- Биохимияға кіріспе. Нәруыздардың құрылысы мен қызметтері

Содержание
- 2. Дәріс жоспары: 1. Биохимияға кіріспе. 2. Протеиногендік амин қышқылдарының құрылысы мен жіктелуі. 3. Нәруыздардың бірінші, екінші,
- 3. КІРІСПЕ Биологиялық химия, тіршілік туралы ғылым, яғни организмнің құрамына кіретін заттардың құрылысын, биологиялық қызметін, олардың алмасуын
- 5. 8 8
- 6. 7 7
- 10. Ақуыздардың екінші реттік құрылысы Екінші реттік құрылыстың үш түрі болады: α-серіппелі, β-құрылымды (қатпарлы қабатты) және ретсіз
- 12. Толық-β-құрылым (қатпарлы қабат) бірнеше полипептидтік тізбектерінің карбонил және имин топтарының арасында пайда болған сутектік байланыстардың нәтижесінде
- 13. Толық-β - құрылымындағы полипептидтік тізбектер бір біріне паралелльді орналасып, парақ қағазды қатарлап бүкпелеп түзген фигураға ұқсас
- 14. Қысқа-β-құрылым, бір полипептидтік тізбектің бойында α-серіппелі және ретсіз құрылымдармен кезектесіп орналасады. Бір полипептидтік тізбек өзі өзіне
- 15. Ақуыздардың үшінші реттік құрылысы Екінші реттік құрылымды полипептидтік тізбектің кеңістікте белгілі көлемде орналасуы нәтижесінде түзілген құрылымды
- 19. Ақуыздардың төртінші реттік құрылысы Екі және одан да көп полипептидтік тізбектен тұратын ақуыздар үшін төртінші реттік
- 20. Олигомерлік ақуыздарга темекі мозайкалық вирусы (РНҚ-ның бір молекуласынан және бір текті ақуыздың 2130 суббөлігінен тұратын), фосфорилаза
- 21. Нәруыздардың суперекіншілік құрылымы Әрбір нәруыздар кеңістік кұрылысы ерекше және ол біріншілік құрылыммен анықталады. Дегенмен құрылымы және
- 22. α/β -Бөшкелік суперекіншілік құрылым Мұндай құрылым, шынында да бөшке тәрізді болып келуі, яғни әрбір β-құрылым (1.16-суретте
- 23. «α-Спираль – бұрылыс – α -спираль» құрылымдық мотив Мұндай «құрылымдык мотив» көптеген ДНҚ- байланыстырушы нәруыздарда кездеседі.
- 24. «Мырышты саусақ түріндегі суперекіншілік құрылым» Суперекіншілік құрылымның бұл түрі де ДНҚ-ны байланыстырушы нәруыздарда жиі кездеседі. «Мырышты
- 25. 12 амин қышқылдарының қалдықтарына амин қышқылдарының орналасу ретінде біріне-бірі жақын орналасқан екі цистеин қалдығы, екі гистидин
- 26. Лейцин қалдықтары әрбір 6 амин қышқылдарынан кейін қайталанып орналасады. Әрбір α-спиральдың бір орамында 3,6 амин қышқылы
- 27. «Лейцинді тістесіп-бірігу» арқылы нәруыздардың қосылуын гистондарды мысалға алып қарастыруға болады. Гистондарядролық нәруыздар, олардың құрамында оң зарядталған
- 28. НӘРУЫЗДАРДЫҢ ДОМЕНДІ ҚҰРЫЛЫМЫ Егер нәруызда полипептидтік тізбек 200-ден астам амин қышқылдары қалдықтарынан тұратын болса, оның кеңістіктегі
- 29. ДОМЕНДІК ҚҰРЫЛЫМДЫ АҚУЫЗДАР Домендік ақуыздардың құрылысы олигомерлік ақуыздардың құрылысына ұқсас. Бірақта олигомерлік ақуыздардың әрбір полипептидтік тізбегінен
- 30. Жеңіл полипептидтік тізбектері (200 а.қ. тұратын) екі домендік, ал ауыр полипептидтік тізбектері (450-700 а.қ. тұратын) төрт
- 32. Дәріс мазмұны бойынша кері байланыс сұрақтары: 1. Нәруыздардың бірінші, екінші, үшінші реттік құрылыстарына түсініктеме беріңіз. 2.
- 34. Скачать презентацию
Слайд 2Дәріс жоспары:
1. Биохимияға кіріспе.
2. Протеиногендік амин қышқылдарының құрылысы мен жіктелуі.
3. Нәруыздардың бірінші,
Дәріс жоспары:
1. Биохимияға кіріспе.
2. Протеиногендік амин қышқылдарының құрылысы мен жіктелуі.
3. Нәруыздардың бірінші,

