- Основы нейроэндокринной регуляции функций

Содержание
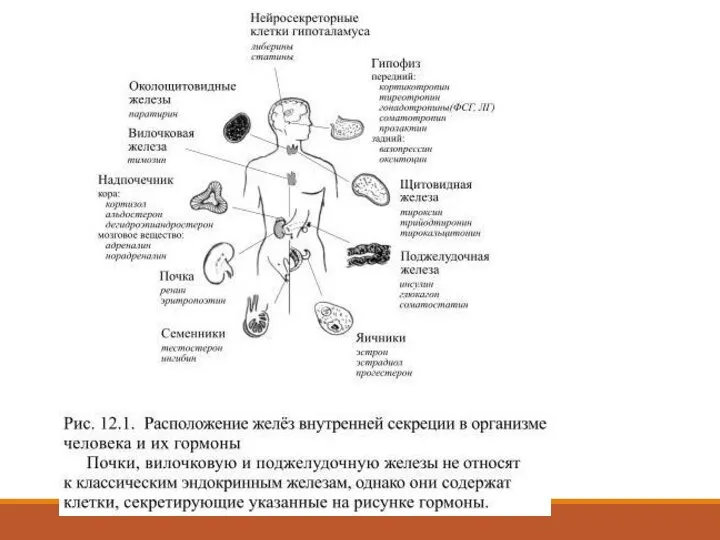
- 2. Эндокринные железы или железы внутренней секреции выделяют в кровь химические вещества, предназначенные для передачи сигнала другим
- 3. Секреция некоторых гормонов (например, инсулина или тестостерона) может изменяться под влиянием нейромедиаторов. Всё это даёт основание
- 5. Происхождение, секреция, транспорт и действие гормонов В зависимости от химической структуры гормоны подразделяются на три группы:
- 6. 3-Стероидные гормоны образуются из молекулы холестерина: это гормоны коры надпочечников (кортизол, кортикостерон, альдостерон, дегидроэпиандростерон) и половых
- 7. Некоторые пептидные гормоны могут образовываться разными способами в результате ферментативного расщепления различных предшественников. В большинстве секретирующих
- 8. Выделение некоторых гормонов увеличивается или уменьшается в зависимости от времени суток (например, выделение соматотропина из передней
- 9. Некоторые гормоны могут поступать к месту действия через лимфу или через внеклеточную жидкость, но большинство из
- 10. Рецепторы для гормонов, которые не могут пройти через клеточную мембрану (белковые и пептидные гормоны, а также
- 11. Стероидные и тиреоидные гормоны, освободившись от транспортных белков, диффундируют через клеточную мембрану (в клетке некоторые из
- 12. Регуляция образования гормонов Концентрация гормонов в крови может изменяться в зависимости от скорости их образования. Существуют
- 13. В результате снижения продукции кортикотропина выделение кортизола из надпочечников уменьшается и соответственно нормализуется уровень этого гормона
- 14. Если же секреция гормонов зависит от управляемого параметра, как, например, в случае образования гормонов поджелудочной железы,
- 15. По такой же схеме регулируется продукция двух гормонов, контролирующих уровень кальция в крови: тирокальцитонин выделяется особыми
- 16. Регуляция по принципу обратной связи осуществляется быстро: темп секреции того или иного гормона может измениться уже
- 18. Роль гипоталамуса в регуляции образования гормонов передней доли гипофиза (гипоталамо-аденогипофизарная система) В 1928 году Эрнст и
- 19. Густая сеть капилляров гипоталамической области собирается в т.н. портальные вены, они доставляют кровь к передней доле
- 21. Выделяемые сравнительно небольшими, т.н. парвоцеллюлярными гипоталамическими нейронами вещества оказались пептидами, одни из которых стимулировали, а другие
- 23. Чтобы понять функциональный смысл взаимоотношений гипоталамуса и гипофиза, следует сопоставить два обстоятельства. Во-первых, в гипоталамус, как
- 24. Ещё два гормона гипофиза – соматотропин и пролактин также играют важную роль в эндокринной регуляции функций.
- 26. Физиологическая роль гормонов аденогипофиза Как уже было сказано, четыре гормона аденогипофиза предназначены для регуляции деятельности других
