- Регуляция иммунных процессов

Содержание
- 2. COДЕРЖАНИЕ 1.Значение регуляции для иммунных процессов 2.Внутрисистемная регуляция иммунных процессов 3.Печёночная и нейро-эндокринная регуляция 4.Генетическая регуляция
- 3. Иммунные ответы - хорошо регулируемый процесс. Регуляция имеет большое значение для: 1) обеспечения нужного уровня специфичности
- 4. ВНУТРИСИСТЕМНЫЕ МЕХАНИЗМЫ Механизм отрицательной обратной связи Идиотип-антиидиотипические и эрготип-антиэрготипические взаимодействия Цитокины и костимулирующие молекулы Естественные регуляторные
- 5. ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ ПАССИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ Выработка высокоспецифических эффекторных Т-лимфоцитов и иммуноглобулинов приводит
- 7. ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ АКТИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ В эксперименте при одновременном введении животному антигена
- 8. ИДИОТИП-АНТИИДИОТИПИЧЕСКИЕ МОЛЕКУЛЫ Любая антигенспецифическая молекула имеет уникальную конформацию своего антиген-связывающего сайта, который комплементарен структуре антигена. Этот
- 9. Идиотип-антиидиотипические молекулы как рецепторы могут в свою очередь сами распознавать либо связывающие сайты и их части,
- 10. НОБЕЛЕВСКАЯ ПРЕМИЯ (1984) N.K. JERNE Теории иммунитета (клоны лимфоцитов, идиотипические сети)
- 11. ЭРГОТИП- АНТИЭРГОТИПИЧЕСКИЕ ОТВЕТЫ По аналогии с идиотип-антиидиотипическими взаимодействиями на другие рецепторные молекулы (эрготипы, например СD3, CD4,
- 12. РОЛЬ КОСТИМУЛИРУЮЩИХ МОЛЕКУЛ Сигнал с TCR является важнейшим, но не единственным для активации Т-лимфоцитов. Концепция о
- 13. Особенности костимулирующих молекул Одна из пары молекул представлена на Т-лимфоците, другая – на АПК. Чаще сигнал
- 14. Эффекты костимулирующих молекул Действие костимулирующих молекул направлено на усиление сигнала от рецептора/корецептора. Содержат в своем цитоплазматическом
- 15. CD28-CD80/CD86 CD28 спонтанно экспрессируется на большинстве Т-лимфоцитов (80%). CD80 (B7-1) не экспрессируется, СD86 (B7-2) - слабо
- 16. СD40-CD40L (CD154) CD40 конститутивно экспрессируется на АПК (В-лф, ДК, МФ). CD40L экпрессируют активированные Т-клетки, а также
- 17. ICOSL-ICOS ICOS (Inducible costimulator- индуцибельный костимулятор) –экспрессируется на Т-клетках, активированных через TCR и CD28. ICOSL- экпрессируется
- 18. Костимулирующие молекулы, участвующие в презентации антигена
- 20. Коингибиторные рецепторы Известно более 20 мембранных молекул, способных подавлять активацию клеток, вызванную через другие рецепторы. В
- 21. CTLA-4 -CD80/CD86 CTLA-4 (CD152)- гомолог молекулы CD28, связывается с лигандами (CD80/CD86) с большей аффинностью. Экспрессируется на
- 22. PD1- PD-L1, PD-L2 PD1(Programed cell death-1- запрограммированная смерть клетки) – гомолог молекулы CD28. PD1 экспрессируется на
- 23. Блокаторы «контрольных иммунных точек» (immune checkpoint inhibitors) – новые подходы в иммунотерапии рака Молекулы CTLA-4 и
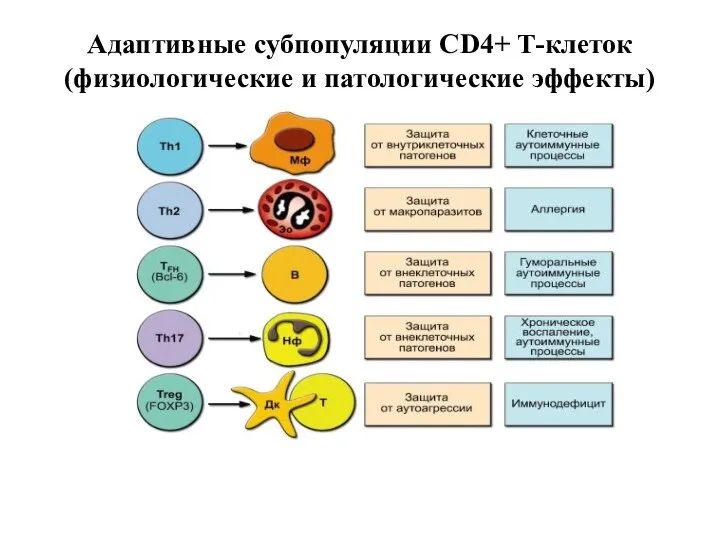
- 24. Адаптивные регуляторные клетки, T-хелпер 1 и T-хелпер 2, идентифицируются, главным образом, по панели цитокинов, которые они
- 25. Различные клетки вовлечены в дифференцировку Тh1 и Тh2. Макрофаги, дендритные клетки и NK-клетки стимулируют развитие Тh1
- 26. На физиологическом уровне, Тh1 включают T-клеточный иммунный ответ и участвуют в переключении синтеза антител; наоборот, Th2
- 28. ИММУНОРЕГУЛЯТОРНЫЕ Т-ЛИМФОЦИТЫ
- 29. Основные субпопуляции CD4+ Т-клеток (пути дифференцировки)
