- Современная схема кроветворения. Регуляция гемопоэза

Содержание
- 2. Современная теория кроветворения Современная теория кроветворения базируется на унитарной теории А.А. Максимова (1918), согласно которой все
- 3. С Х Е К М Р А О В Е Т В О Р Е Н
- 4. Современная теория кроветворения Нормальное кроветворение поликлональное, т. е. осуществляется одновременно многими клонами. Размер индивидуального клона -
- 5. Дифференцировка клеток гемопоэза Клетки гемопоэза условно подразделены на 5-6 отделов, границы между которыми весьма размыты, а
- 6. Дифференцировка клеток гемопоэза I отдел – тотипотентная эмбриональная стволовая клетка (ЭСК), находится на самом верху иерархической
- 7. Стволовые кроветворные клетки СКК закладываются в период эмбриогенеза и расходуются последовательно, образуя сменяющие друг друга клоны
- 8. Стволовые кроветворные клетки Отдел СКК гетерогенен, представлен 2 категориями предшественников, обладающих различным пролиферативным потенциалом. Основная масса
- 9. Стволовые кроветворные клетки Гетерогенность пула СКК и степень их дифференцировки устанавливается на основе экспрессии ряда дифференцировочных
- 10. Стволовые кроветворные клетки Одним из основных методов изучения СКК является метод колониеобразования in vivo или in
- 11. Дифференцировка клеток гемопоэза III отдел - По мере снижения пролиферативного потенциала СКК дифференцируются в полиолигопотентные коммитированные
- 12. Дифференцировка клеток гемопоэза Клетки IV отдела - монопотентные коммитированные предшественники являются родоначальными для одного ростка гемопоэза:
- 13. СКК и клетки-предшественники обладают способностью к миграции - выходу в кровь и возвращению в костный мозг,
- 14. Дифференцировка клеток гемопоэза V отдел морфологически распознаваемых клеток включает: дифференцирующиеся, созревающие зрелые клетки всех 8 клеточных
- 15. Регуляция гемопоэза Кроветворная ткань - динамичная, постоянно обновляющаяся клеточная система организма. В минуту в кроветворных органах
- 16. Регуляция гемопоэза Гемопоэз инициируется ростовыми факторами, цитокинами и непрерывно поддерживается благодаря пулу СКК. Стволовые кроветворные клетки
- 17. Регуляция гемопоэза Обновление СКК происходит медленно и при готовности к дифференцировке (процесс коммитирования), они выходят из
- 18. Схема регуляции эритро- и гранулоцито-монопоэза
- 19. Регуляторы гемопоэза Выделяют позитивные и негативные регуляторы гемопоэза. Позитивные регуляторы необходимы: для выживания СКК и их
- 20. Факторы регуляции гемопоэза Факторы регуляции гемопоэза подразделяются на короткодистантные (для СКК) и дальнодействующие для коммитированных предшественников
- 21. Факторы регуляции гемопоэза 2. Линейно-неспецифические факторы: ИЛ-3, ИЛ-4, ГМ-КСФ (для гранулоцитомонопоэза). 3. Позднедействующие линейно-специфические факторы, которые
- 23. Скачать презентацию
Слайд 2Современная теория кроветворения
Современная теория кроветворения базируется на унитарной теории А.А. Максимова (1918),
Современная теория кроветворения
Современная теория кроветворения базируется на унитарной теории А.А. Максимова (1918),

Слайд 3С
Х
Е К
М Р
А О
В
Е
Т
В
О
Р
Е
Н
И
Я
Х
Е К
М Р
А О
В
Е
Т
В
О
Р
Е
Н
И
Я
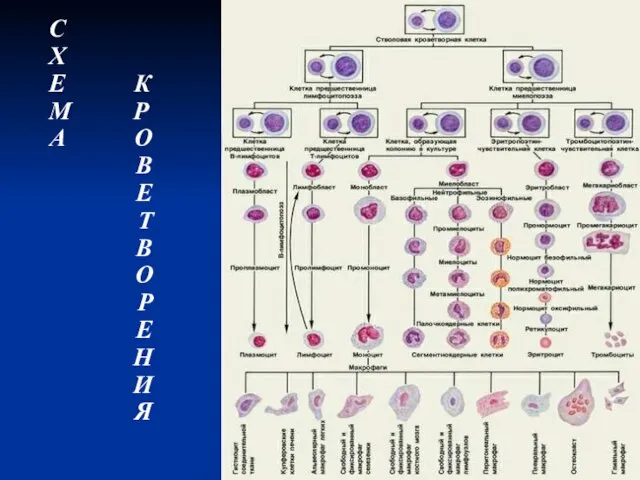
Слайд 4Современная теория кроветворения
Нормальное кроветворение поликлональное, т. е. осуществляется одновременно многими клонами.
Размер индивидуального
Современная теория кроветворения
Нормальное кроветворение поликлональное, т. е. осуществляется одновременно многими клонами.
Размер индивидуального

