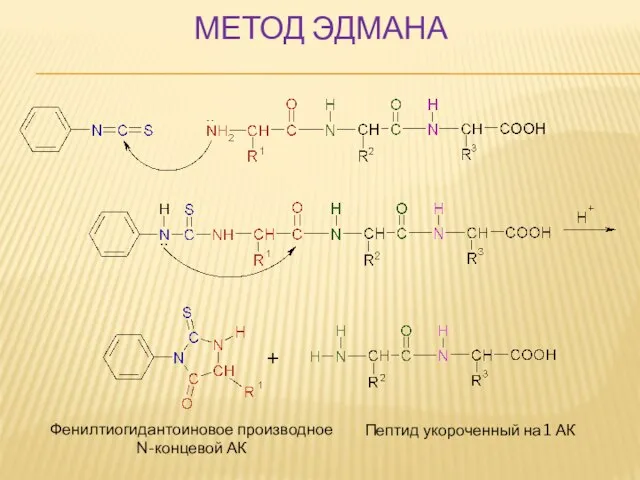
Слайд 53МЕТОД ЭДМАНА
Фенилтиогидантоиновое производное
N-концевой АК
Пептид укороченный на 1 АК
МЕТОД ЭДМАНА
Фенилтиогидантоиновое производное
N-концевой АК
Пептид укороченный на 1 АК
Слайд 54СИНТЕЗ ПЕПТИДОВ И БЕЛКОВ
Защита аминогруппы (БОК-защита)
Защита карбоксильной группы (образование сложных эфиров)
Образование
СИНТЕЗ ПЕПТИДОВ И БЕЛКОВ
Защита аминогруппы (БОК-защита)
Защита карбоксильной группы (образование сложных эфиров)
Образование

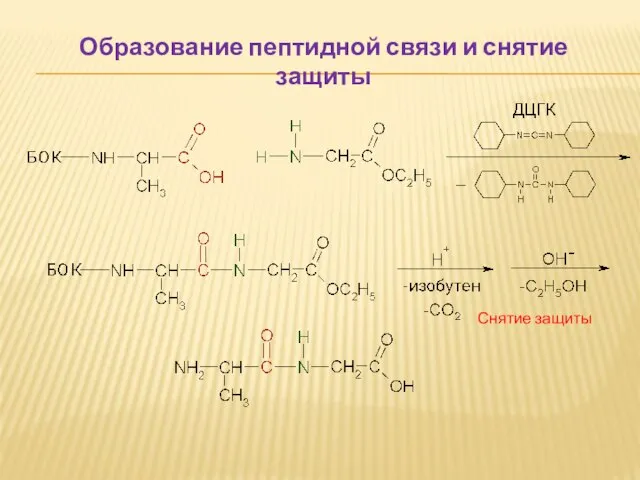
Слайд 55Снятие защиты
Образование пептидной связи и снятие защиты
Снятие защиты
Образование пептидной связи и снятие защиты
Слайд 56УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
первичная структура –
аминокислотная последовательность
вторичная структура – локальные высокоупорядоченные
УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
первичная структура –
аминокислотная последовательность
вторичная структура – локальные высокоупорядоченные

Слайд 57УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
УРОВНИ СТРУКТУРНОЙ ОРГАНИЗАЦИИ БЕЛКА
Слайд 58ПЕРВИЧНАЯ СТРУКТУРА
первичная структура –последовательность аминокислотных остатков в молекуле белка или пептида.
ПЕРВИЧНАЯ СТРУКТУРА
первичная структура –последовательность аминокислотных остатков в молекуле белка или пептида.

Слайд 59Blood smear in which the red cells show variation in size and
Blood smear in which the red cells show variation in size and
Слайд 60Количество возможных первичных структур ОГРОМНО!
20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих
Количество возможных первичных структур ОГРОМНО!
20 аминокислот могут дать примерно 100000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000000 белков, состоящих

Слайд 61ВТОРИЧНАЯ СТРУКТУРА
Вторичная структура- локальные высокоупорядоченные конформации белковой цепи – спирали и складчатые
ВТОРИЧНАЯ СТРУКТУРА
Вторичная структура- локальные высокоупорядоченные конформации белковой цепи – спирали и складчатые

Слайд 62Α-СПИРАЛЬ
Правые α-спирали полипептидной цепи стабилизируются водородными связями, где С=О группы остова
Α-СПИРАЛЬ
Правые α-спирали полипептидной цепи стабилизируются водородными связями, где С=О группы остова
Слайд 63ВОДОРОДНЫЕ СВЯЗИ В Α-СПИРАЛЯХ
-ВАЛ-ФЕН-ТИР-АЛА-ЛЕЙ-
Первый Второй Третий Четвёртый
Остатки АК:
α-спираль – 413 спираль:
ВОДОРОДНЫЕ СВЯЗИ В Α-СПИРАЛЯХ
-ВАЛ-ФЕН-ТИР-АЛА-ЛЕЙ-
Первый Второй Третий Четвёртый
Остатки АК:
α-спираль – 413 спираль:
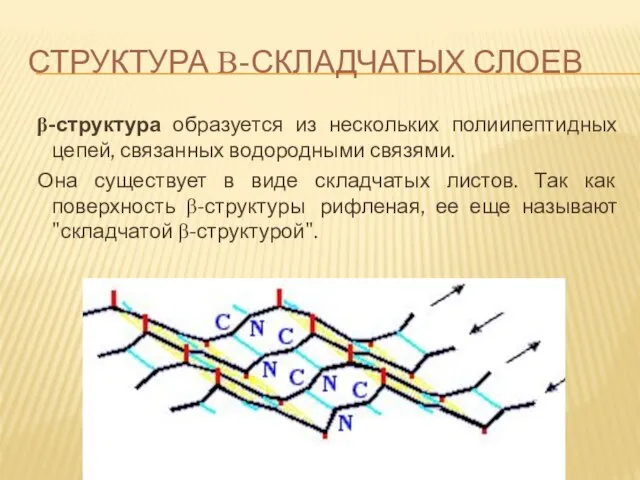
Слайд 64СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ
β-структура образуется из нескольких полиипептидных цепей, связанных водородными связями.
СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ
β-структура образуется из нескольких полиипептидных цепей, связанных водородными связями.
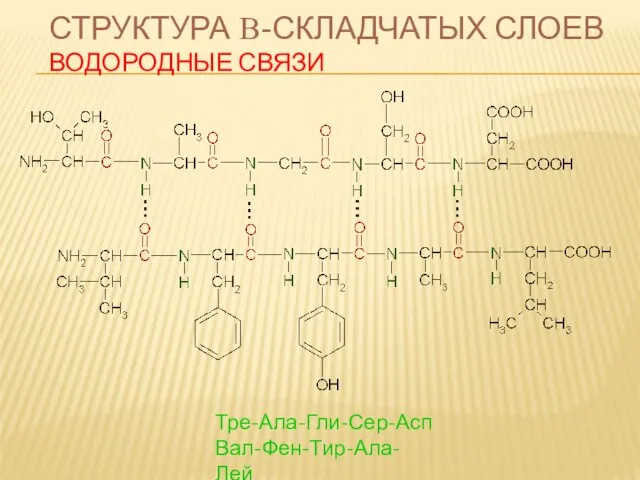
Слайд 65СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ
ВОДОРОДНЫЕ СВЯЗИ
Тре-Ала-Гли-Сер-Асп
Вал-Фен-Тир-Ала-Лей
СТРУКТУРА Β-СКЛАДЧАТЫХ СЛОЕВ
ВОДОРОДНЫЕ СВЯЗИ
Тре-Ала-Гли-Сер-Асп
Вал-Фен-Тир-Ала-Лей
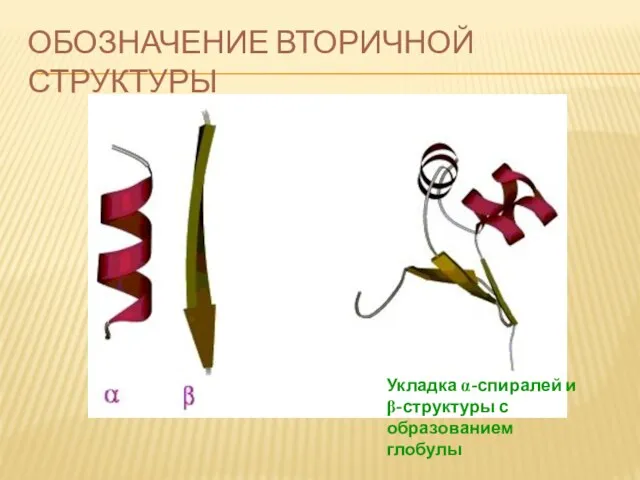
Слайд 66ОБОЗНАЧЕНИЕ ВТОРИЧНОЙ СТРУКТУРЫ
Укладка α-спиралей и
β-структуры с образованием глобулы
ОБОЗНАЧЕНИЕ ВТОРИЧНОЙ СТРУКТУРЫ
Укладка α-спиралей и
β-структуры с образованием глобулы
Слайд 67ТРЕТИЧНАЯ СТРУКТУРА
третичная структура – форма белковой молекулы; трёхмерная структура белка.
