- Стійкість рослин

Содержание
- 2. Види стійкості Стрес Солестійкість Зимостійкість(холодо і морозостійкість та інші несприятливі фактори зими) Посухостійкість Стійкість до промислових
- 3. Cтрес - фактор впливу зовнiшнiх умов середовища, який викликає негативний вплив на рослини. Основнi поняття i
- 4. В дiї на тваринний органiзм i рослини є подiбнiсть i вiдмiнностi - у обох є 3
- 5. Специфічні і неспецифічні реакції на стрес Дiя любого стресу викликає ряд пристосувальних реакцiй. При малiй iнтенсивностi
- 6. На дiю бiльшостi стресiв i особливо сильних i при короткому часi дiї рослини вiдповiдають неспецифiчними реакцiями,
- 7. За походженням всi екологiчнi фактори подiляють на такi групи: 1.Абiотичнi - клiматичнi, едафiчнi, топографiчнi 2.Бiотичнi: фiтогеннi(
- 8. За характером дiї стрес як екологiчний фактор може бути: прямодiючим i опосередкованим ( вплив на органiзм
- 9. Вплив стресу Вплив практично всiх стресiв або факторiв можна вкласти в двi кривi - крива Сакса
- 10. В той же час по вiдношенню до дiї фактора *подразника) рослини можуть бути: стенобiонтними - зона
- 11. Сукупнiсть екологiчних факторiв називають дiючим середовищем, або середовищем iснування. Таких середовищ iснування є чотири - водне,
- 12. Здатність рослини переносити дію несприятливих факторів і давати в таких умовах потомство називається стійкістю або стрес-толерантністю
- 13. Стійкість є кінцевим результатом адаптації. Адаптація (лат. adaptio - пристосування, прикладання) - це генетично детермінований процес
- 14. Вибір рослиною стратегії (способу) адаптації залежить від багатьох факторів. Однак ключовим фактором є час, що надається
- 15. Еволюційні (філогенетичні) адаптації - це адаптації, що виникають в ході еволюційного процесу (філогенезу) на основі генетичних
- 16. Існує нескінченна безліч шляхів адаптації рослин. Тим не менш, все адаптації умовно можна розділити лише на
- 17. Пасивна адаптація - «втеча» від шкідливого дії стресора або співіснування з ним. Цей тип адаптації має
- 18. При малому часi дiї фактора у високiй дозi спостерiгаються в основному неспецифiчнi реакцiї - якi виникають
- 20. СОЛЕСТIЙКIСТЬ РОСЛИН За Б.П.Строгоновим грунти за ступенем засоленості подiляються на: 1. практично незасоленi- менше 0,1% солей
- 21. Якісний склад засолення Негативний вплив засолення залежить не тiльки вiд кiлькiсного вмiсту солей, але i вiд
- 22. Тип засолення визначається звичайно за вмiстом анiонiв в грунтi- розрiзняють як основнi типи - хлоридне, сульфатне,
- 23. У розподiлi типiв засолення по територiї бувшого СРСР встановлена така закономiрнiсть - як з пiвночi на
- 24. Токсичний механiзм засолення При засоленнi замiсть амiдiв- глутамiну та аспарагiну- утворюються амiни- путресцин та кадаверин, якi
- 25. Захиснi механiзми рослин. При засоленнi активується синтез органiчних кислот, якi зв'язують надлишковий амiак. Знижується синтез нормальних
- 26. Екологiчнi групи рослин по вiдношенню до засолення Справжнi галофiти-евгалофiти- накопичують солi у вакуолях. Солевидiляючi галофiти- крiногалофiти-
- 28. ЗИМОСТIЙКIСТЬ ХОЛОДОСТIЙКIСТЬ. В першу чергу це поняття вiдноситься до теплолюбних рослин, якi витримують температури від 2-4
- 29. Причини ушкодження від холоду Сакс вважав, що це порушення водообмiну, за Жолкевичем порушення iонного обмiну. Лайонс
- 30. Важливою причиною пошкоджень рослин є перехiд у мембранах насичених жирних кислот з рiдинно-кристалiчного стану в стан
- 31. Загартовування до холоду - низькими i перемiнними температурами, обробка мiкроелементами, ретардантами, солями калiю та аммонiю. Одним
- 32. МОРОЗОСТIЙКIСТЬ Однiєю з перших ознак дiї морозу в рослинах є поява льоду в рослинах. Є три
- 33. ФАКТОРИ морозостiйкостi - у деревних i багаторiчних рослин 1. Своєчасний вхiд в стан спокою, який залежить
- 34. ФАКТОРИ морозостiйкостi 5. Низькi температури не вводять, а виводять рослину iз стану спокою, але до низьких
- 35. IНШI НЕСПРИЯТЛИВI ФАКТОРИ ЗИМИ льодяна кiрка, випирання, випрiвання, вимокання, видування, зимово-весняна посуха, морозобiй у дерев, вiдлиги,
- 37. Автотранспортне навантаження на вул. Івана Франка, 110 (відтінок біля Польського консульства)
- 38. Стiйкiсть до промислових ксенобiотикiв В основному це викиди промисловостi i автотранспорту. Як приклад це стiйкiсть до
- 39. Механізми стійкості до важких металів
- 40. Стiйкiсть рослин до ксенобіотиків обумовлена наявнiстю специфiчних бiлкiв - фiтохелатинiв, якi зв'язують ВМ через групи залишкiв
- 41. Газо, димо, пилостiйкiсть. За фізичними властивостями Викиди у атмосферу, по величинi частинок,електромагнiтному спектру та швидкостi осiдання