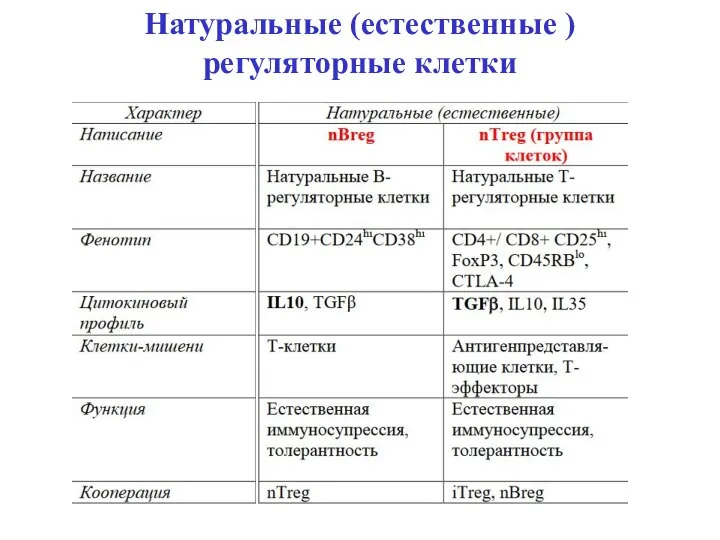
- 30. Натуральные и адаптивные регуляторные Т-клетки (Тreg) Натуральные регуляторные Т-клетки (Тreg) – это группа клеток с фенотипами
- 31. Натуральные (естественные ) регуляторные клетки Климов В.В. Климов В.В.
- 32. Адаптивные (индуцибельные) регуляторные Т-клетки
- 33. ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ Цитокины, небольшие секретируемые белки с молекулярной массой 15-40 кДа, оказывают в гормональных концентрациях через
- 34. Системные (провоспалительные) цитокины: IL1, IL6, TNFα/β Проиммуногенный (провоспалительный) профиль: IL5, IL8, IL12, IL17A-F, IL18, IL19, IL20,
- 35. ЦИТОКИНОВЫЕ ПРОФИЛИ регуляторных лимфоцитов Th1 (CD4+) ? IFNγ, IL2, TNFβ, IL18 Th2 (CD4+) ? IL4, IL5,
- 36. Адаптивные субпопуляции CD4+ Т-клеток (физиологические и патологические эффекты)
- 37. ПЕЧЁНОЧНАЯ РЕГУЛЯЦИЯ Печень является главной “биохимической лабораторией”, которая обеспечивает общую метаболическую регуляцию гомеостаза. Почти все клеточные
- 38. Здоровая печень – иммунологически толерогенный орган Толерогенный режим поддерживают: клетки Купфера, печеночные макрофаги, дендритные клетки, миелоидные
- 39. ЭНДОКРИННАЯ РЕГУЛЯЦИЯ Гормональная регуляция, в упрощённой интерпретации, включает стимулирующее влияние на иммунные процессы соматотропина, ТТГ, трийодтиронина,
- 40. Эффекты глюкокортикоидов В физиологических концентрациях: - регулируют позитивную и негативную селекцию лимфоцитов в ходе лимфопоэза; -
- 41. РЕГУЛЯЦИЯ СО СТОРОНЫ ЦНС Центральная нервная система обеспечивает общую координацию иммунных процессов. Имеется по крайней мере
- 42. РОЛЬ НЕЙРОТРОПИНОВ В ИММУНОРЕГУЛЯЦИИ Нейротропины — семейство структурно связанных факторов роста, регулирующих выживание и функционирование различных
- 43. Нужный уровень специфичности по отношению к исходному антигену. Нужный уровень силы иммунных ответов. Необходимая специфичность достигается:
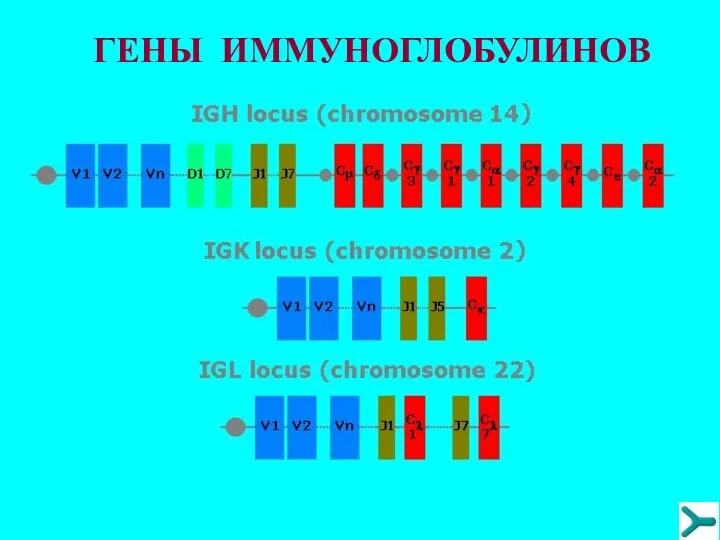
- 44. Гены иммуноглобулинов Количество наследуемого от родителей генетического материала (ДНК), определяющего биосинтез антител - около 120 структурных
- 45. Синтез Н-цепей иммуноглобулинов Рекомбинацию ДНК генов иммуноглобулинов (и генов TCR в T-лимфоцитах) катализируют специальные ферменты -
- 46. ГЕНЫ ИММУНОГЛОБУЛИНОВ
- 48. ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ IgM IgG1 IgG2/3 IgA IgG4 IgE T helper 1 plasma cells IL4,IL5,IL6,IL13
- 49. Рекомбинация ДНК при переключении изотипов иммуноглобулинов
- 50. Механизмы формирования разнообразия генов V-участков иммуноглобулинов и TCR Формирование разнообразия генов для миллионов вариантов V-участков молекул
- 51. ГИПЕРМУТАГЕНЕЗ – запланированное повышение в ходе клональной экспансии В-лимфоцитов частоты точечных мутаций в V-генах, кодирующих вариабельную
- 52. НОБЕЛЕВСКАЯ ПРЕМИЯ (1987) S. TONEGAWA Открытие генетического принципа генерации разнообразия антител
- 53. ГЕНЫ TCR
- 54. Желобок отдельной молекулы HLA у конкретного индивидума может загружать различные антигены с разной эффективностью. Поэтому в
- 56. Генетический полиморфизм МНС Каждый человек наследует определенный набор генов МНС (один на всю жизнь!) Гены МНС
- 58. Скачать презентацию
Слайд 2COДЕРЖАНИЕ
1.Значение регуляции для иммунных процессов
2.Внутрисистемная регуляция иммунных процессов
3.Печёночная и нейро-эндокринная
регуляция
4.Генетическая
COДЕРЖАНИЕ
1.Значение регуляции для иммунных процессов
2.Внутрисистемная регуляция иммунных процессов
3.Печёночная и нейро-эндокринная
регуляция
4.Генетическая