Слайд 3КІРІСПЕ
Биологиялық химия, тіршілік туралы ғылым, яғни организмнің құрамына кіретін заттардың құрылысын, биологиялық
Биологиялық химия, тіршілік туралы ғылым, яғни организмнің құрамына кіретін заттардың құрылысын, биологиялық

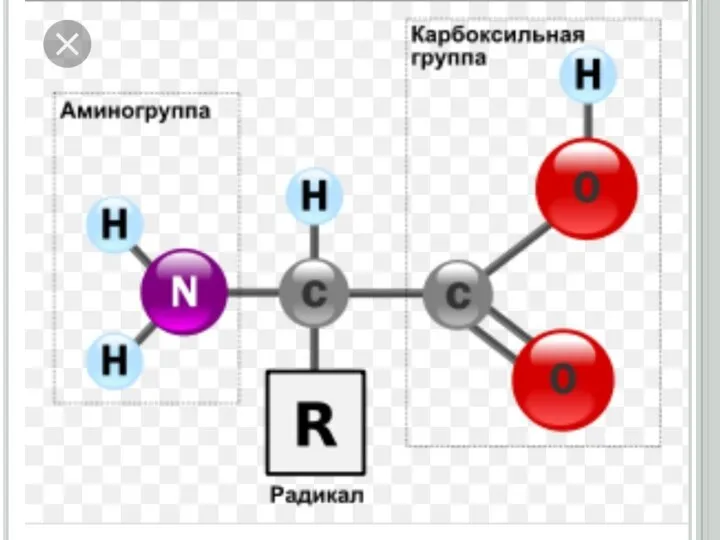
Слайд 58
8
8
8
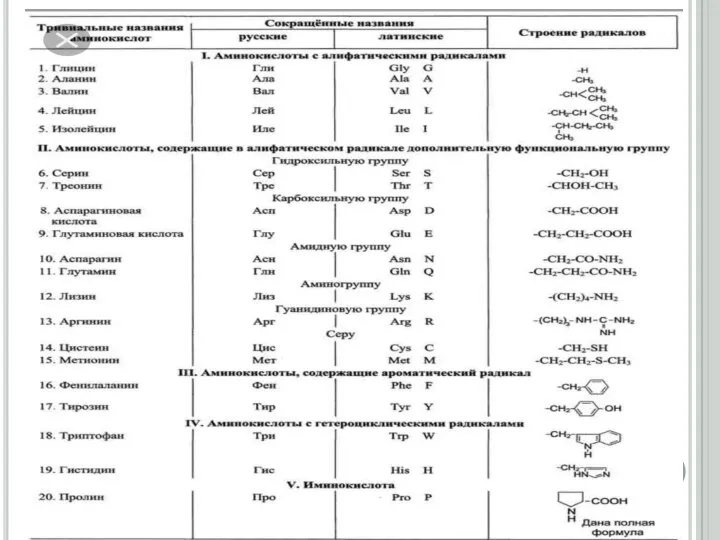
Слайд 67
7
7
7
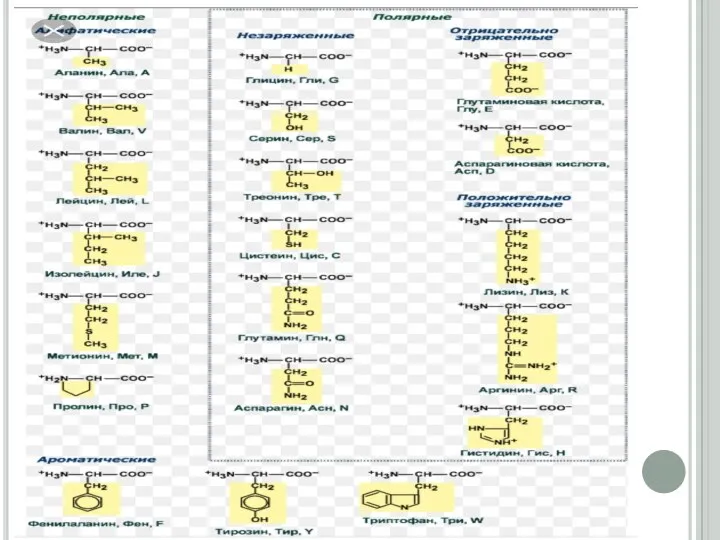