- 42. За хiмiчним складом з врахуванням токсичної дiї на рослини викиди можна подiлити на: 1.Кислi гази-найбiльш токсичнi
- 43. Дiя парiв ртутi вiдрiзняеться вiд дii кислих газiв, вуглеводневi сполуки викликають епiнастiю та iншi порушення формотворчих
- 44. Токсичний вплив газiв,пилу та диму на рослини проявляється як шляхом прямої дiї на метаболiчнi та формотворчi
- 45. До бiльш загальних хапактеристик дії викидів можна вiднести такi: 1.Ступiнь пошкодження рослин залежить вiд багатьох факторiв-температури,iнтенсивностi
- 46. СТIЙКIСТЬ ДО ХВОРОБ. Хвороби це бiотичнi фактори - мiкроорганiзми, вiруси- якi отримують поживнi речовини вiд хазяїна.
- 47. Є вертикальна стiйкiсть - коли є стiйкiсть проти одних рас i немає проти iнших (визначається окремими
- 48. Синтез фiтоалексинiв залежить вiд вiку рослин- молодi рослини гороху синтезують бiльше пiзатину, нiж старi. Але синтез
- 49. Захиснi реакцiї у рослин можуть iндукуватись рядом факторiв рiзної природи, якi отримали назву елiситорiв (провокаторiв), якими
- 50. Видовий iмунiтет є майже у всiх рослин. Першим барьером для патогена є покрови та клiтинна стiнка
- 51. На завершальнiй стадiї утворення імунних реакцій синтезуються так званi РR-бiлки, якi є в бiльшостi гiдролiтичними ферментами
- 52. Дефіцит кисню- гіпоксія. При затопленнi та заболочуваннi рослини страждають вiд анаеробiзу- нестачi кисню. Розрiзняють гiпоксiю -
- 53. Крiм того при анаеробiозi протони виходять з вакуолi у цитоплазму i збiльшують її кислотнiсть- це приводить
- 54. В умовах гiпоксiї синтез бiльшостi бiлкiв припиняється, за виключення приблизно 20 полiпептiдiв, якi називають бiлками анаеробного
- 55. Радiостiйкiсть рослин Первиннi механiзми дiї рiзних типiв опромiнення досить подiбнi. Загальна їх особливiсть полягає в тому,
- 56. Основним актом взаємодiї iонiзуючої радiацiї з живим органiзмом є іонізація в результатi якої електрони вiдриваються вiд
- 57. Пiд радiочутливiстю розумiють здатнiсть органiзму реагувати на мiнiмальнi дози радiацiї, якi спричинюють до нелетальних радiобiологiчних ефектiв.
- 58. Слiд вiдрiзняти енергiю, яка падає на бiологiчний об'єкт, вiд енергiї яка поглинається i спричиняє фiзичний вплив.
- 59. Одиниці радіоактивності Так як бiологiчна дiя залежить не тiльки вiд поглинутої дози, але i вiд якостi
- 60. Розрiзняють пряму та опосередковану дiю радiацiї на органiзм. Пряма дiя полягає у перетвореннях молекул в мiсцi
- 61. Згiдно теорiї "мiшеней" - попадання iонiзуючої частинки в чуттєву частину мiшенi( молекула чи структура клiтини) викликає
- 62. Опосередкована дiя полягає в ушкодженнях молекул, мембран та iнших структур живого, якi викликаються продуктами радiолiзу води,
- 63. Замiна кисню любим iншим газом приводить до радiозахисного ефекту. Дiя опромiнення в аеробних умовах (аноксiя) вища,
- 64. Захисна дiя гiпоксiї полягає в тому, що без кисню не утворюються пероксиднi вiльнi радикали, якi посилюють
- 65. Первиннi ушкодження можуть розвиватись: 1) внаслiдок виникнення радiотоксинiв - лiпiдних пероксидiв, хiнонiв, якi приводять до окислення
- 66. два класи радiобiологiчних ефектiв - соматичнi та генетичнi. Соматичнi - це змiни в органiзмi на протязi
- 67. Iз тканин рослин до радiацiї найбiльш чутливi меристеми - причому типовою реакцiєю рослин на опромiнення є
- 68. Можна пiдвищити радiостiйкiсть рослин введенням радiопротекторiв , до яких вiдносяться глутатiон, тiосечовина, цистамiн, цистеїн, цистин, амiноетилтiуронiй
- 69. Посуха— нестача чи відсутність опадів протягом тривалого періоду часу при підвищених температурах та зниженні вологості повітря,
- 70. Click to add title in here Click to add title in here Click to add title
- 71. Циклон Антициклон Опускається з нагрiванням Пiднiмається з охолодженням
- 72. Атмосферна посуха Характеризується високою температурою й низькою відносною вологістю повітря (10-20 %). Її згубна дія полягає
- 73. Грунтова посуха Настає при відсутності опадів та тривалій атмосферній. Спостерігається переважно у другій половині літа, коли
- 74. Вплив посухи може бути прямим i опосередкованим Прямий вплив - при впливi посухи в перiод дiлення
- 75. Ксерофiти подiляються на такі типи: Несукулентні види: а)Справжнi ксерофiти – евксерофiти- мають невеликi листки з великою
- 76. Оберігання їх від дії цього несприятливого фактора Зниження ступеня шкоди від впливу посухи Зниження інтенсивності транспірації
- 78. Скачать презентацию
Слайд 2Види стійкості
Стрес
Солестійкість
Зимостійкість(холодо і морозостійкість та інші несприятливі фактори зими)
Посухостійкість
Стійкість до промислових
Види стійкості
Стрес
Солестійкість
Зимостійкість(холодо і морозостійкість та інші несприятливі фактори зими)
Посухостійкість
Стійкість до промислових