Укладка нерегулярных областей
ТРЕТИЧНАЯ СТРУКТУРА
третичная структура – форма белковой молекулы; трёхмерная структура белка.
Укладка нерегулярных областей

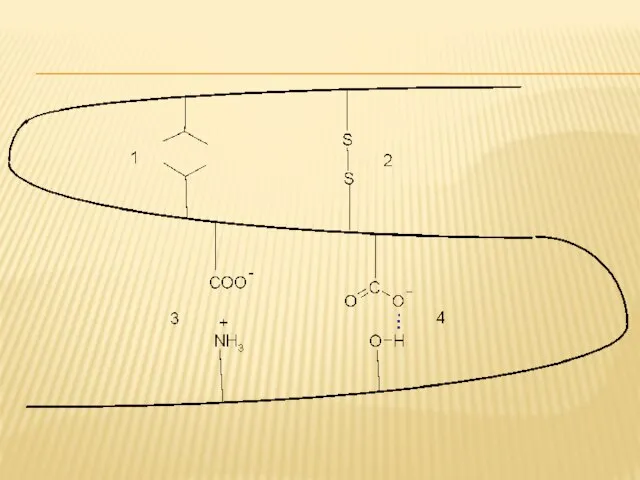
Слайд 68ВЗАИМОДЕЙСТВИЯ ОСТАТКОВ АК В ТРЕТИЧНОЙ СТРУКТУРЕ
Остатки АК в белковой глобуле взаимодействуют за
ВЗАИМОДЕЙСТВИЯ ОСТАТКОВ АК В ТРЕТИЧНОЙ СТРУКТУРЕ
Остатки АК в белковой глобуле взаимодействуют за

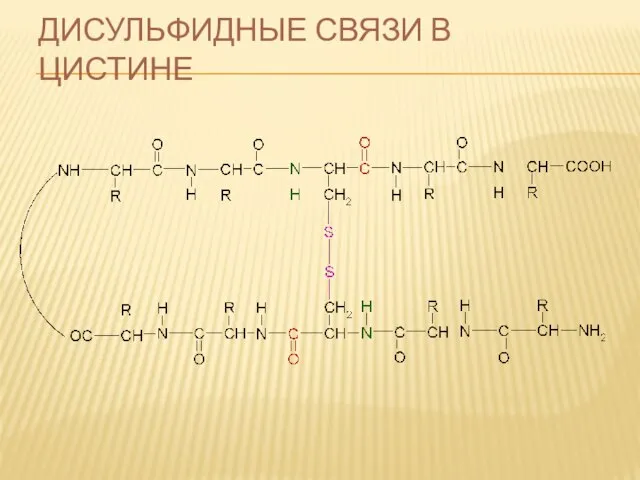
Слайд 70ДИСУЛЬФИДНЫЕ СВЯЗИ В ЦИСТИНЕ
ДИСУЛЬФИДНЫЕ СВЯЗИ В ЦИСТИНЕ
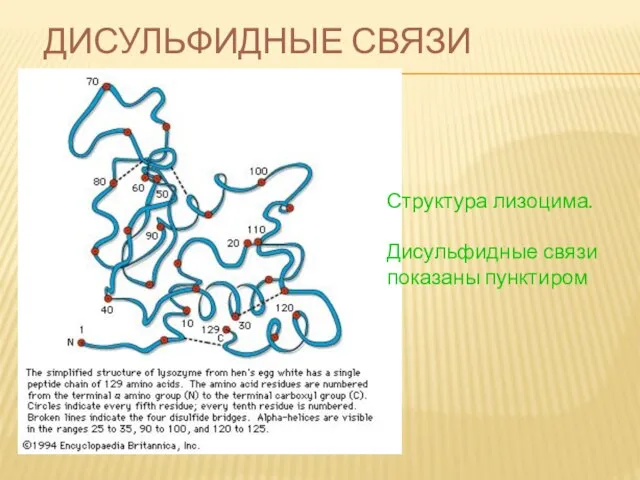
Слайд 71ДИСУЛЬФИДНЫЕ СВЯЗИ
Структура лизоцима.
Дисульфидные связи
показаны пунктиром
ДИСУЛЬФИДНЫЕ СВЯЗИ
Структура лизоцима.
Дисульфидные связи
показаны пунктиром
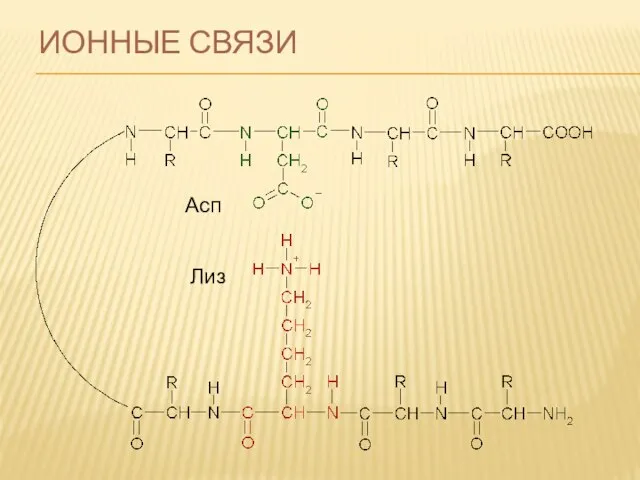
Слайд 72ИОННЫЕ СВЯЗИ
Асп
Лиз
ИОННЫЕ СВЯЗИ
Асп
Лиз
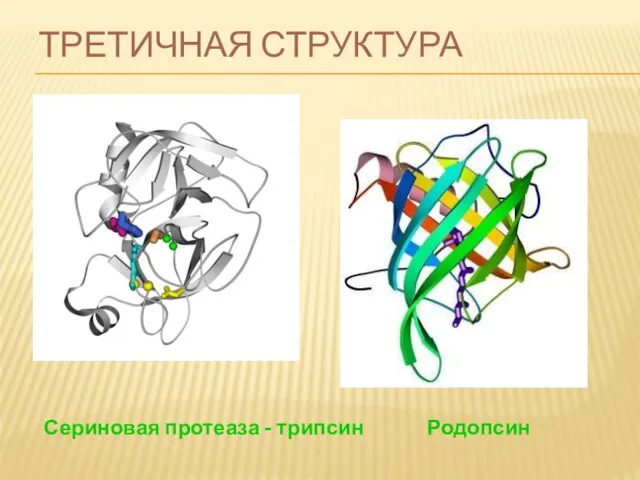
Слайд 73ТРЕТИЧНАЯ СТРУКТУРА
Сериновая протеаза - трипсин
Родопсин
ТРЕТИЧНАЯ СТРУКТУРА
Сериновая протеаза - трипсин
Родопсин
Слайд 74ТРЕТИЧНАЯ СТРУКТУРА
ацилтрансфераза пиктатлиаза С
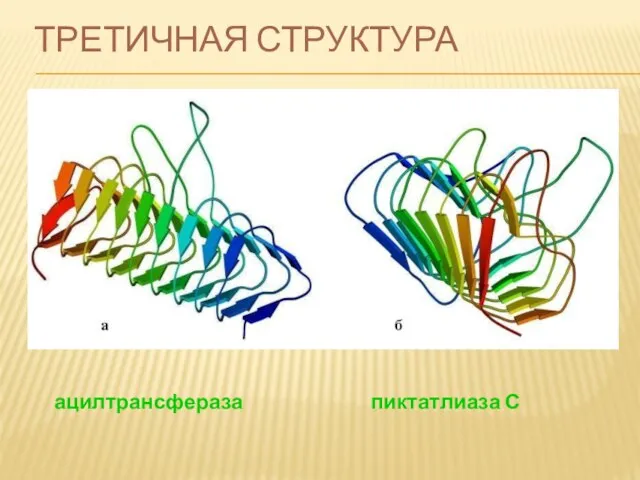
ТРЕТИЧНАЯ СТРУКТУРА
ацилтрансфераза пиктатлиаза С
Слайд 75ТРЕТИЧНАЯ СТРУКТУРА
α и β цепи гемоглобина лошади
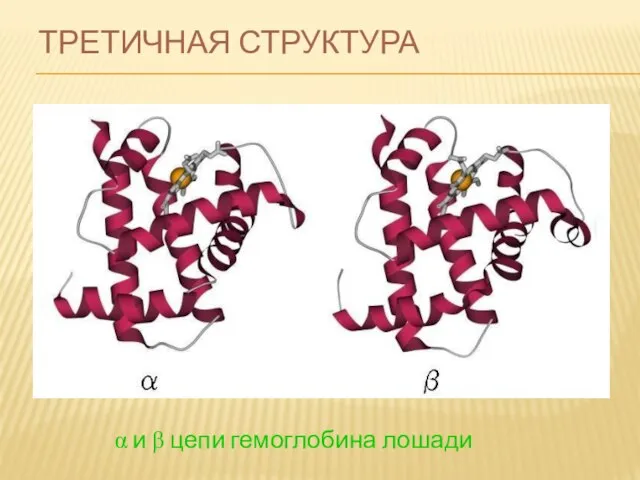
ТРЕТИЧНАЯ СТРУКТУРА
α и β цепи гемоглобина лошади
Слайд 76ТРЕТИЧНАЯ СТРУКТУРА
Порин
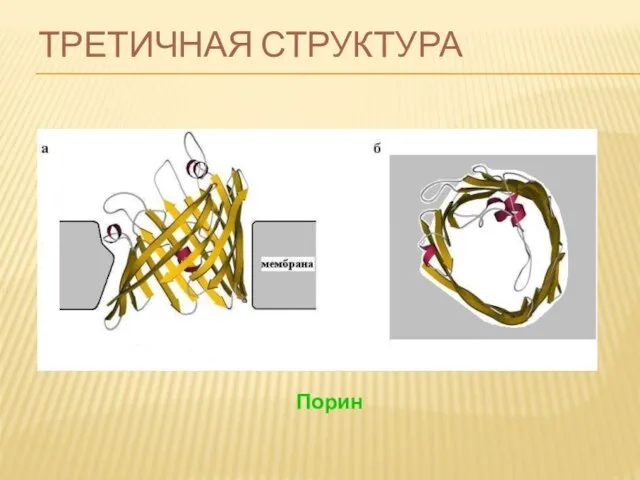
ТРЕТИЧНАЯ СТРУКТУРА
Порин
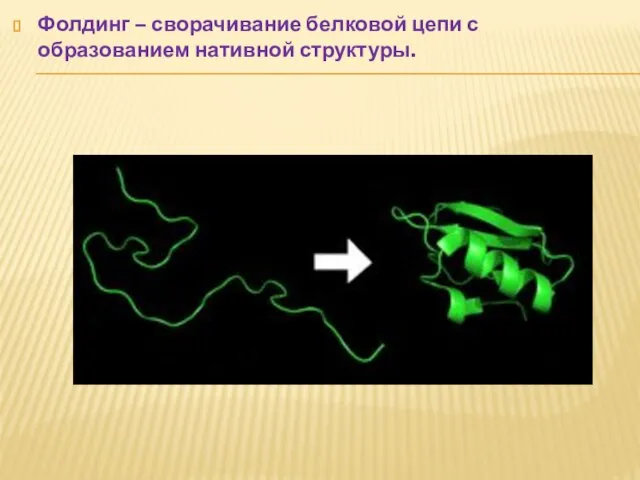
Слайд 77Фолдинг – сворачивание белковой цепи с образованием нативной структуры.