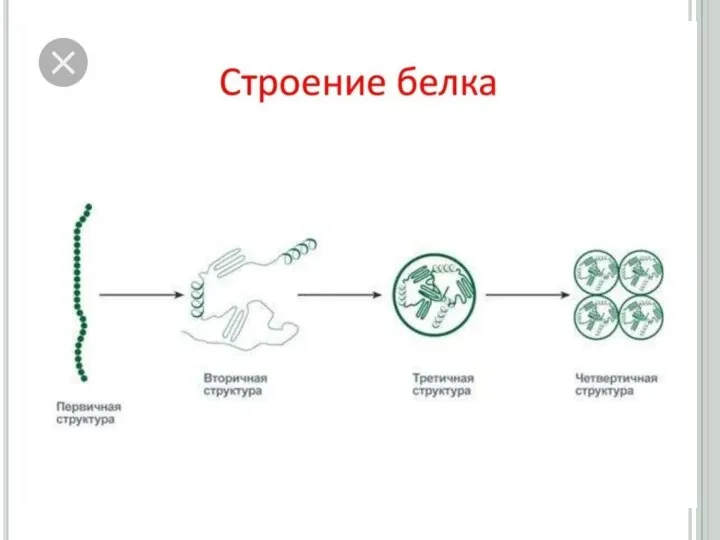
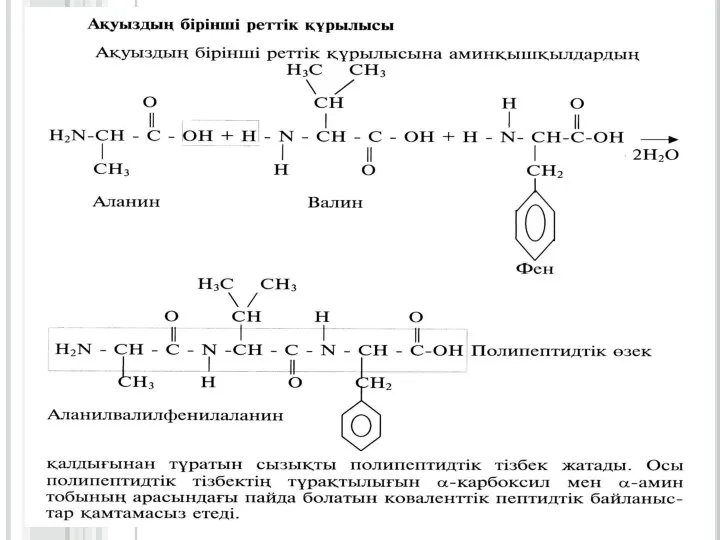
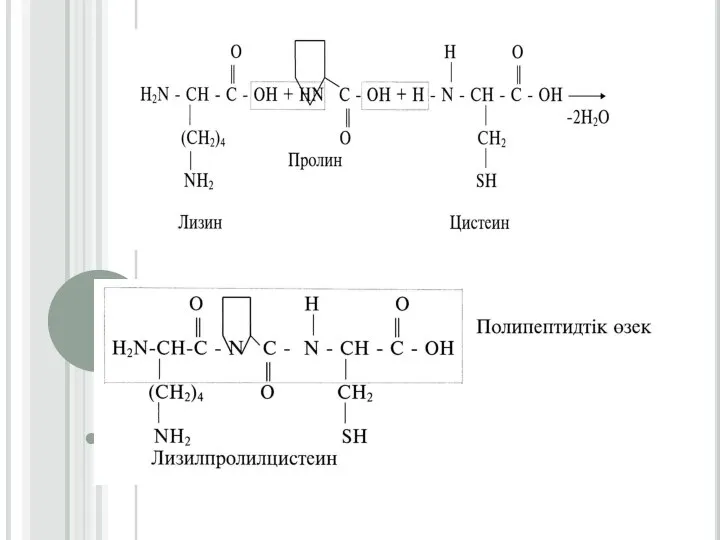
Слайд 10Ақуыздардың екінші реттік құрылысы
Екінші реттік құрылыстың үш түрі болады: α-серіппелі, β-құрылымды (қатпарлы
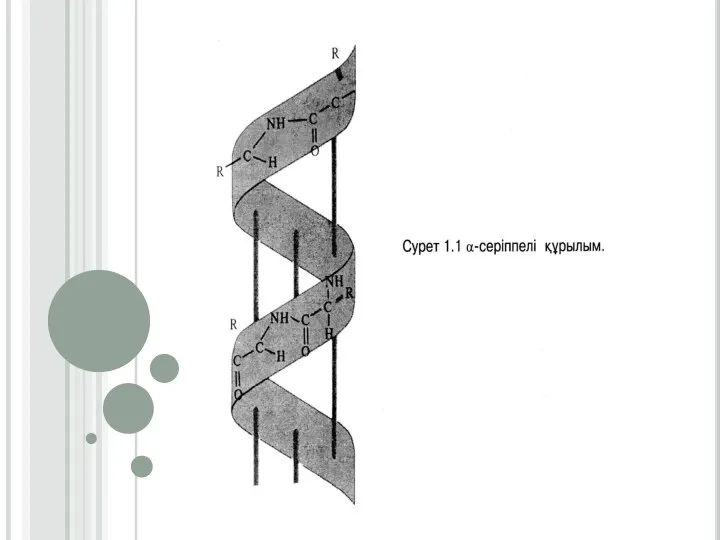
Ақуыздардың екінші реттік құрылысы
Екінші реттік құрылыстың үш түрі болады: α-серіппелі, β-құрылымды (қатпарлы