Слайд 3Cтрес - фактор впливу зовнiшнiх умов середовища, який викликає негативний вплив на
Cтрес - фактор впливу зовнiшнiх умов середовища, який викликає негативний вплив на

Слайд 4В дiї на тваринний органiзм i рослини є подiбнiсть i вiдмiнностi -
В дiї на тваринний органiзм i рослини є подiбнiсть i вiдмiнностi -

Слайд 5Специфічні і неспецифічні реакції на стрес
Дiя любого стресу викликає ряд пристосувальних реакцiй.
Специфічні і неспецифічні реакції на стрес
Дiя любого стресу викликає ряд пристосувальних реакцiй.

Слайд 6На дiю бiльшостi стресiв i особливо сильних i при короткому часi дiї
На дiю бiльшостi стресiв i особливо сильних i при короткому часi дiї

Слайд 7За походженням всi екологiчнi фактори подiляють на такi групи:
1.Абiотичнi - клiматичнi, едафiчнi,
За походженням всi екологiчнi фактори подiляють на такi групи:
1.Абiотичнi - клiматичнi, едафiчнi,

Слайд 8За характером дiї стрес як екологiчний фактор може бути:
прямодiючим i опосередкованим (
За характером дiї стрес як екологiчний фактор може бути:
прямодiючим i опосередкованим (

Слайд 9Вплив стресу
Вплив практично всiх стресiв або факторiв можна вкласти в двi кривi
Вплив стресу
Вплив практично всiх стресiв або факторiв можна вкласти в двi кривi

Слайд 10В той же час по вiдношенню до дiї фактора *подразника) рослини можуть
В той же час по вiдношенню до дiї фактора *подразника) рослини можуть

Слайд 11Сукупнiсть екологiчних факторiв називають дiючим середовищем, або середовищем iснування.
Таких середовищ iснування
Сукупнiсть екологiчних факторiв називають дiючим середовищем, або середовищем iснування.
Таких середовищ iснування

Слайд 12Здатність рослини переносити дію несприятливих факторів і давати в таких умовах потомство
Здатність рослини переносити дію несприятливих факторів і давати в таких умовах потомство