Фолдинг – сворачивание белковой цепи с образованием нативной структуры.
Слайд 78Неправильный фолдинг может привести к различным болезням, например к губчатым энцефалопатиям:
Синдром Крейцфельда-Якоба
Неправильный фолдинг может привести к различным болезням, например к губчатым энцефалопатиям:
Синдром Крейцфельда-Якоба

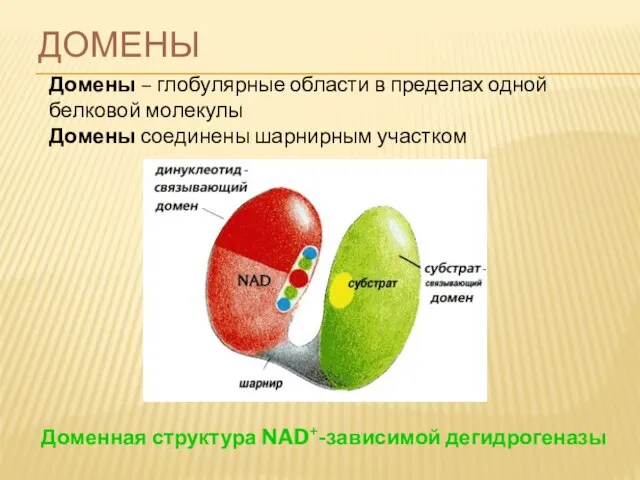
Слайд 79ДОМЕНЫ
Домены – глобулярные области в пределах одной белковой молекулы
Домены соединены шарнирным
ДОМЕНЫ
Домены – глобулярные области в пределах одной белковой молекулы
Домены соединены шарнирным
Слайд 80ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Четвертичная структура- агрегат нескольких белковых молекул образующих одну структуру
Взаимодействия: ионные, водородные,
ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Четвертичная структура- агрегат нескольких белковых молекул образующих одну структуру
Взаимодействия: ионные, водородные,

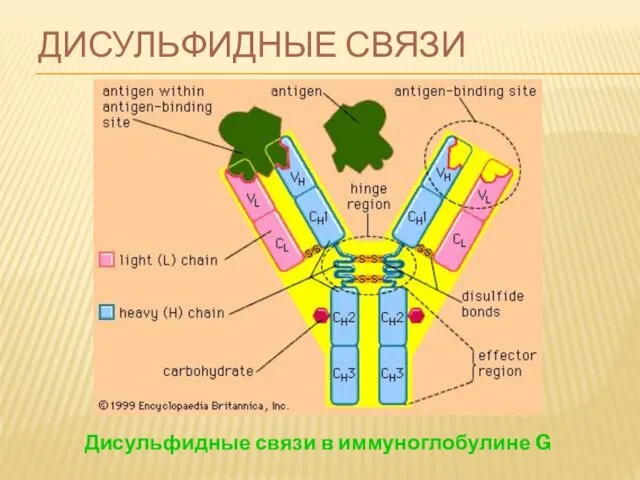
Слайд 81ДИСУЛЬФИДНЫЕ СВЯЗИ
Дисульфидные связи в иммуноглобулине G
ДИСУЛЬФИДНЫЕ СВЯЗИ
Дисульфидные связи в иммуноглобулине G
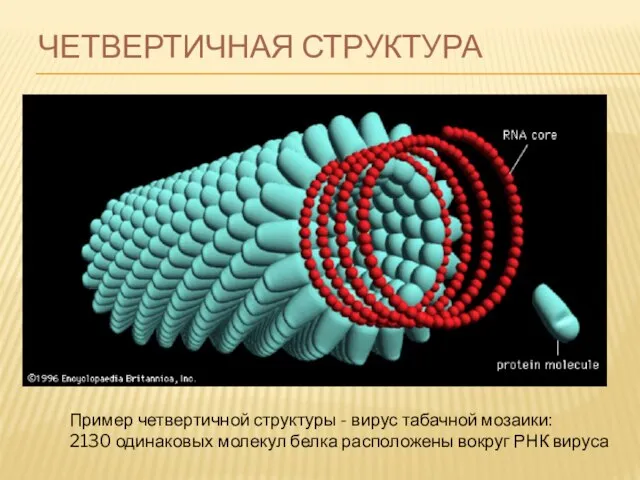
Слайд 82ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Пример четвертичной структуры - вирус табачной мозаики:
2130 одинаковых молекул белка расположены
ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Пример четвертичной структуры - вирус табачной мозаики:
2130 одинаковых молекул белка расположены
Слайд 83НУКЛЕИНОВЫЕ КИСЛОТЫ
НК- это полинуклеотиды,
т.е. биополимеры, мономерами которых являются нуклеотиды.