- 27. Соматотропин (соматотропный гормон – СТГ) или гормон роста действует на большинство тканей, изменяя в них характер
- 28. Одновременно с этим он может препятствовать выделению из внутрисекреторных клеток поджелудочной железы инсулина, который понижает уровень
- 29. Секреции соматотропин-либерина в гипоталамусе способствуют дофамин и серотонин. Гормоны коры надпочечников кортизол и кортикостерон угнетают секрецию
- 30. По-видимому, при раздражении механорецепторов соска во время кормления происходит афферентная импульсация к гипоталамусу, которая и вызывает
- 32. Гипоталамус и гормоны нейрогипофиза Нейрогипофизом называют его заднюю долю, которая образована окончаниями очень крупных (магноцеллюлярных) нервных
- 33. Чем чаще возбуждаются магноцеллюлярные клетки гипоталамуса, тем больше гормонов будет поступать в кровь в нейрогипофизе. Вазопрессин
- 35. При образовании мочи из кровеносных капилляров, образующих клубочки нефронов, фильтруется огромное количество жидкости (до 180 литров
- 36. Основным стимулом для повышенной секреции вазопресссина является увеличение осмотического давления крови и ликвора, что чаще всего
- 37. В связи с её повышенным выведением концентрация солей в крови увеличивается, что приводит к выравниванию осмотического
- 38. При увеличении объёма крови они растягиваются больше обычного, что вызывает повышение частоты сигналов от находящихся в
- 39. Он задерживает воду в почках, а это приводит к увеличению объёма жидкости в сосудистом русле и
- 40. Второй гормон нейрогипофиза – окситоцин действует преимущественно на гладкие мышцы матки и способные к сокращению миоэпителиальные
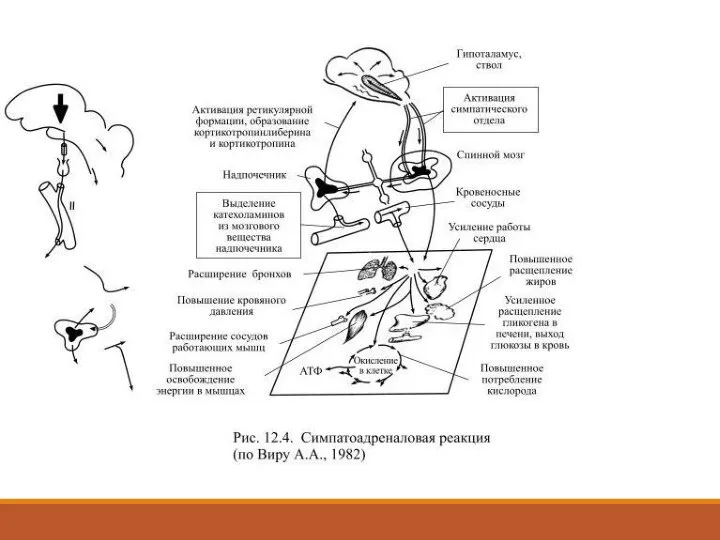
- 42. Гормоны мозгового вещества надпочечников и симпатоадреналовая реакция Находящиеся поблизости от почек парные железы – надпочечники состоят
- 43. Эти гормоны, особенно адреналин, стимулируют расщепление гликогена в печени и мышцах, липидов (жиров) в жировой ткани
- 44. На окисление освободившейся глюкозы нужно израсходовать больше кислорода, чем обычно, а потому необходимо увеличить его доставку
- 45. Выделившиеся в кровь гормоны надпочечников усиливают его действие на клетки-мишени, а кроме того они доставляются током
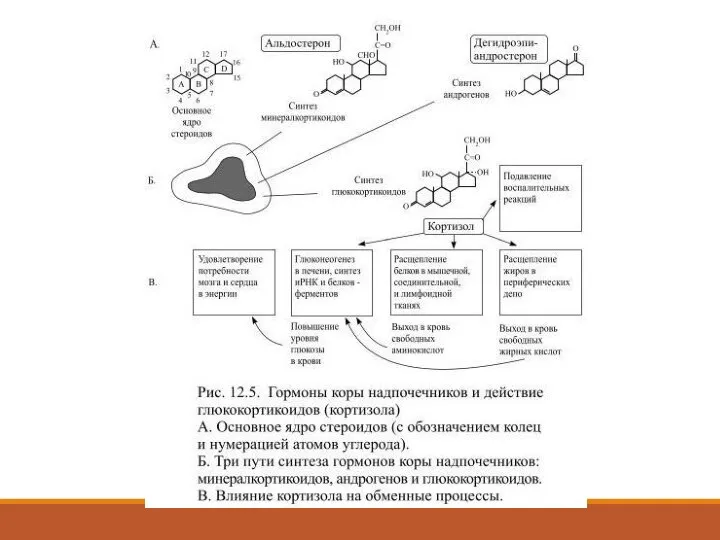
- 46. Гормоны коры надпочечников В трёх морфологически отличающихся областях коры надпочечников из получаемого с пищей или из
- 48. Секрецию кортизола контролирует кортикотропин (АКТГ), вырабатываемый в передней доле гипофиза. Чем больше выделяется в кровь кортикотропина
- 50. Скачать презентацию
Слайд 2Эндокринные железы или железы внутренней секреции выделяют в кровь химические вещества, предназначенные
Эндокринные железы или железы внутренней секреции выделяют в кровь химические вещества, предназначенные