Слайд 5Дифференцировка клеток гемопоэза
Клетки гемопоэза условно подразделены на 5-6 отделов, границы между которыми
Дифференцировка клеток гемопоэза
Клетки гемопоэза условно подразделены на 5-6 отделов, границы между которыми

Слайд 6Дифференцировка клеток гемопоэза
I отдел – тотипотентная эмбриональная стволовая клетка (ЭСК), находится на
Дифференцировка клеток гемопоэза
I отдел – тотипотентная эмбриональная стволовая клетка (ЭСК), находится на

Слайд 7Стволовые кроветворные клетки
СКК закладываются в период эмбриогенеза и расходуются последовательно, образуя сменяющие
Стволовые кроветворные клетки
СКК закладываются в период эмбриогенеза и расходуются последовательно, образуя сменяющие

Слайд 8Стволовые кроветворные клетки
Отдел СКК гетерогенен, представлен 2 категориями предшественников, обладающих различным пролиферативным
Стволовые кроветворные клетки
Отдел СКК гетерогенен, представлен 2 категориями предшественников, обладающих различным пролиферативным

Слайд 9Стволовые кроветворные клетки
Гетерогенность пула СКК и степень их дифференцировки устанавливается на основе
Стволовые кроветворные клетки
Гетерогенность пула СКК и степень их дифференцировки устанавливается на основе

Слайд 10Стволовые кроветворные клетки
Одним из основных методов изучения СКК является метод колониеобразования in
Стволовые кроветворные клетки
Одним из основных методов изучения СКК является метод колониеобразования in

Слайд 11Дифференцировка клеток гемопоэза
III отдел - По мере снижения пролиферативного потенциала СКК дифференцируются
Дифференцировка клеток гемопоэза
III отдел - По мере снижения пролиферативного потенциала СКК дифференцируются

Слайд 12Дифференцировка клеток гемопоэза
Клетки IV отдела - монопотентные коммитированные предшественники являются родоначальными для
Дифференцировка клеток гемопоэза
Клетки IV отдела - монопотентные коммитированные предшественники являются родоначальными для

Слайд 13СКК и клетки-предшественники обладают способностью к миграции - выходу в кровь и
СКК и клетки-предшественники обладают способностью к миграции - выходу в кровь и

Слайд 14Дифференцировка клеток гемопоэза
V отдел морфологически распознаваемых клеток включает:
дифференцирующиеся,
созревающие
зрелые клетки
Дифференцировка клеток гемопоэза
V отдел морфологически распознаваемых клеток включает:
дифференцирующиеся,
созревающие
зрелые клетки

Слайд 15Регуляция гемопоэза
Кроветворная ткань - динамичная, постоянно обновляющаяся клеточная система организма.
В минуту
Регуляция гемопоэза
Кроветворная ткань - динамичная, постоянно обновляющаяся клеточная система организма.
В минуту

Слайд 16Регуляция гемопоэза
Гемопоэз инициируется ростовыми факторами, цитокинами и непрерывно поддерживается благодаря пулу СКК.
Регуляция гемопоэза
Гемопоэз инициируется ростовыми факторами, цитокинами и непрерывно поддерживается благодаря пулу СКК.

Слайд 17Регуляция гемопоэза
Обновление СКК происходит медленно и при готовности к дифференцировке (процесс коммитирования),
Регуляция гемопоэза
Обновление СКК происходит медленно и при готовности к дифференцировке (процесс коммитирования),