Нуклеотиды-фосфорные эфиры
НУКЛЕИНОВЫЕ КИСЛОТЫ
НК- это полинуклеотиды,
т.е. биополимеры, мономерами которых являются нуклеотиды.
Нуклеотиды-фосфорные эфиры

Слайд 84НУКЛЕИНОВЫЕ КИСЛОТЫ
НУКЛЕИНОВЫЕ КИСЛОТЫ
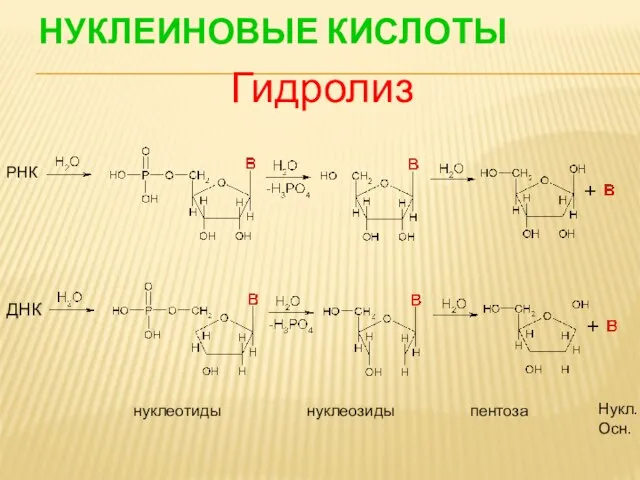
Слайд 85НУКЛЕИНОВЫЕ КИСЛОТЫ
РНК
ДНК
нуклеотиды нуклеозиды пентоза
Нукл.
Осн.
Гидролиз
НУКЛЕИНОВЫЕ КИСЛОТЫ
РНК
ДНК
нуклеотиды нуклеозиды пентоза
Нукл.
Осн.
Гидролиз
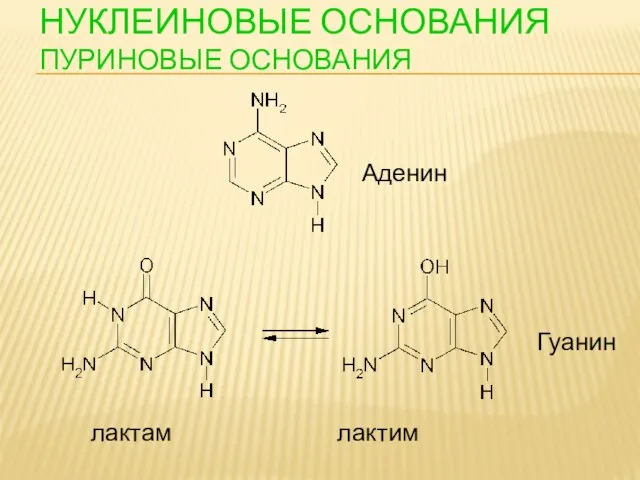
Слайд 86НУКЛЕИНОВЫЕ ОСНОВАНИЯ
ПУРИНОВЫЕ ОСНОВАНИЯ
Аденин
Гуанин
лактам лактим
НУКЛЕИНОВЫЕ ОСНОВАНИЯ
ПУРИНОВЫЕ ОСНОВАНИЯ
Аденин
Гуанин
лактам лактим
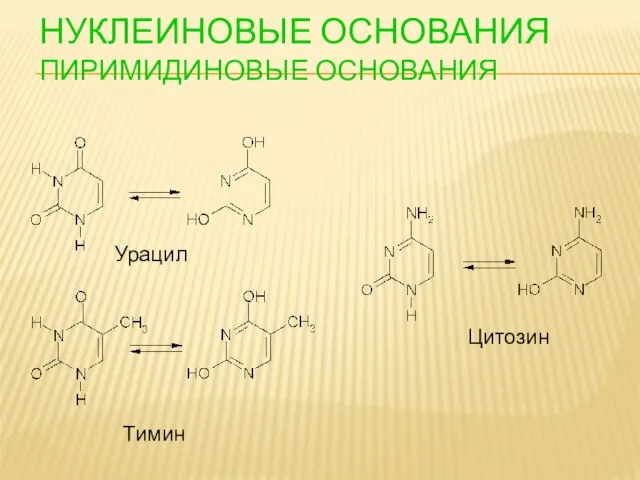
Слайд 87НУКЛЕИНОВЫЕ ОСНОВАНИЯ
ПИРИМИДИНОВЫЕ ОСНОВАНИЯ
Урацил
Тимин
Цитозин
НУКЛЕИНОВЫЕ ОСНОВАНИЯ
ПИРИМИДИНОВЫЕ ОСНОВАНИЯ
Урацил
Тимин
Цитозин
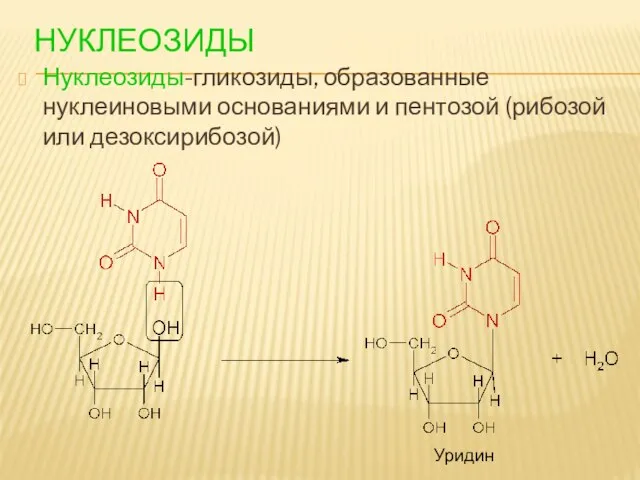
Слайд 88НУКЛЕОЗИДЫ
Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой)
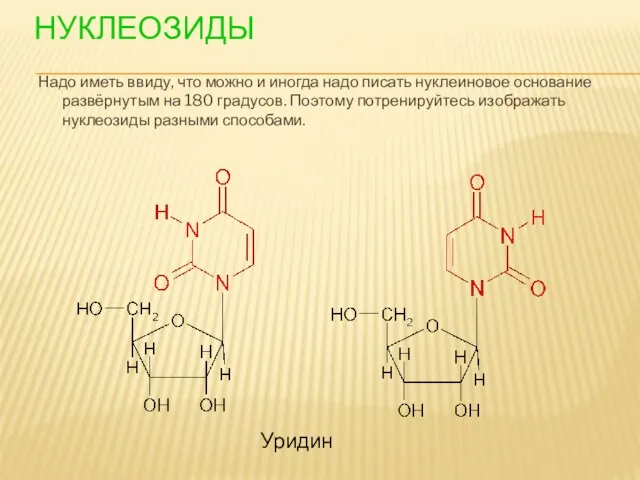
Уридин
НУКЛЕОЗИДЫ
Нуклеозиды-гликозиды, образованные нуклеиновыми основаниями и пентозой (рибозой или дезоксирибозой)
Уридин
Слайд 89НУКЛЕОЗИДЫ
Надо иметь ввиду, что можно и иногда надо писать нуклеиновое основание развёрнутым
НУКЛЕОЗИДЫ