Слайд 13Стійкість є кінцевим результатом адаптації.
Адаптація (лат. adaptio - пристосування, прикладання) - це
Стійкість є кінцевим результатом адаптації.
Адаптація (лат. adaptio - пристосування, прикладання) - це

Слайд 14Вибір рослиною стратегії (способу) адаптації залежить від багатьох факторів.
Однак ключовим фактором є
Вибір рослиною стратегії (способу) адаптації залежить від багатьох факторів.
Однак ключовим фактором є

Слайд 15Еволюційні (філогенетичні) адаптації - це адаптації, що виникають в ході еволюційного процесу
Еволюційні (філогенетичні) адаптації - це адаптації, що виникають в ході еволюційного процесу

Слайд 16Існує нескінченна безліч шляхів адаптації рослин. Тим не менш, все адаптації умовно
Існує нескінченна безліч шляхів адаптації рослин. Тим не менш, все адаптації умовно

Слайд 17Пасивна адаптація - «втеча» від шкідливого дії стресора або співіснування з ним.
Пасивна адаптація - «втеча» від шкідливого дії стресора або співіснування з ним.

Слайд 18При малому часi дiї фактора у високiй дозi спостерiгаються в основному неспецифiчнi
При малому часi дiї фактора у високiй дозi спостерiгаються в основному неспецифiчнi


Слайд 20СОЛЕСТIЙКIСТЬ РОСЛИН
За Б.П.Строгоновим грунти за ступенем засоленості подiляються на:
1. практично незасоленi- менше
СОЛЕСТIЙКIСТЬ РОСЛИН
За Б.П.Строгоновим грунти за ступенем засоленості подiляються на:
1. практично незасоленi- менше

Слайд 21Якісний склад засолення
Негативний вплив засолення залежить не тiльки вiд кiлькiсного вмiсту солей,
Якісний склад засолення
Негативний вплив засолення залежить не тiльки вiд кiлькiсного вмiсту солей,

Слайд 22Тип засолення
визначається звичайно за вмiстом анiонiв в грунтi- розрiзняють як основнi типи
Тип засолення
визначається звичайно за вмiстом анiонiв в грунтi- розрiзняють як основнi типи

Слайд 23У розподiлi типiв засолення по територiї бувшого СРСР встановлена така закономiрнiсть
- як
У розподiлi типiв засолення по територiї бувшого СРСР встановлена така закономiрнiсть
- як

Слайд 24Токсичний механiзм засолення
При засоленнi замiсть амiдiв- глутамiну та аспарагiну- утворюються амiни- путресцин
Токсичний механiзм засолення
При засоленнi замiсть амiдiв- глутамiну та аспарагiну- утворюються амiни- путресцин

Слайд 25Захиснi механiзми рослин.
При засоленнi активується синтез органiчних кислот, якi зв'язують надлишковий
Захиснi механiзми рослин.
При засоленнi активується синтез органiчних кислот, якi зв'язують надлишковий

Слайд 26Екологiчнi групи рослин по вiдношенню до засолення
Справжнi галофiти-евгалофiти- накопичують солi у
Екологiчнi групи рослин по вiдношенню до засолення
Справжнi галофiти-евгалофiти- накопичують солi у


Слайд 28ЗИМОСТIЙКIСТЬ
ХОЛОДОСТIЙКIСТЬ.
В першу чергу це поняття вiдноситься до теплолюбних рослин, якi витримують
ЗИМОСТIЙКIСТЬ
ХОЛОДОСТIЙКIСТЬ.
В першу чергу це поняття вiдноситься до теплолюбних рослин, якi витримують

Слайд 29Причини ушкодження від холоду
Сакс вважав, що це порушення водообмiну, за Жолкевичем порушення
Причини ушкодження від холоду
Сакс вважав, що це порушення водообмiну, за Жолкевичем порушення

Слайд 30Важливою причиною пошкоджень рослин
є перехiд у мембранах насичених жирних кислот з рiдинно-кристалiчного
Важливою причиною пошкоджень рослин
є перехiд у мембранах насичених жирних кислот з рiдинно-кристалiчного

Слайд 31Загартовування до холоду
- низькими i перемiнними температурами, обробка мiкроелементами, ретардантами, солями калiю
Загартовування до холоду
- низькими i перемiнними температурами, обробка мiкроелементами, ретардантами, солями калiю

Слайд 32МОРОЗОСТIЙКIСТЬ
Однiєю з перших ознак дiї морозу в рослинах є поява льоду в
МОРОЗОСТIЙКIСТЬ
Однiєю з перших ознак дiї морозу в рослинах є поява льоду в