Слайд 18Схема регуляции эритро- и гранулоцито-монопоэза
Схема регуляции эритро- и гранулоцито-монопоэза
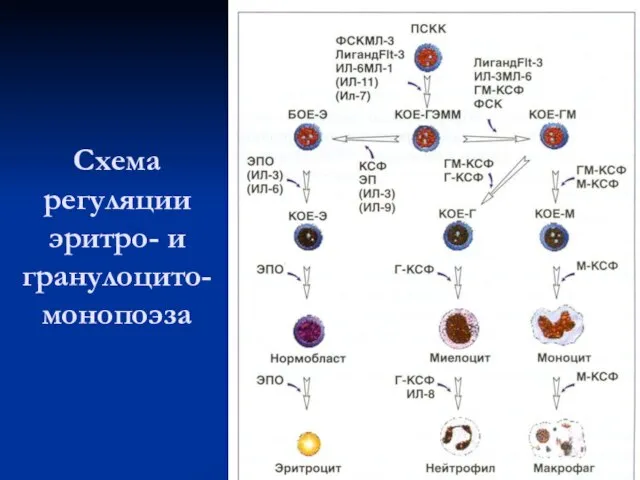
Слайд 19Регуляторы гемопоэза
Выделяют позитивные и негативные регуляторы гемопоэза.
Позитивные регуляторы необходимы:
для выживания
Регуляторы гемопоэза
Выделяют позитивные и негативные регуляторы гемопоэза.
Позитивные регуляторы необходимы:
для выживания

Слайд 20Факторы регуляции гемопоэза
Факторы регуляции гемопоэза подразделяются на короткодистантные (для СКК) и дальнодействующие
Факторы регуляции гемопоэза
Факторы регуляции гемопоэза подразделяются на короткодистантные (для СКК) и дальнодействующие

Слайд 21Факторы регуляции гемопоэза
2. Линейно-неспецифические факторы:
ИЛ-3,
ИЛ-4,
ГМ-КСФ (для гранулоцитомонопоэза).
3. Позднедействующие линейно-специфические факторы,
Факторы регуляции гемопоэза
2. Линейно-неспецифические факторы:
ИЛ-3,
ИЛ-4,
ГМ-КСФ (для гранулоцитомонопоэза).
3. Позднедействующие линейно-специфические факторы,

 психол культура
психол культура Всероссийская перепись населения
Всероссийская перепись населения Человек в экономических отношениях 7 класс
Человек в экономических отношениях 7 класс Образцы гистологических препаратов
Образцы гистологических препаратов Организация опытно-экспериментальной и инновационной деятельности в образовательных учреждениях Прохоровского районав 2011-2012 у
Организация опытно-экспериментальной и инновационной деятельности в образовательных учреждениях Прохоровского районав 2011-2012 у Тренажёр
Тренажёр Система (гр. Systema — целое, составленное из частей; соединение) — множество закономерно связанных друг с другом элементов
Система (гр. Systema — целое, составленное из частей; соединение) — множество закономерно связанных друг с другом элементов Фотошкола Пикча. Профессия - фотограф
Фотошкола Пикча. Профессия - фотограф Коучинг Катализирующая роль коучинга в становлении и развитии бизнес-объединений и усовершенствовании образовательных програ
Коучинг Катализирующая роль коучинга в становлении и развитии бизнес-объединений и усовершенствовании образовательных програ Читаешь книги – хорошо. Не читаешь - плохо
Читаешь книги – хорошо. Не читаешь - плохо Кейс. Таргетированная реклама ВК. СК-Моторс (автосалон)
Кейс. Таргетированная реклама ВК. СК-Моторс (автосалон) баннер
баннер Управленческий учет и его место в информационной и учетной системах организации
Управленческий учет и его место в информационной и учетной системах организации Презентация на тему Китай в средние века (6 класс)
Презентация на тему Китай в средние века (6 класс) Костюм древнего египтянина
Костюм древнего египтянина Дизайн гостиничного домика (фотографии)
Дизайн гостиничного домика (фотографии) ...В слове МЫ сто тысяч Я.
...В слове МЫ сто тысяч Я. Презентация на тему Диагностика готовности к школе как условие успешной адаптации первоклассников
Презентация на тему Диагностика готовности к школе как условие успешной адаптации первоклассников Презентация о себе
Презентация о себе Закуски из овощей
Закуски из овощей Уголовное право
Уголовное право Правописание НН-Н в полных прилагательных и причастиях
Правописание НН-Н в полных прилагательных и причастиях От сердца к сердцу
От сердца к сердцу Ориентирование на местности (2 класс)
Ориентирование на местности (2 класс) Кожевенное сырье
Кожевенное сырье Лесопарк – территория здоровья
Лесопарк – территория здоровья Предметно-развивающая среда
Предметно-развивающая среда Color chart and structure chart
Color chart and structure chart