Слайд 12Толық-β-құрылым (қатпарлы қабат) бірнеше полипептидтік тізбектерінің карбонил және имин топтарының арасында пайда
Толық-β-құрылым (қатпарлы қабат) бірнеше полипептидтік тізбектерінің карбонил және имин топтарының арасында пайда

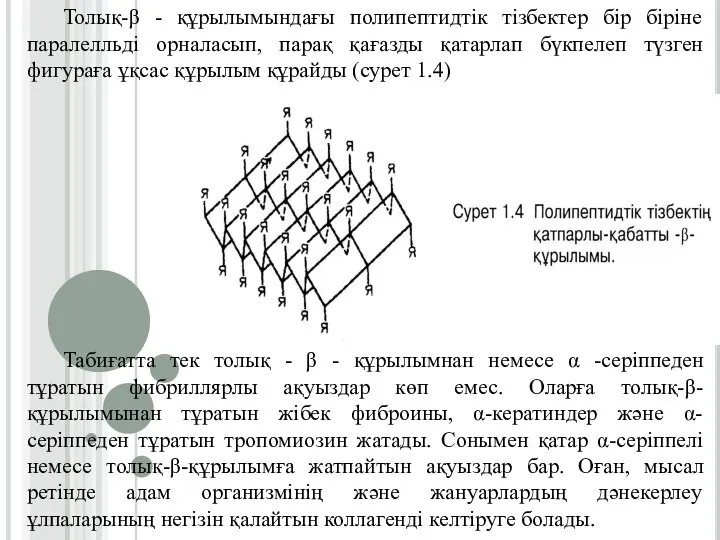
Слайд 13Толық-β - құрылымындағы полипептидтік тізбектер бір біріне паралелльді орналасып, парақ қағазды қатарлап
Толық-β - құрылымындағы полипептидтік тізбектер бір біріне паралелльді орналасып, парақ қағазды қатарлап
Слайд 14Қысқа-β-құрылым, бір полипептидтік тізбектің бойында α-серіппелі және ретсіз құрылымдармен кезектесіп орналасады. Бір
Қысқа-β-құрылым, бір полипептидтік тізбектің бойында α-серіппелі және ретсіз құрылымдармен кезектесіп орналасады. Бір

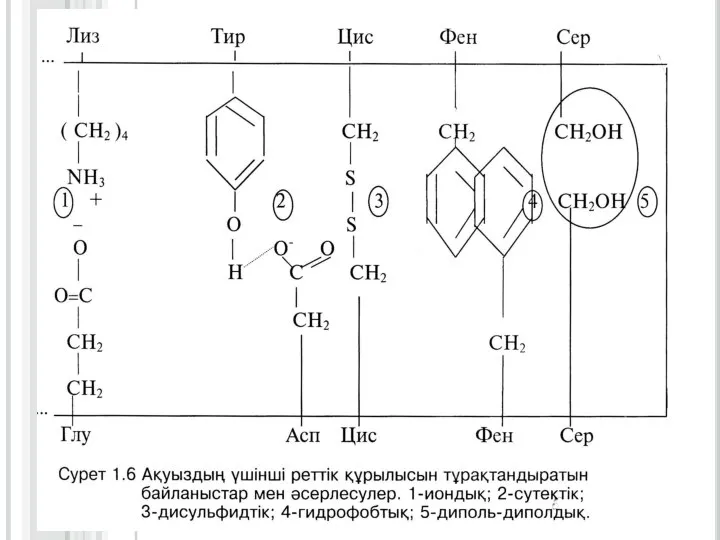
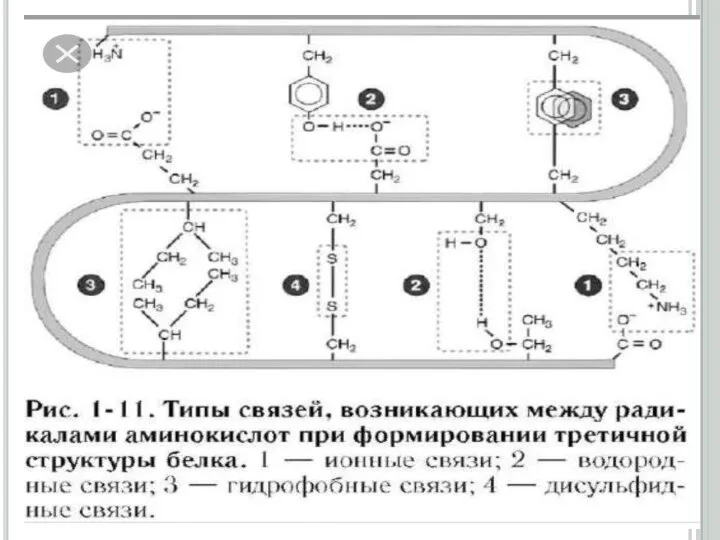
Слайд 15Ақуыздардың үшінші реттік құрылысы
Екінші реттік құрылымды полипептидтік тізбектің кеңістікте белгілі көлемде орналасуы
Ақуыздардың үшінші реттік құрылысы
Екінші реттік құрылымды полипептидтік тізбектің кеңістікте белгілі көлемде орналасуы