Слайд 33ФАКТОРИ морозостiйкостi - у деревних i багаторiчних рослин
1. Своєчасний вхiд в стан
ФАКТОРИ морозостiйкостi - у деревних i багаторiчних рослин
1. Своєчасний вхiд в стан

Слайд 34ФАКТОРИ морозостiйкостi
5. Низькi температури не вводять, а виводять рослину iз стану спокою,
ФАКТОРИ морозостiйкостi
5. Низькi температури не вводять, а виводять рослину iз стану спокою,

Слайд 35IНШI НЕСПРИЯТЛИВI ФАКТОРИ ЗИМИ
льодяна кiрка,
випирання,
випрiвання,
вимокання,
видування,
зимово-весняна посуха, морозобiй
IНШI НЕСПРИЯТЛИВI ФАКТОРИ ЗИМИ
льодяна кiрка,
випирання,
випрiвання,
вимокання,
видування,
зимово-весняна посуха, морозобiй


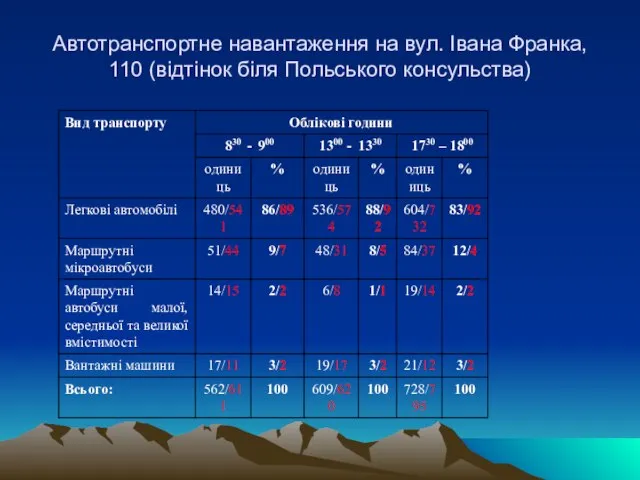
Слайд 37Автотранспортне навантаження на вул. Івана Франка, 110 (відтінок біля Польського консульства)
Автотранспортне навантаження на вул. Івана Франка, 110 (відтінок біля Польського консульства)
Слайд 38Стiйкiсть до промислових ксенобiотикiв
В основному це викиди промисловостi i автотранспорту. Як приклад
Стiйкiсть до промислових ксенобiотикiв
В основному це викиди промисловостi i автотранспорту. Як приклад

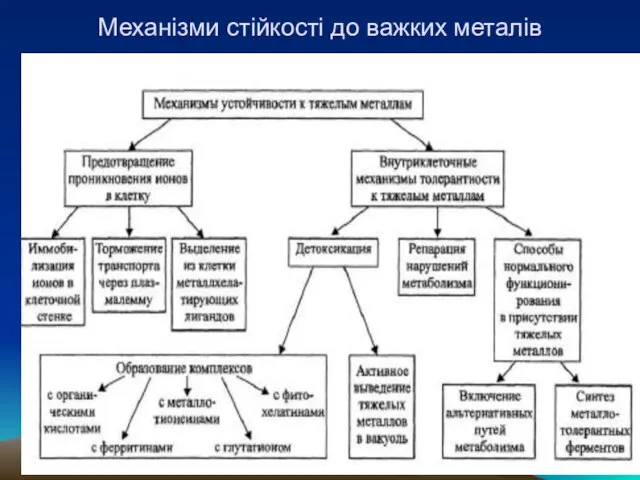
Слайд 39Механізми стійкості до важких металів
Механізми стійкості до важких металів
Слайд 40Стiйкiсть рослин до ксенобіотиків
обумовлена наявнiстю специфiчних бiлкiв - фiтохелатинiв, якi зв'язують
Стiйкiсть рослин до ксенобіотиків
обумовлена наявнiстю специфiчних бiлкiв - фiтохелатинiв, якi зв'язують

Слайд 41Газо, димо, пилостiйкiсть.
За фізичними властивостями Викиди у атмосферу, по величинi частинок,електромагнiтному спектру
Газо, димо, пилостiйкiсть.
За фізичними властивостями Викиди у атмосферу, по величинi частинок,електромагнiтному спектру

Слайд 42За хiмiчним складом з врахуванням токсичної дiї на рослини викиди можна подiлити
За хiмiчним складом з врахуванням токсичної дiї на рослини викиди можна подiлити