Надо иметь ввиду, что можно и иногда надо писать нуклеиновое основание развёрнутым
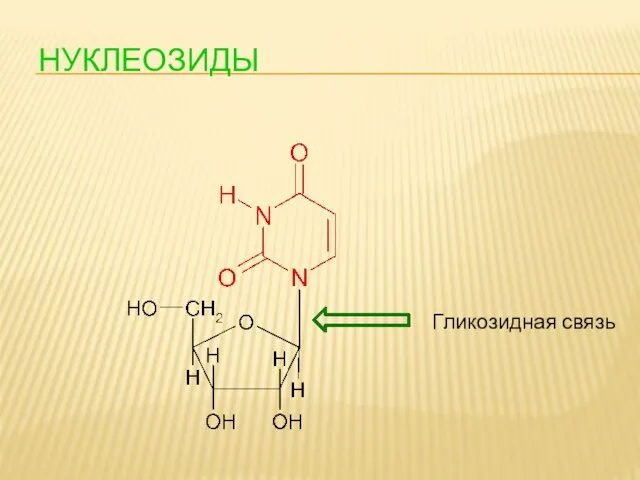
Слайд 90НУКЛЕОЗИДЫ
Гликозидная связь
НУКЛЕОЗИДЫ
Гликозидная связь
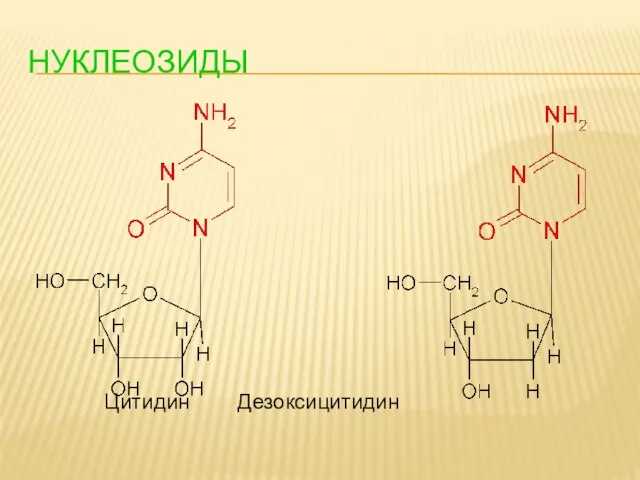
Слайд 91НУКЛЕОЗИДЫ
Цитидин Дезоксицитидин
НУКЛЕОЗИДЫ
Цитидин Дезоксицитидин
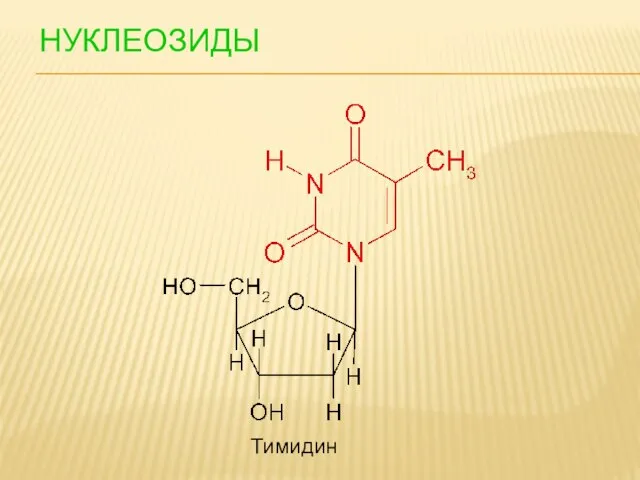
Слайд 92НУКЛЕОЗИДЫ
Тимидин
НУКЛЕОЗИДЫ
Тимидин
Слайд 93НУКЛЕОЗИДЫ
Аденозин Дезоксиаденозин
НУКЛЕОЗИДЫ
Аденозин Дезоксиаденозин
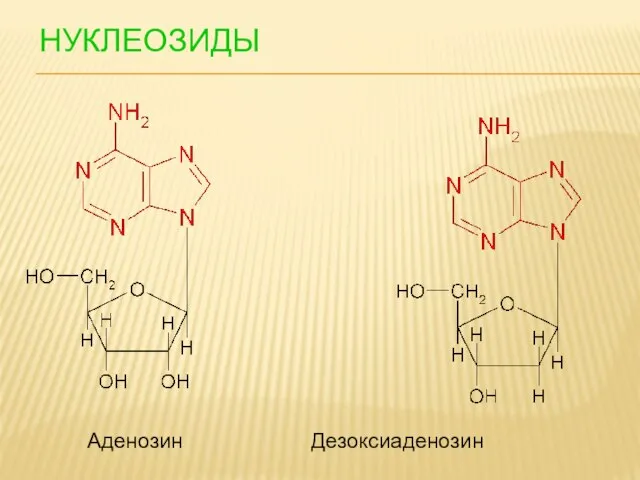
Слайд 94НУКЛЕОЗИДЫ
Аденозин Дезоксиаденозин
Аденозин и дезоксиаденозин можно изобразить и так:
НУКЛЕОЗИДЫ
Аденозин Дезоксиаденозин
Аденозин и дезоксиаденозин можно изобразить и так:
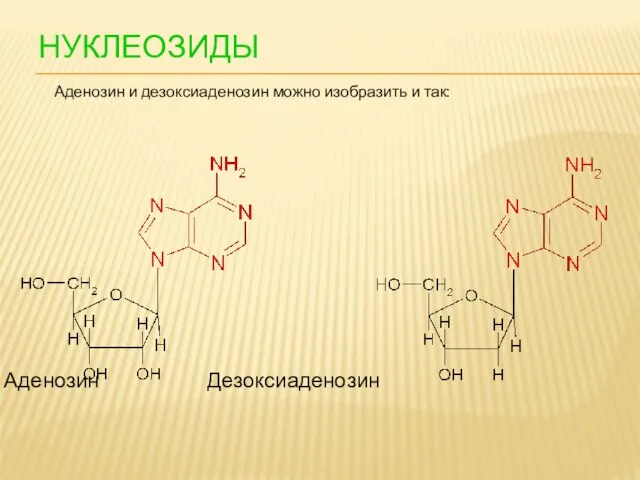
Слайд 95НУКЛЕОЗИДЫ
Гуанозин Дезоксигуанозин
НУКЛЕОЗИДЫ
Гуанозин Дезоксигуанозин
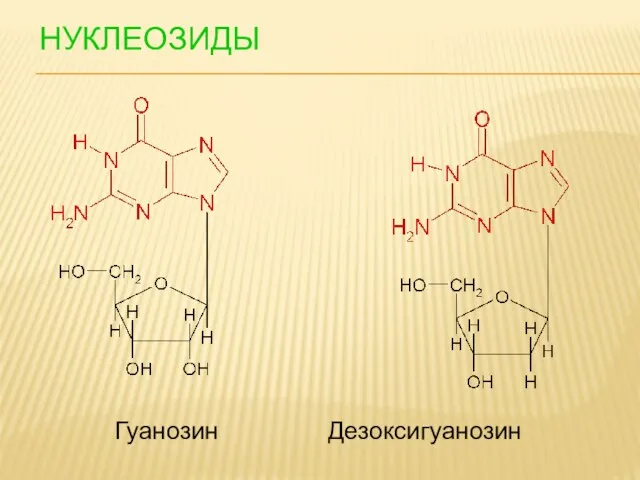
Слайд 96НУКЛЕОЗИДЫ
Гуанозин Дезоксигуанозин
Гуанозин и дезоксигуанозин можно изобразить и так:
НУКЛЕОЗИДЫ
Гуанозин Дезоксигуанозин
Гуанозин и дезоксигуанозин можно изобразить и так:
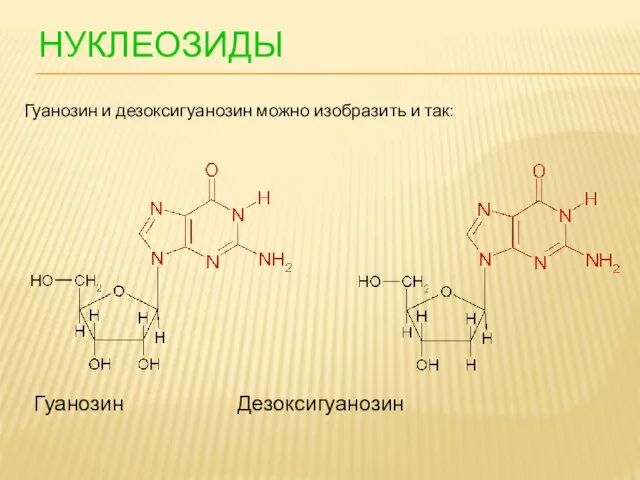
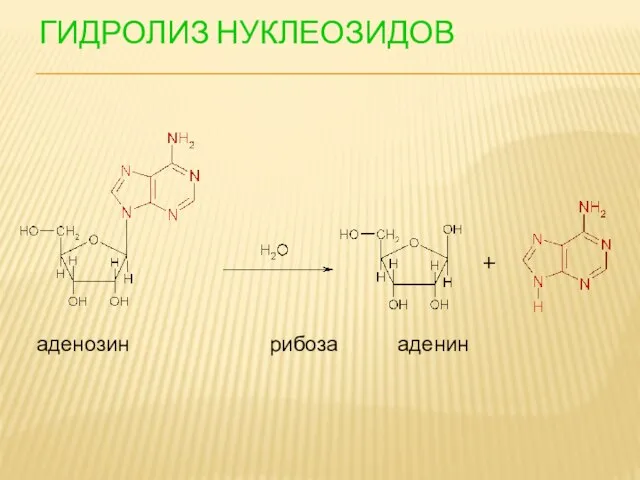
Слайд 97ГИДРОЛИЗ НУКЛЕОЗИДОВ
аденозин рибоза аденин