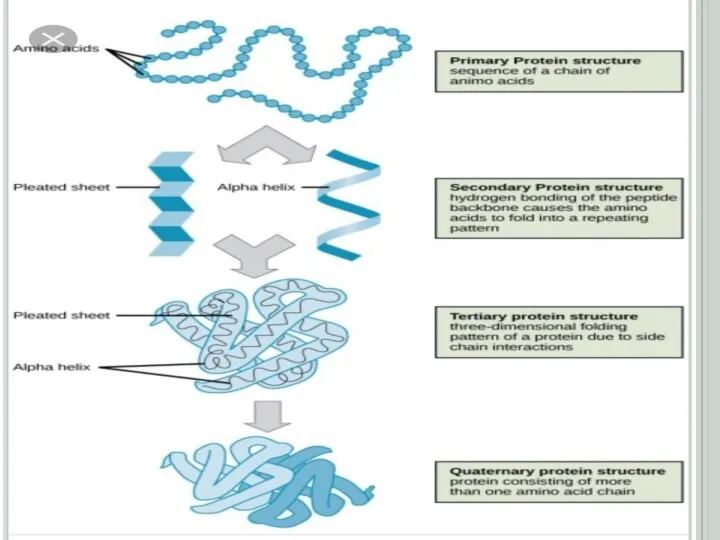
Слайд 19Ақуыздардың төртінші реттік құрылысы
Екі және одан да көп полипептидтік тізбектен тұратын ақуыздар
Ақуыздардың төртінші реттік құрылысы
Екі және одан да көп полипептидтік тізбектен тұратын ақуыздар

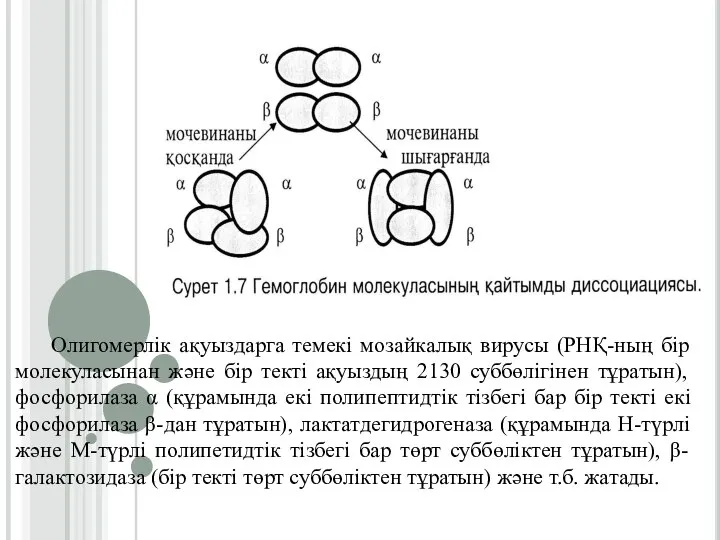
Слайд 20Олигомерлік ақуыздарга темекі мозайкалық вирусы (РНҚ-ның бір молекуласынан және бір текті ақуыздың
Олигомерлік ақуыздарга темекі мозайкалық вирусы (РНҚ-ның бір молекуласынан және бір текті ақуыздың
Слайд 21Нәруыздардың суперекіншілік құрылымы
Әрбір нәруыздар кеңістік кұрылысы ерекше және ол біріншілік құрылыммен анықталады.
Нәруыздардың суперекіншілік құрылымы
Әрбір нәруыздар кеңістік кұрылысы ерекше және ол біріншілік құрылыммен анықталады.