Слайд 43Дiя парiв ртутi вiдрiзняеться вiд дii кислих газiв,
вуглеводневi сполуки викликають епiнастiю та
Дiя парiв ртутi вiдрiзняеться вiд дii кислих газiв,
вуглеводневi сполуки викликають епiнастiю та

Слайд 44Токсичний вплив газiв,пилу та диму
на рослини проявляється як шляхом прямої дiї на
Токсичний вплив газiв,пилу та диму
на рослини проявляється як шляхом прямої дiї на

Слайд 45До бiльш загальних хапактеристик дії викидів можна вiднести такi:
1.Ступiнь пошкодження рослин залежить
До бiльш загальних хапактеристик дії викидів можна вiднести такi:
1.Ступiнь пошкодження рослин залежить

Слайд 46СТIЙКIСТЬ ДО ХВОРОБ.
Хвороби це бiотичнi фактори - мiкроорганiзми, вiруси- якi отримують поживнi
СТIЙКIСТЬ ДО ХВОРОБ.
Хвороби це бiотичнi фактори - мiкроорганiзми, вiруси- якi отримують поживнi

Слайд 47Є вертикальна стiйкiсть - коли є стiйкiсть проти одних рас i немає
Є вертикальна стiйкiсть - коли є стiйкiсть проти одних рас i немає

Слайд 48Синтез фiтоалексинiв залежить вiд вiку рослин- молодi рослини гороху синтезують бiльше пiзатину,
Синтез фiтоалексинiв залежить вiд вiку рослин- молодi рослини гороху синтезують бiльше пiзатину,

Слайд 49Захиснi реакцiї у рослин
можуть iндукуватись рядом факторiв рiзної природи, якi отримали назву
Захиснi реакцiї у рослин
можуть iндукуватись рядом факторiв рiзної природи, якi отримали назву

Слайд 50Видовий iмунiтет
є майже у всiх рослин.
Першим барьером для патогена є покрови
Видовий iмунiтет
є майже у всiх рослин.
Першим барьером для патогена є покрови

Слайд 51На завершальнiй стадiї утворення імунних реакцій
синтезуються так званi РR-бiлки, якi є в
На завершальнiй стадiї утворення імунних реакцій
синтезуються так званi РR-бiлки, якi є в

Слайд 52Дефіцит кисню- гіпоксія.
При затопленнi та заболочуваннi рослини страждають вiд анаеробiзу- нестачi кисню.
Дефіцит кисню- гіпоксія.
При затопленнi та заболочуваннi рослини страждають вiд анаеробiзу- нестачi кисню.

Слайд 53Крiм того при анаеробiозi протони виходять з вакуолi у цитоплазму i збiльшують
Крiм того при анаеробiозi протони виходять з вакуолi у цитоплазму i збiльшують

Слайд 54В умовах гiпоксiї
синтез бiльшостi бiлкiв припиняється, за виключення приблизно 20 полiпептiдiв, якi
В умовах гiпоксiї
синтез бiльшостi бiлкiв припиняється, за виключення приблизно 20 полiпептiдiв, якi

Слайд 55Радiостiйкiсть рослин
Первиннi механiзми дiї рiзних типiв опромiнення досить подiбнi. Загальна їх особливiсть
Радiостiйкiсть рослин
Первиннi механiзми дiї рiзних типiв опромiнення досить подiбнi. Загальна їх особливiсть

Слайд 56Основним актом взаємодiї iонiзуючої радiацiї з живим органiзмом є іонізація
в результатi
Основним актом взаємодiї iонiзуючої радiацiї з живим органiзмом є іонізація
в результатi

Слайд 57Пiд радiочутливiстю розумiють здатнiсть органiзму реагувати на мiнiмальнi дози радiацiї, якi спричинюють
Пiд радiочутливiстю розумiють здатнiсть органiзму реагувати на мiнiмальнi дози радiацiї, якi спричинюють

Слайд 58Слiд вiдрiзняти енергiю, яка падає на бiологiчний об'єкт, вiд енергiї яка поглинається
Слiд вiдрiзняти енергiю, яка падає на бiологiчний об'єкт, вiд енергiї яка поглинається

Слайд 59Одиниці радіоактивності
Так як бiологiчна дiя залежить не тiльки вiд поглинутої дози, але
Одиниці радіоактивності
Так як бiологiчна дiя залежить не тiльки вiд поглинутої дози, але