ГИДРОЛИЗ НУКЛЕОЗИДОВ
аденозин рибоза аденин
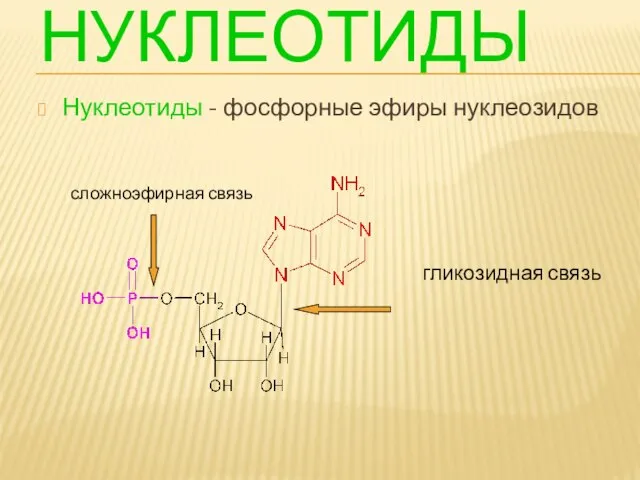
Слайд 98НУКЛЕОТИДЫ
Нуклеотиды - фосфорные эфиры нуклеозидов
гликозидная связь
сложноэфирная связь
НУКЛЕОТИДЫ
Нуклеотиды - фосфорные эфиры нуклеозидов
гликозидная связь
сложноэфирная связь
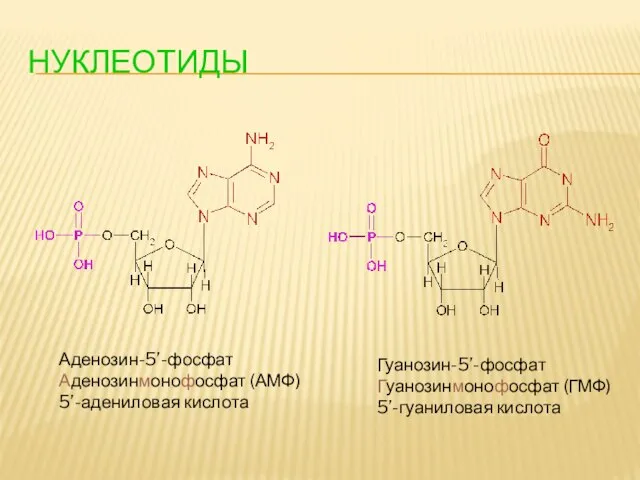
Слайд 99НУКЛЕОТИДЫ
Аденозин-5’-фосфат
Аденозинмонофосфат (АМФ)
5’-адениловая кислота
Гуанозин-5’-фосфат
Гуанозинмонофосфат (ГМФ)
5’-гуаниловая кислота
НУКЛЕОТИДЫ
Аденозин-5’-фосфат
Аденозинмонофосфат (АМФ)
5’-адениловая кислота
Гуанозин-5’-фосфат
Гуанозинмонофосфат (ГМФ)
5’-гуаниловая кислота
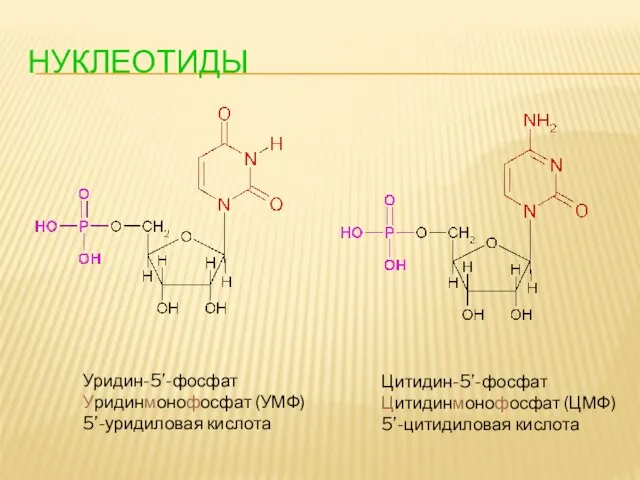
Слайд 100НУКЛЕОТИДЫ
Уридин-5’-фосфат
Уридинмонофосфат (УМФ)
5’-уридиловая кислота
Цитидин-5’-фосфат
Цитидинмонофосфат (ЦМФ)
5’-цитидиловая кислота
НУКЛЕОТИДЫ
Уридин-5’-фосфат
Уридинмонофосфат (УМФ)
5’-уридиловая кислота
Цитидин-5’-фосфат
Цитидинмонофосфат (ЦМФ)
5’-цитидиловая кислота
Слайд 101НУКЛЕОТИДЫ
НУКЛЕОТИДЫ
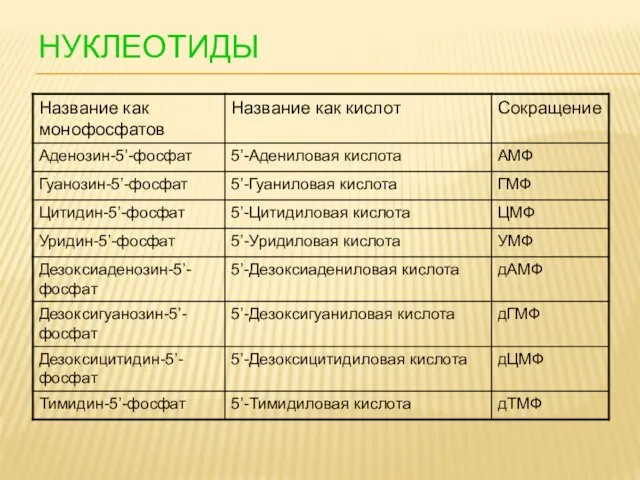
Слайд 102НУКЛЕОТИДЫ
Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам.
У других нуклеотидов в сокращённом
НУКЛЕОТИДЫ
Сокращения АМФ, ГМФ и т.д. относят к 5’-нуклеотидам.