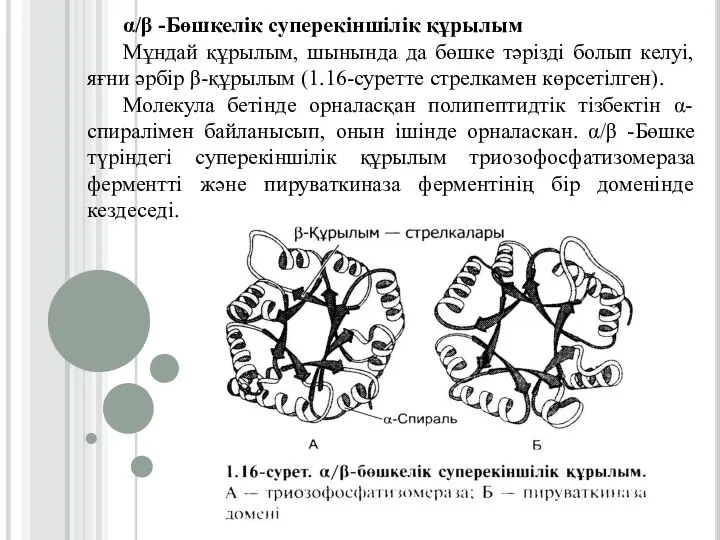
Слайд 22α/β -Бөшкелік суперекіншілік құрылым
Мұндай құрылым, шынында да бөшке тәрізді болып келуі, яғни
α/β -Бөшкелік суперекіншілік құрылым
Мұндай құрылым, шынында да бөшке тәрізді болып келуі, яғни
Слайд 23«α-Спираль – бұрылыс – α -спираль» құрылымдық мотив
Мұндай «құрылымдык мотив» көптеген ДНҚ-
«α-Спираль – бұрылыс – α -спираль» құрылымдық мотив
Мұндай «құрылымдык мотив» көптеген ДНҚ-

Слайд 24«Мырышты саусақ түріндегі суперекіншілік құрылым»
Суперекіншілік құрылымның бұл түрі де ДНҚ-ны байланыстырушы нәруыздарда
«Мырышты саусақ түріндегі суперекіншілік құрылым»
Суперекіншілік құрылымның бұл түрі де ДНҚ-ны байланыстырушы нәруыздарда
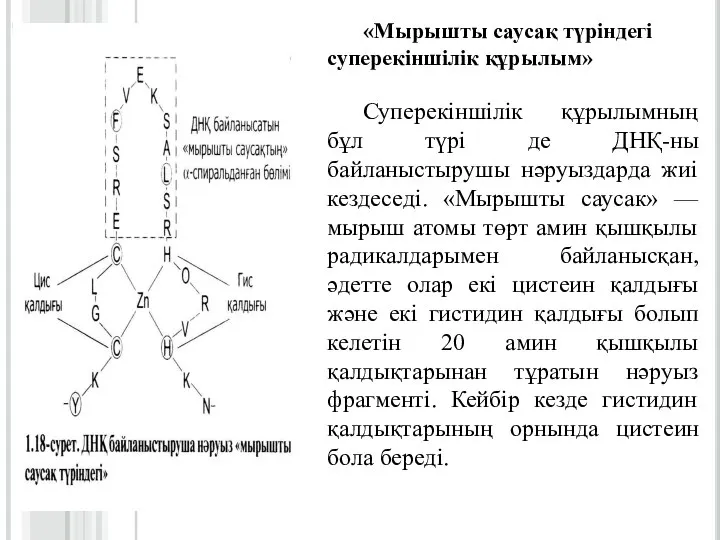
Слайд 2512 амин қышқылдарының қалдықтарына амин қышқылдарының орналасу ретінде біріне-бірі жақын орналасқан екі

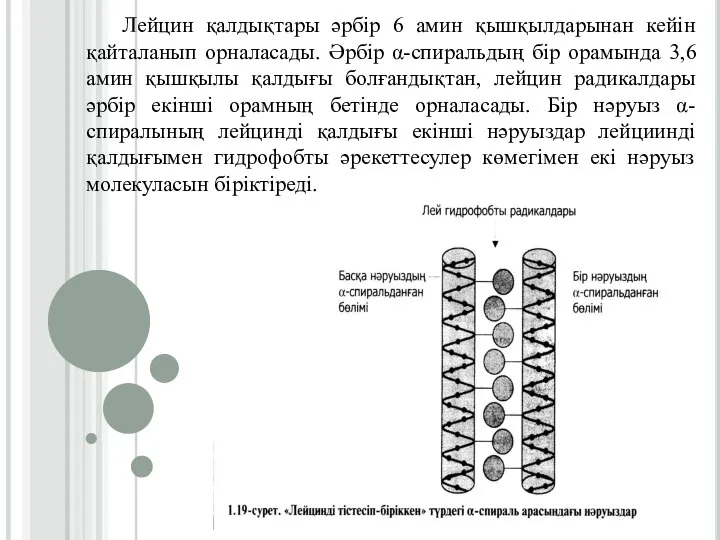
Слайд 26Лейцин қалдықтары әрбір 6 амин қышқылдарынан кейін қайталанып орналасады. Әрбір α-спиральдың бір
Лейцин қалдықтары әрбір 6 амин қышқылдарынан кейін қайталанып орналасады. Әрбір α-спиральдың бір
Слайд 27«Лейцинді тістесіп-бірігу» арқылы нәруыздардың қосылуын гистондарды мысалға алып қарастыруға болады. Гистондарядролық нәруыздар,
«Лейцинді тістесіп-бірігу» арқылы нәруыздардың қосылуын гистондарды мысалға алып қарастыруға болады. Гистондарядролық нәруыздар,