Слайд 60Розрiзняють пряму та опосередковану дiю радiацiї на органiзм.
Пряма дiя полягає у перетвореннях
Розрiзняють пряму та опосередковану дiю радiацiї на органiзм.
Пряма дiя полягає у перетвореннях

Слайд 61Згiдно теорiї "мiшеней" - попадання iонiзуючої частинки в чуттєву частину мiшенi( молекула
Згiдно теорiї "мiшеней" - попадання iонiзуючої частинки в чуттєву частину мiшенi( молекула

Слайд 62Опосередкована дiя
полягає в ушкодженнях молекул, мембран та iнших структур живого, якi викликаються
Опосередкована дiя
полягає в ушкодженнях молекул, мембран та iнших структур живого, якi викликаються

Слайд 63Замiна кисню любим iншим газом приводить до радiозахисного ефекту.
Дiя опромiнення в аеробних
Замiна кисню любим iншим газом приводить до радiозахисного ефекту.
Дiя опромiнення в аеробних

Слайд 64Захисна дiя гiпоксiї полягає в тому, що без кисню не утворюються пероксиднi
Захисна дiя гiпоксiї полягає в тому, що без кисню не утворюються пероксиднi

Слайд 65Первиннi ушкодження можуть розвиватись:
1) внаслiдок виникнення радiотоксинiв - лiпiдних пероксидiв, хiнонiв, якi
Первиннi ушкодження можуть розвиватись:
1) внаслiдок виникнення радiотоксинiв - лiпiдних пероксидiв, хiнонiв, якi

Слайд 66два класи радiобiологiчних ефектiв - соматичнi та генетичнi.
Соматичнi - це змiни
два класи радiобiологiчних ефектiв - соматичнi та генетичнi.
Соматичнi - це змiни

Слайд 67Iз тканин рослин до радiацiї найбiльш чутливi меристеми
- причому типовою реакцiєю рослин
Iз тканин рослин до радiацiї найбiльш чутливi меристеми
- причому типовою реакцiєю рослин

Слайд 68Можна пiдвищити радiостiйкiсть рослин введенням радiопротекторiв
, до яких вiдносяться глутатiон, тiосечовина, цистамiн,
Можна пiдвищити радiостiйкiсть рослин введенням радiопротекторiв
, до яких вiдносяться глутатiон, тiосечовина, цистамiн,

Слайд 69Посуха— нестача чи відсутність опадів протягом тривалого періоду часу при підвищених температурах
Посуха— нестача чи відсутність опадів протягом тривалого періоду часу при підвищених температурах

Слайд 70Click to add title in here
Click to add title in here
Click to add title in here
Click to add title in here

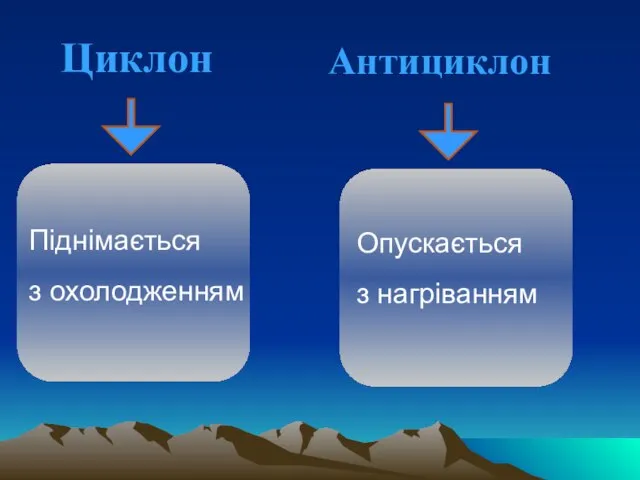
Слайд 71Циклон
Антициклон
Опускається
з нагрiванням
Пiднiмається
з охолодженням
Циклон
Антициклон
Опускається
з нагрiванням
Пiднiмається
з охолодженням
Слайд 72Атмосферна посуха
Характеризується високою температурою й низькою відносною вологістю повітря (10-20 %). Її
Атмосферна посуха
Характеризується високою температурою й низькою відносною вологістю повітря (10-20 %). Її

Слайд 73Грунтова посуха
Настає при відсутності опадів та тривалій атмосферній. Спостерігається переважно у другій
Грунтова посуха
Настає при відсутності опадів та тривалій атмосферній. Спостерігається переважно у другій

Слайд 74Вплив посухи може бути прямим i опосередкованим
Прямий вплив - при впливi
Вплив посухи може бути прямим i опосередкованим
Прямий вплив - при впливi