Слайд 3Секреция некоторых гормонов (например, инсулина или тестостерона) может изменяться под влиянием нейромедиаторов.
Секреция некоторых гормонов (например, инсулина или тестостерона) может изменяться под влиянием нейромедиаторов.

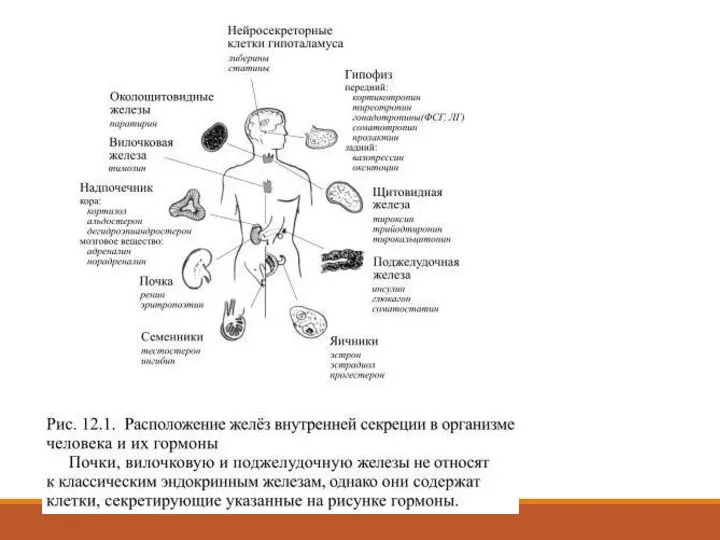
Слайд 5Происхождение, секреция, транспорт и действие гормонов
В зависимости от химической структуры гормоны подразделяются
Происхождение, секреция, транспорт и действие гормонов
В зависимости от химической структуры гормоны подразделяются

Слайд 63-Стероидные гормоны образуются из молекулы холестерина: это гормоны коры надпочечников (кортизол, кортикостерон,
3-Стероидные гормоны образуются из молекулы холестерина: это гормоны коры надпочечников (кортизол, кортикостерон,

Слайд 7Некоторые пептидные гормоны могут образовываться разными способами в результате ферментативного расщепления различных
Некоторые пептидные гормоны могут образовываться разными способами в результате ферментативного расщепления различных

Слайд 8Выделение некоторых гормонов увеличивается или уменьшается в зависимости от времени суток (например,
Выделение некоторых гормонов увеличивается или уменьшается в зависимости от времени суток (например,