Слайд 28НӘРУЫЗДАРДЫҢ ДОМЕНДІ ҚҰРЫЛЫМЫ
Егер нәруызда полипептидтік тізбек 200-ден астам амин қышқылдары қалдықтарынан тұратын
НӘРУЫЗДАРДЫҢ ДОМЕНДІ ҚҰРЫЛЫМЫ
Егер нәруызда полипептидтік тізбек 200-ден астам амин қышқылдары қалдықтарынан тұратын

Слайд 29ДОМЕНДІК ҚҰРЫЛЫМДЫ АҚУЫЗДАР
Домендік ақуыздардың құрылысы олигомерлік ақуыздардың құрылысына ұқсас. Бірақта олигомерлік ақуыздардың
ДОМЕНДІК ҚҰРЫЛЫМДЫ АҚУЫЗДАР
Домендік ақуыздардың құрылысы олигомерлік ақуыздардың құрылысына ұқсас. Бірақта олигомерлік ақуыздардың

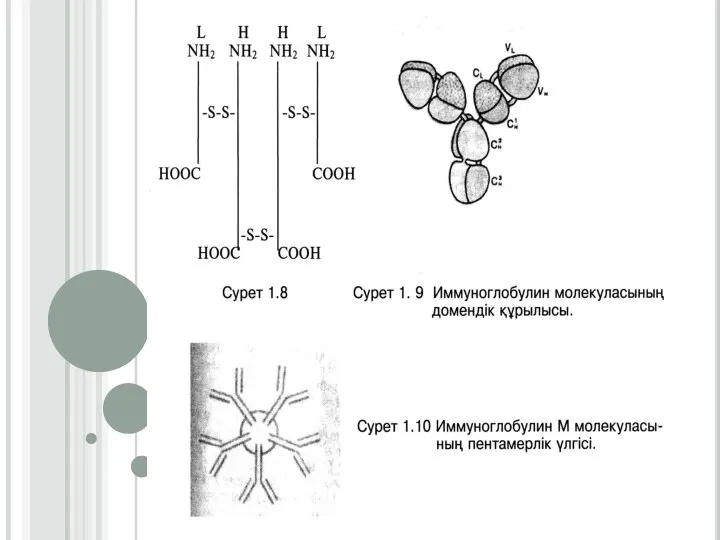
Слайд 30Жеңіл полипептидтік тізбектері (200 а.қ. тұратын) екі домендік, ал ауыр полипептидтік тізбектері
Жеңіл полипептидтік тізбектері (200 а.қ. тұратын) екі домендік, ал ауыр полипептидтік тізбектері

Слайд 32Дәріс мазмұны бойынша кері байланыс сұрақтары:
1. Нәруыздардың бірінші, екінші, үшінші реттік құрылыстарына
Дәріс мазмұны бойынша кері байланыс сұрақтары:
1. Нәруыздардың бірінші, екінші, үшінші реттік құрылыстарына