У других нуклеотидов в сокращённом
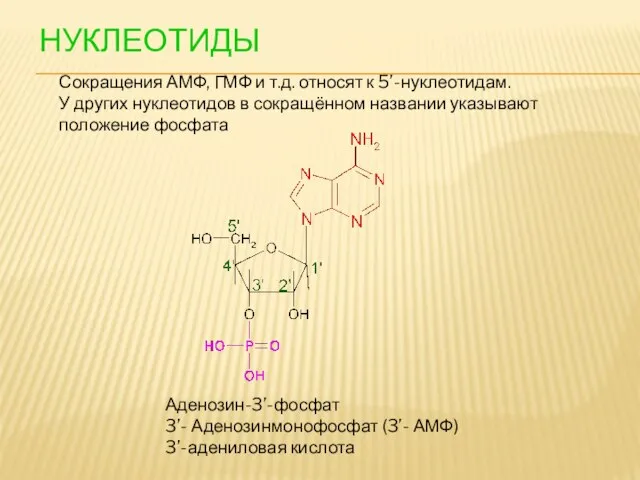
Слайд 103НУКЛЕОТИДЫ
цAMФ
НУКЛЕОТИДЫ
цAMФ
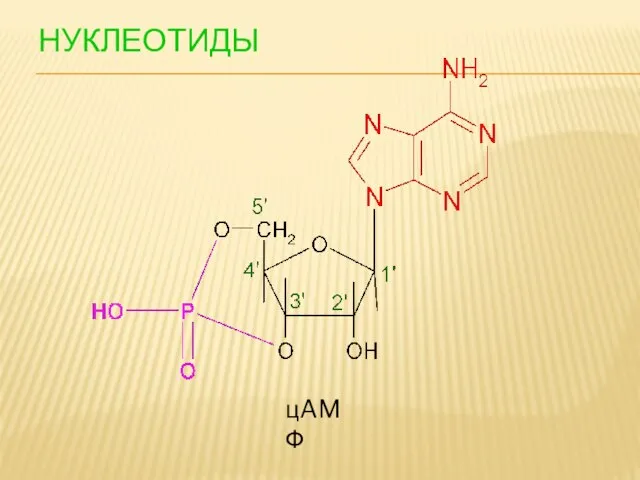
Слайд 104Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль вторичного посредника
Циклический аденозинмонофосфат (циклический AMФ, цAMФ, cAMP) — Циклический нуклеотид, играющий роль вторичного посредника

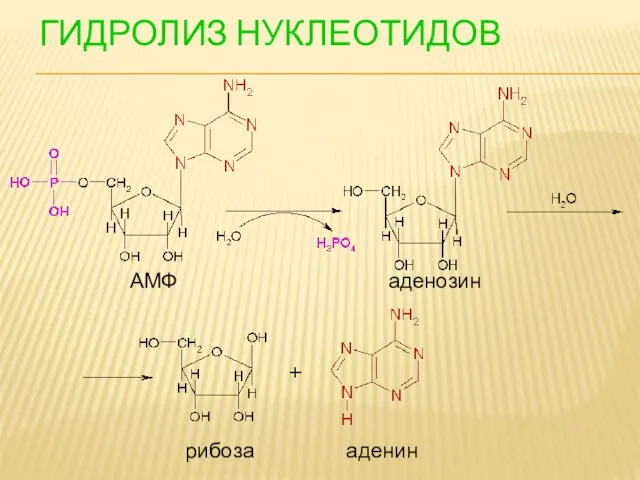
Слайд 105ГИДРОЛИЗ НУКЛЕОТИДОВ
АМФ
аденозин
рибоза
аденин
ГИДРОЛИЗ НУКЛЕОТИДОВ
АМФ
аденозин
рибоза
аденин
Слайд 106НУКЛЕОТИДЫ - КОФЕРМЕНТЫ
АМФ
АДФ
АТФ
Ангидридные
связи
Сложноэфирная
связь
АТФ – универсальный макроэрг
НУКЛЕОТИДЫ - КОФЕРМЕНТЫ
АМФ
АДФ
АТФ
Ангидридные
связи
Сложноэфирная
связь
АТФ – универсальный макроэрг
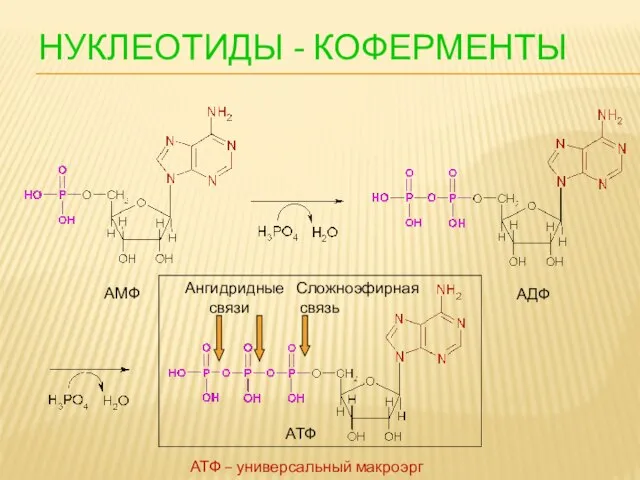
Слайд 107 Образование АТФ происходит
1. В результате субстратного фосфорилирования (гликолиз, одна реакция ЦТК)
2. Окислительное
Образование АТФ происходит
1. В результате субстратного фосфорилирования (гликолиз, одна реакция ЦТК)
2. Окислительное

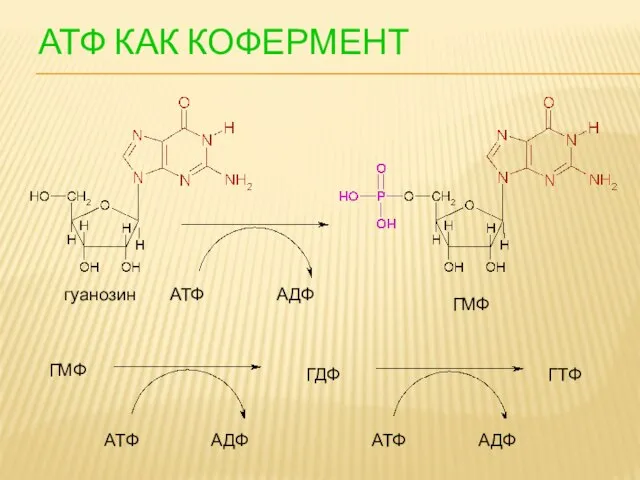
Слайд 108АТФ КАК КОФЕРМЕНТ
АТФ АДФ
АТФ АДФ
АТФ АДФ
ГМФ
ГДФ
ГТФ
ГМФ
гуанозин
АТФ КАК КОФЕРМЕНТ
АТФ АДФ
АТФ АДФ
АТФ АДФ
ГМФ
ГДФ
ГТФ
ГМФ
гуанозин
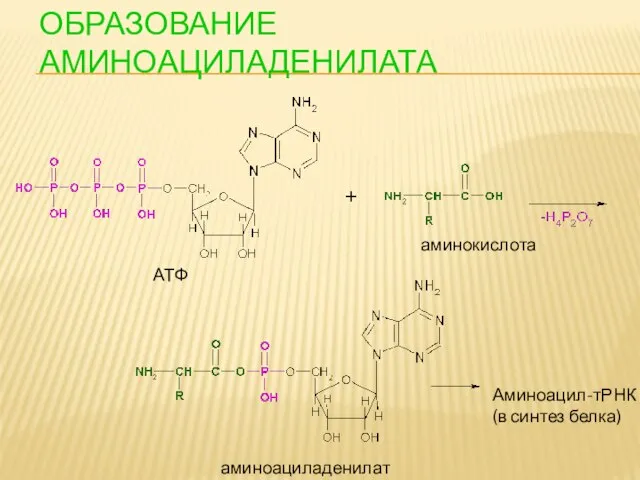
Слайд 109ОБРАЗОВАНИЕ АМИНОАЦИЛАДЕНИЛАТА
АТФ
аминокислота
аминоациладенилат
Аминоацил-тРНК
(в синтез белка)
ОБРАЗОВАНИЕ АМИНОАЦИЛАДЕНИЛАТА
АТФ
аминокислота
аминоациладенилат
Аминоацил-тРНК
(в синтез белка)
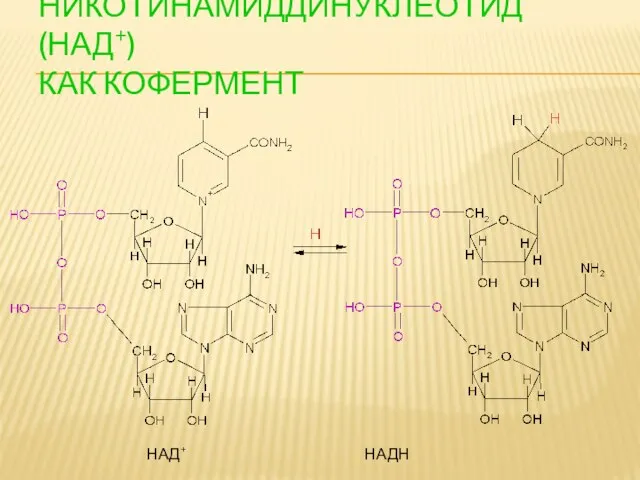
Слайд 110НИКОТИНАМИДДИНУКЛЕОТИД (НАД+)
КАК КОФЕРМЕНТ
НАД+ НАДН
НИКОТИНАМИДДИНУКЛЕОТИД (НАД+)
КАК КОФЕРМЕНТ
НАД+ НАДН
Слайд 111ПЕРВИЧНАЯ СТРУКТУРА НК
Первичная структура НК - последовательность нуклеотидов
5’- конец ТГАЦТААГТАЦЦ 3’-конец
ПЕРВИЧНАЯ СТРУКТУРА НК
Первичная структура НК - последовательность нуклеотидов
5’- конец ТГАЦТААГТАЦЦ 3’-конец

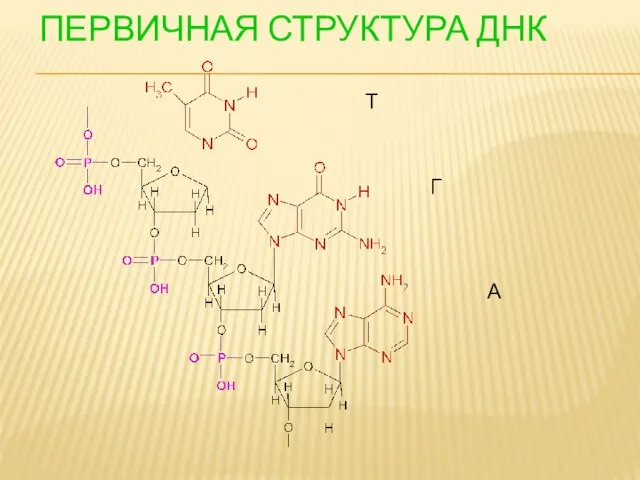
Слайд 112ПЕРВИЧНАЯ СТРУКТУРА ДНК
Т
Г
А
ПЕРВИЧНАЯ СТРУКТУРА ДНК
Т
Г
А
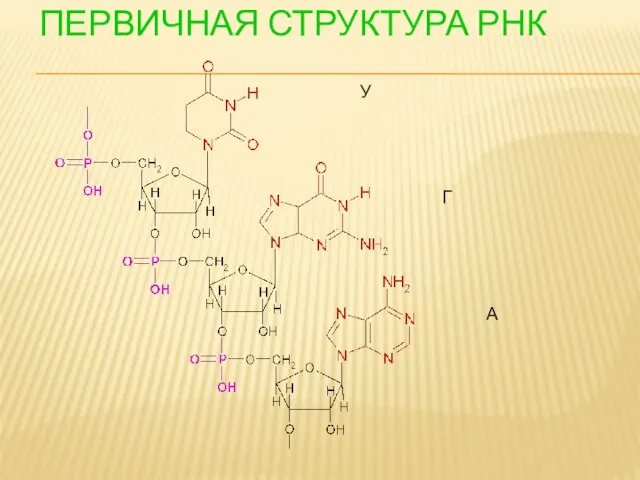
Слайд 113ПЕРВИЧНАЯ СТРУКТУРА РНК
У
Г
А
ПЕРВИЧНАЯ СТРУКТУРА РНК
У
Г
А
Слайд 114ВТОРИЧНАЯ СТРУКТУРА
Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953)
Две цепи
ВТОРИЧНАЯ СТРУКТУРА
Вторичная структура ДНК – двойная правая спираль (Уотсон, Крик, 1953)
Две цепи

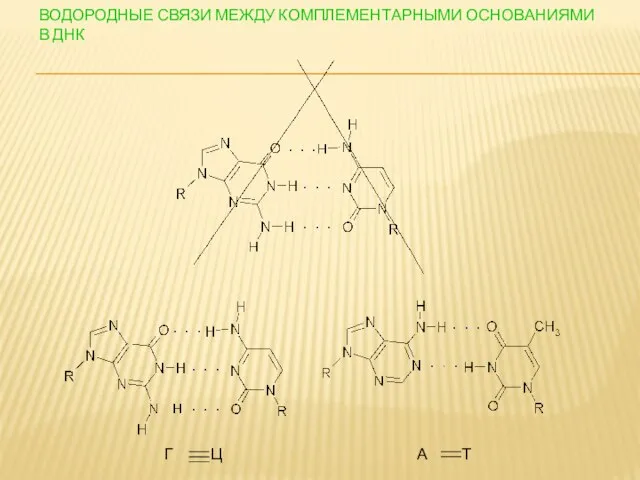
Слайд 115ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ КОМПЛЕМЕНТАРНЫМИ ОСНОВАНИЯМИ В ДНК
Г Ц
А Т
ВОДОРОДНЫЕ СВЯЗИ МЕЖДУ КОМПЛЕМЕНТАРНЫМИ ОСНОВАНИЯМИ В ДНК
Г Ц
А Т
 ОАО Туймазинский мясокомбинат
ОАО Туймазинский мясокомбинат Новая форма ГИА
Новая форма ГИА Права и обязанности в Конституции России
Права и обязанности в Конституции России Изделия в технике канзаши и в технике холодного фарфора