Слайд 9Некоторые гормоны могут поступать к месту действия через лимфу или через внеклеточную
Некоторые гормоны могут поступать к месту действия через лимфу или через внеклеточную

Слайд 10Рецепторы для гормонов, которые не могут пройти через клеточную мембрану (белковые и
Рецепторы для гормонов, которые не могут пройти через клеточную мембрану (белковые и

Слайд 11Стероидные и тиреоидные гормоны, освободившись от транспортных белков, диффундируют через клеточную мембрану
Стероидные и тиреоидные гормоны, освободившись от транспортных белков, диффундируют через клеточную мембрану

Слайд 12Регуляция образования гормонов
Концентрация гормонов в крови может изменяться в зависимости от скорости
Регуляция образования гормонов
Концентрация гормонов в крови может изменяться в зависимости от скорости

Слайд 13В результате снижения продукции кортикотропина выделение кортизола из надпочечников уменьшается и соответственно
В результате снижения продукции кортикотропина выделение кортизола из надпочечников уменьшается и соответственно

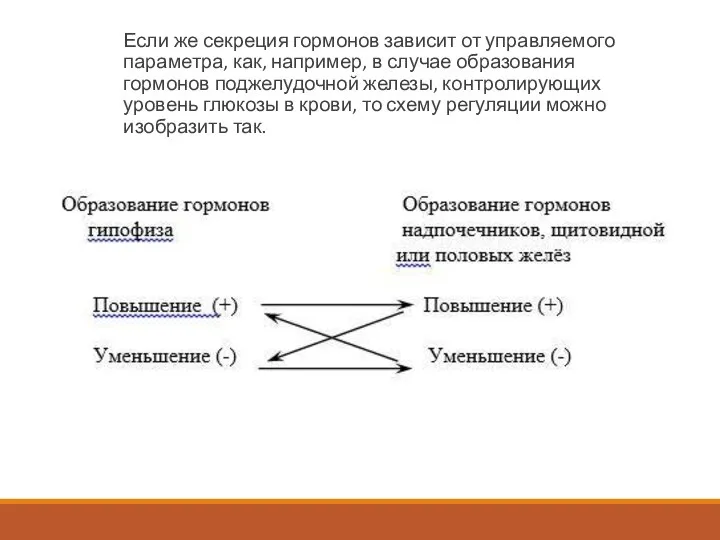
Слайд 14Если же секреция гормонов зависит от управляемого параметра, как, например, в случае
Если же секреция гормонов зависит от управляемого параметра, как, например, в случае
Слайд 15По такой же схеме регулируется продукция двух гормонов, контролирующих уровень кальция в
По такой же схеме регулируется продукция двух гормонов, контролирующих уровень кальция в

Слайд 16Регуляция по принципу обратной связи осуществляется быстро: темп секреции того или иного
Регуляция по принципу обратной связи осуществляется быстро: темп секреции того или иного

Слайд 18Роль гипоталамуса в регуляции образования гормонов передней доли гипофиза (гипоталамо-аденогипофизарная система)
В 1928
Роль гипоталамуса в регуляции образования гормонов передней доли гипофиза (гипоталамо-аденогипофизарная система)
В 1928

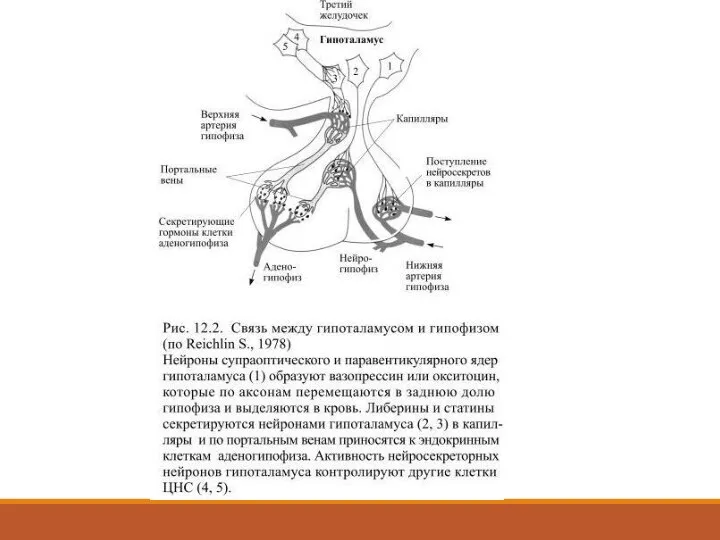
Слайд 19 Густая сеть капилляров гипоталамической области собирается в т.н. портальные вены, они
Густая сеть капилляров гипоталамической области собирается в т.н. портальные вены, они