 Стоп ВИЧ и СПИД
Стоп ВИЧ и СПИД Клинический случай
Клинический случай Иммунопрофилактика. Информация к размышлению…
Иммунопрофилактика. Информация к размышлению… Дезинфекция
Дезинфекция Сестринский процесс при заболеваниях печени и жёлчевыводящей системы
Сестринский процесс при заболеваниях печени и жёлчевыводящей системы Доплаты медицинским работникам, работающим с COVID-19, Вологодская область
Доплаты медицинским работникам, работающим с COVID-19, Вологодская область Гигиена питания
Гигиена питания Oral Ferric Maltol for the Treatment of Iron-Deficiency Anemia in Patients With CKD
Oral Ferric Maltol for the Treatment of Iron-Deficiency Anemia in Patients With CKD Техніка накладання найбільш поширених пов’язок
Техніка накладання найбільш поширених пов’язок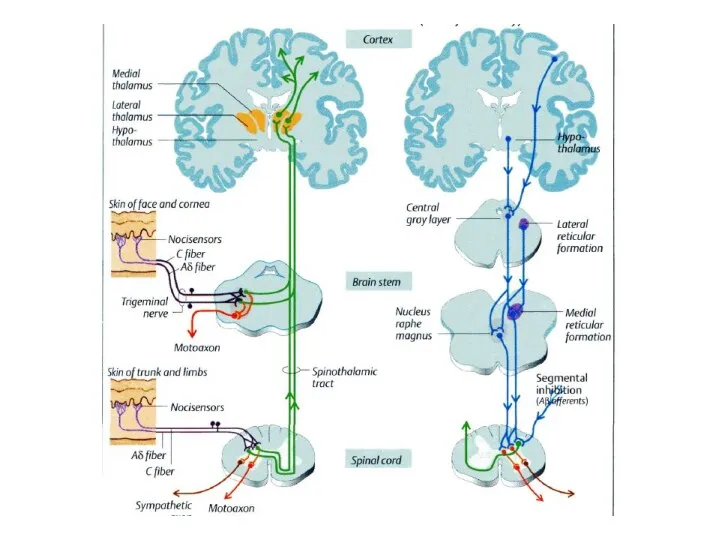 Боль. Локализация боли
Боль. Локализация боли Психопрофилактика
Психопрофилактика Су факторымен байланысты аурулардың алдын алу жөніндегі шаралар
Су факторымен байланысты аурулардың алдын алу жөніндегі шаралар Новые возбудители ИППП
Новые возбудители ИППП Гигиенические аспекты зооантопонозных болезней
Гигиенические аспекты зооантопонозных болезней Клинический случай по конкурсу абдоминальной хирургии
Клинический случай по конкурсу абдоминальной хирургии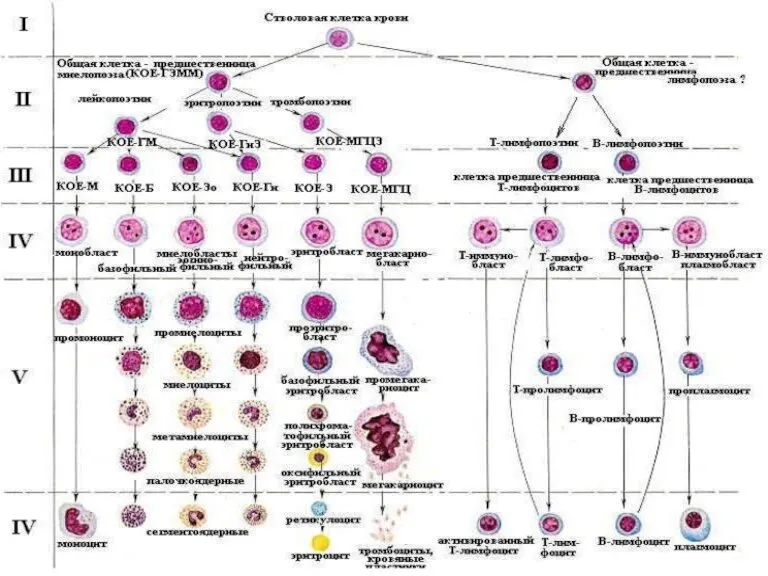 Болезни системы крови
Болезни системы крови Меры и правовые акты Республики Узбекистан, направленные на профилактику спида, наркомании и алкоголизма
Меры и правовые акты Республики Узбекистан, направленные на профилактику спида, наркомании и алкоголизма Общая характеристика травм. Раны и их доврачебная обработка. Правила бинтования и виды повязок
Общая характеристика травм. Раны и их доврачебная обработка. Правила бинтования и виды повязок Основные поражающие факторы табакокурения
Основные поражающие факторы табакокурения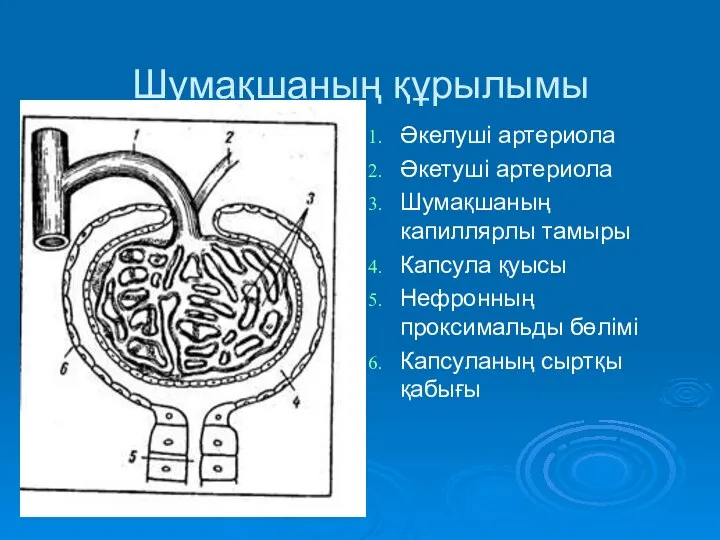 Шумақшаның құрылымы
Шумақшаның құрылымы Эндоваскулярная хирургия
Эндоваскулярная хирургия Профилактические вакцины от гриппа
Профилактические вакцины от гриппа Қан тапсыру-қайырымды іс
Қан тапсыру-қайырымды іс Пневмония. Классификация
Пневмония. Классификация Типовые нарушения водно-солевого обмена
Типовые нарушения водно-солевого обмена Онколитические вирусы. Доставка опухолеспецифичных антигенов антигенпрезентирующим клеткам
Онколитические вирусы. Доставка опухолеспецифичных антигенов антигенпрезентирующим клеткам Закупки в сфере здравоохранения в 2019 году
Закупки в сфере здравоохранения в 2019 году Гирудотерапия
Гирудотерапия