Слайд 75Ксерофiти подiляються на такі типи:
Несукулентні види:
а)Справжнi ксерофiти –
евксерофiти- мають невеликi
листки з
Ксерофiти подiляються на такі типи:
Несукулентні види:
а)Справжнi ксерофiти –
евксерофiти- мають невеликi
листки з

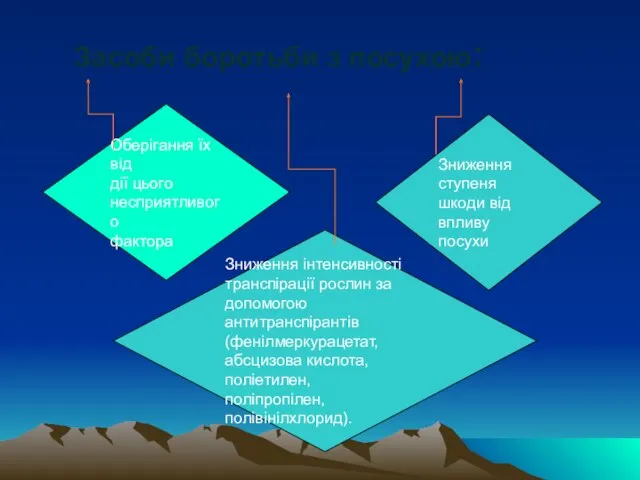
Слайд 76Оберігання їх від
дії цього
несприятливого
фактора
Зниження ступеня
шкоди від впливу
Оберігання їх від
дії цього
несприятливого
фактора
Зниження ступеня
шкоди від впливу
 тематичний словник
тематичний словник Современная модель системы высшего образования
Современная модель системы высшего образования Навчання дітей елементів грамоти
Навчання дітей елементів грамоти Унылая пора! Очей очарованье!
Унылая пора! Очей очарованье! Ancient Egyptian Hieroglyph Template
Ancient Egyptian Hieroglyph Template Презентация_5786
Презентация_5786 Презентация.Создание презентации.
Презентация.Создание презентации. Развитие методов стратегического анализа. Разработка моделей 6х5 и 10х5
Развитие методов стратегического анализа. Разработка моделей 6х5 и 10х5 Электронный журнал – помощник в составлении аналитической документации
Электронный журнал – помощник в составлении аналитической документации Государственные символы России
Государственные символы России Обоснование выбора компетенции
Обоснование выбора компетенции Портфолио. Латыпова Галия Мансуровна
Портфолио. Латыпова Галия Мансуровна Презентация на тему Звери запасаются жиром
Презентация на тему Звери запасаются жиром Презентация на тему Влияние на жизнь и здоровье людей природных катастроф
Презентация на тему Влияние на жизнь и здоровье людей природных катастроф «…И образ незабвенный» Урок разработала и провела: Хрящикова Людмила Николаевна, учитель русского языка и литературы
«…И образ незабвенный» Урок разработала и провела: Хрящикова Людмила Николаевна, учитель русского языка и литературы Скульптура и МДК. Отчет по проделанной работе
Скульптура и МДК. Отчет по проделанной работе Силы, действующие на ракету в плотных слоях атмосферы
Силы, действующие на ракету в плотных слоях атмосферы Михаил Евграфович Салтыков-Щедрин (1826 - 1889)
Михаил Евграфович Салтыков-Щедрин (1826 - 1889) Об организации приема в образовательные учреждения среднего профессионального и начального профессионального образования на 2012-
Об организации приема в образовательные учреждения среднего профессионального и начального профессионального образования на 2012- Тема 2 Экономические системы и институты
Тема 2 Экономические системы и институты Выборы - 2007 в местные органы управления часть 2. Политические партии
Выборы - 2007 в местные органы управления часть 2. Политические партии Голография
Голография Модели объектов и процессов
Модели объектов и процессов Интерактивный тест: ДРЕВНИЙ ЕГИПЕТ «Найди на рисунке…»
Интерактивный тест: ДРЕВНИЙ ЕГИПЕТ «Найди на рисунке…» Внешняя политика. Политика односторонних уступок Западу
Внешняя политика. Политика односторонних уступок Западу Система работы классного руководителяпо организации совместной деятельности школы и семьи через создание центров в классном к
Система работы классного руководителяпо организации совместной деятельности школы и семьи через создание центров в классном к Презентация на тему Отношения в обществе
Презентация на тему Отношения в обществе АГТ – Уралкоммуникационное агентство
АГТ – Уралкоммуникационное агентство