Слайд 21Выделяемые сравнительно небольшими, т.н. парвоцеллюлярными гипоталамическими нейронами вещества оказались пептидами, одни из
Выделяемые сравнительно небольшими, т.н. парвоцеллюлярными гипоталамическими нейронами вещества оказались пептидами, одни из

Слайд 23Чтобы понять функциональный смысл взаимоотношений гипоталамуса и гипофиза, следует сопоставить два обстоятельства.
Чтобы понять функциональный смысл взаимоотношений гипоталамуса и гипофиза, следует сопоставить два обстоятельства.

Слайд 24Ещё два гормона гипофиза – соматотропин и пролактин также играют важную роль
Ещё два гормона гипофиза – соматотропин и пролактин также играют важную роль

Слайд 26Физиологическая роль гормонов аденогипофиза
Как уже было сказано, четыре гормона аденогипофиза предназначены для
Физиологическая роль гормонов аденогипофиза
Как уже было сказано, четыре гормона аденогипофиза предназначены для

Слайд 27Соматотропин (соматотропный гормон – СТГ) или гормон роста действует на большинство тканей,
Соматотропин (соматотропный гормон – СТГ) или гормон роста действует на большинство тканей,

Слайд 28Одновременно с этим он может препятствовать выделению из внутрисекреторных клеток поджелудочной железы

Слайд 29Секреции соматотропин-либерина в гипоталамусе способствуют дофамин и серотонин. Гормоны коры надпочечников кортизол
Секреции соматотропин-либерина в гипоталамусе способствуют дофамин и серотонин. Гормоны коры надпочечников кортизол

Слайд 30По-видимому, при раздражении механорецепторов соска во время кормления происходит афферентная импульсация к
По-видимому, при раздражении механорецепторов соска во время кормления происходит афферентная импульсация к

Слайд 32Гипоталамус и гормоны нейрогипофиза
Нейрогипофизом называют его заднюю долю, которая образована окончаниями очень
Гипоталамус и гормоны нейрогипофиза
Нейрогипофизом называют его заднюю долю, которая образована окончаниями очень

Слайд 33Чем чаще возбуждаются магноцеллюлярные клетки гипоталамуса, тем больше гормонов будет поступать в
Чем чаще возбуждаются магноцеллюлярные клетки гипоталамуса, тем больше гормонов будет поступать в