Изделия в технике канзаши и в технике холодного фарфора это надо скинуть шкарину
это надо скинуть шкарину Презентация на тему Изобразительное искусство в семье пластических искусств
Презентация на тему Изобразительное искусство в семье пластических искусств  Методы контроля при занятии физической культурой
Методы контроля при занятии физической культурой Основы религиозных культур и светской этики.
Основы религиозных культур и светской этики. Фотоаппарат. СРС
Фотоаппарат. СРС Проектирование электронного контента: основные принципы электронные курсы банк тестов
Проектирование электронного контента: основные принципы электронные курсы банк тестов ФОРЕКС - производный финансовый инструмент, в основе которого лежит обязательство компании-маркетмейкера выплатить клиенту разни
ФОРЕКС - производный финансовый инструмент, в основе которого лежит обязательство компании-маркетмейкера выплатить клиенту разни The role of the service industry in the modern Economy
The role of the service industry in the modern Economy  Интернет-проект «Забытый памятник незабытой войны»
Интернет-проект «Забытый памятник незабытой войны» Генерация идей для создания успешных проектов
Генерация идей для создания успешных проектов Сетевые технологии: терминология, устройства
Сетевые технологии: терминология, устройства Полисахариды
Полисахариды Обрушение смыслов: как вернуть себе желание жить после большой потери?
Обрушение смыслов: как вернуть себе желание жить после большой потери? Модели реализации межшкольными учебными комбинатами профориентационной работы со школьниками в рамках сетевого взаимодействия
Модели реализации межшкольными учебными комбинатами профориентационной работы со школьниками в рамках сетевого взаимодействия  Уравнение плоскости в пространстве
Уравнение плоскости в пространстве Политика. Повторение. 9 класс
Политика. Повторение. 9 класс Технология контроля поверхностного слоя песчано-глинистой литейной формы
Технология контроля поверхностного слоя песчано-глинистой литейной формы В мире «кривых». Работа Яковлевой Яны 10b 2009 г.
В мире «кривых». Работа Яковлевой Яны 10b 2009 г. Оптические приборы. Глаз
Оптические приборы. Глаз Математические модели и оптимальные процессы в макросистемах (термодинамика и экономика)
Математические модели и оптимальные процессы в макросистемах (термодинамика и экономика) От А до К
От А до К 7. Презентация 2 этап (пример)
7. Презентация 2 этап (пример) Этюд простой архитектурной постройки сельского типа
Этюд простой архитектурной постройки сельского типа Расследование преступлений, совершенных организованными преступными группами
Расследование преступлений, совершенных организованными преступными группами