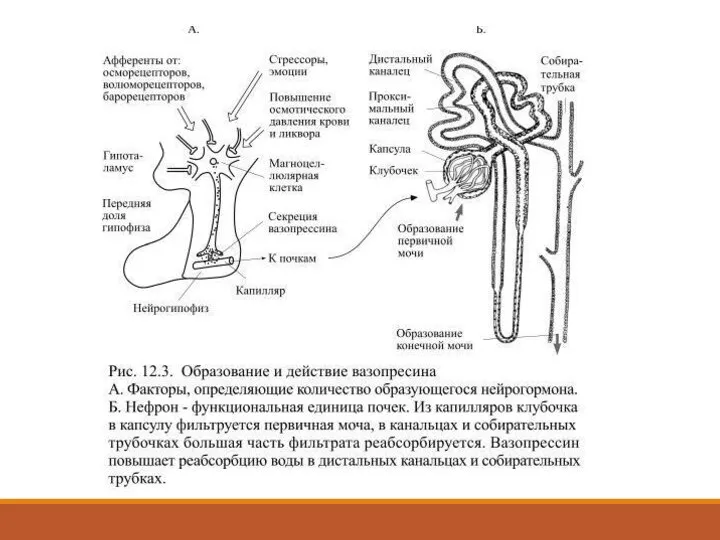
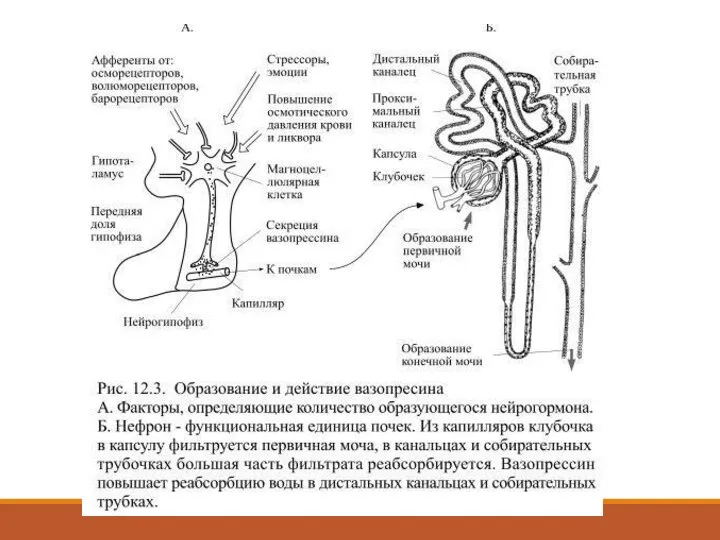
Слайд 35При образовании мочи из кровеносных капилляров, образующих клубочки нефронов, фильтруется огромное количество

Слайд 36Основным стимулом для повышенной секреции вазопресссина является увеличение осмотического давления крови и
Основным стимулом для повышенной секреции вазопресссина является увеличение осмотического давления крови и

Слайд 37В связи с её повышенным выведением концентрация солей в крови увеличивается, что
В связи с её повышенным выведением концентрация солей в крови увеличивается, что

Слайд 38При увеличении объёма крови они растягиваются больше обычного, что вызывает повышение частоты

Слайд 39Он задерживает воду в почках, а это приводит к увеличению объёма жидкости
Он задерживает воду в почках, а это приводит к увеличению объёма жидкости

Слайд 40Второй гормон нейрогипофиза – окситоцин действует преимущественно на гладкие мышцы матки и


Слайд 42Гормоны мозгового вещества надпочечников и симпатоадреналовая реакция
Находящиеся поблизости от почек парные железы
Гормоны мозгового вещества надпочечников и симпатоадреналовая реакция
Находящиеся поблизости от почек парные железы

Слайд 43Эти гормоны, особенно адреналин, стимулируют расщепление гликогена в печени и мышцах, липидов

Слайд 44На окисление освободившейся глюкозы нужно израсходовать больше кислорода, чем обычно, а потому
На окисление освободившейся глюкозы нужно израсходовать больше кислорода, чем обычно, а потому

Слайд 45Выделившиеся в кровь гормоны надпочечников усиливают его действие на клетки-мишени, а кроме
Выделившиеся в кровь гормоны надпочечников усиливают его действие на клетки-мишени, а кроме

Слайд 46Гормоны коры надпочечников
В трёх морфологически отличающихся областях коры надпочечников из получаемого
Гормоны коры надпочечников
В трёх морфологически отличающихся областях коры надпочечников из получаемого

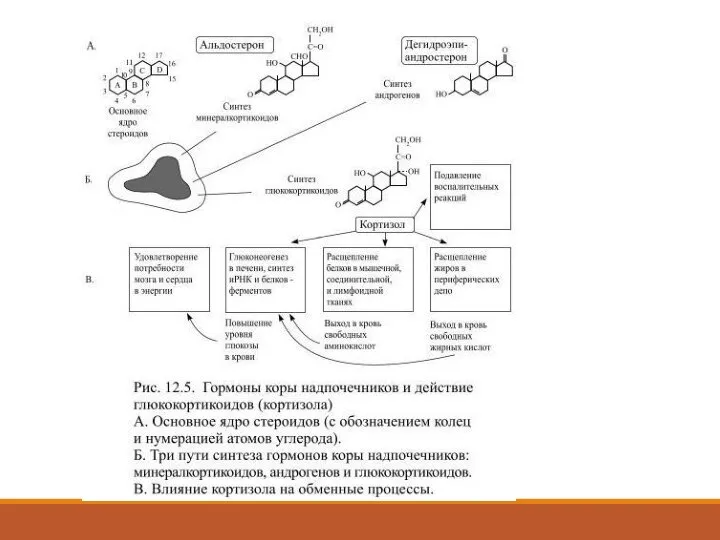
Слайд 48Секрецию кортизола контролирует кортикотропин (АКТГ), вырабатываемый в передней доле гипофиза.
Чем больше
Секрецию кортизола контролирует кортикотропин (АКТГ), вырабатываемый в передней доле гипофиза.
Чем больше

 Космос и математика
Космос и математика Лекарственные средства
Лекарственные средства Беременность при заболеваниях почек и эндокринной патологии
Беременность при заболеваниях почек и эндокринной патологии Моё призвание – медицина
Моё призвание – медицина Привлечение и обучение волонтеров работе с лицами, страдающими тяжелыми заболеваниями, координация работы волонтеров
Привлечение и обучение волонтеров работе с лицами, страдающими тяжелыми заболеваниями, координация работы волонтеров Шок. Классификация шока
Шок. Классификация шока 9. Фармакология ЛП, применяемых при гипертензии
9. Фармакология ЛП, применяемых при гипертензии Возрастные ограничения лекарств. Пояснения к слайдам
Возрастные ограничения лекарств. Пояснения к слайдам Физиотерапия в стоматологии
Физиотерапия в стоматологии Электромиография. Регистрация электрических процессов мышц
Электромиография. Регистрация электрических процессов мышц Острые респираторные вирусные инфекции. Взгляд врачей-терапевтов
Острые респираторные вирусные инфекции. Взгляд врачей-терапевтов Паралич Белла
Паралич Белла Специалисты кардиохирургического Центра принца Султана бен Абдель Азиза
Специалисты кардиохирургического Центра принца Султана бен Абдель Азиза Физическая реабилитация при заболеваниях дыхательной системы (бронхиальная астма)
Физическая реабилитация при заболеваниях дыхательной системы (бронхиальная астма) Рвота беременных. Поражение печени во время беременности
Рвота беременных. Поражение печени во время беременности Лучевая диагностика желчно-каменной болезни
Лучевая диагностика желчно-каменной болезни Как попасть в страну здоровья ?
Как попасть в страну здоровья ? Виды брака в фармации. Законодательная база и ответственность за оборот недоброкачественных лп
Виды брака в фармации. Законодательная база и ответственность за оборот недоброкачественных лп Биомеханика нижней челюсти
Биомеханика нижней челюсти Экологические аспекты канцерогенеза в Центральном Черноземье
Экологические аспекты канцерогенеза в Центральном Черноземье Контролируемый нагрев лазерным излучением внутрикистозной жидкости
Контролируемый нагрев лазерным излучением внутрикистозной жидкости Безглютеновая диета
Безглютеновая диета Пристрій довготривалого моніторування ЕКГ
Пристрій довготривалого моніторування ЕКГ Стажировки для медиков в США
Стажировки для медиков в США Дополнительные методы обследования при кариесе зубов
Дополнительные методы обследования при кариесе зубов Правила здорового сна
Правила здорового сна Памятка владельцам сельскохозяйственной птицы. Ветеринарная служба Республики Башкортостан
Памятка владельцам сельскохозяйственной птицы. Ветеринарная служба Республики Башкортостан ستون فقرات
ستون فقرات