Слайд 3Иммунные ответы - хорошо регулируемый процесс. Регуляция имеет большое значение для:
1)
Иммунные ответы - хорошо регулируемый процесс. Регуляция имеет большое значение для:
1)

Слайд 4ВНУТРИСИСТЕМНЫЕ МЕХАНИЗМЫ
Механизм отрицательной обратной связи
Идиотип-антиидиотипические и эрготип-антиэрготипические взаимодействия
Цитокины и костимулирующие молекулы
Естественные регуляторные
ВНУТРИСИСТЕМНЫЕ МЕХАНИЗМЫ
Механизм отрицательной обратной связи
Идиотип-антиидиотипические и эрготип-антиэрготипические взаимодействия
Цитокины и костимулирующие молекулы
Естественные регуляторные

Слайд 5ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ
ПАССИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ
Выработка высокоспецифических эффекторных Т-лимфоцитов и
ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ
ПАССИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ
Выработка высокоспецифических эффекторных Т-лимфоцитов и


Слайд 7ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ
АКТИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ
В эксперименте при одновременном введении
ПРИНЦИП ОТРИЦАТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ
АКТИВНЫЙ МЕХАНИЗМ СУПРЕССИИ ИММУННЫХ ОТВЕТОВ
В эксперименте при одновременном введении

Слайд 8ИДИОТИП-АНТИИДИОТИПИЧЕСКИЕ МОЛЕКУЛЫ
Любая антигенспецифическая молекула имеет уникальную конформацию своего антиген-связывающего сайта, который комплементарен
ИДИОТИП-АНТИИДИОТИПИЧЕСКИЕ МОЛЕКУЛЫ
Любая антигенспецифическая молекула имеет уникальную конформацию своего антиген-связывающего сайта, который комплементарен

Слайд 9Идиотип-антиидиотипические молекулы как рецепторы могут в свою очередь сами распознавать либо связывающие
Идиотип-антиидиотипические молекулы как рецепторы могут в свою очередь сами распознавать либо связывающие

Слайд 10НОБЕЛЕВСКАЯ ПРЕМИЯ (1984)
N.K. JERNE
Теории иммунитета (клоны лимфоцитов, идиотипические сети)
НОБЕЛЕВСКАЯ ПРЕМИЯ (1984)
N.K. JERNE
Теории иммунитета (клоны лимфоцитов, идиотипические сети)

Слайд 11ЭРГОТИП- АНТИЭРГОТИПИЧЕСКИЕ ОТВЕТЫ
По аналогии с идиотип-антиидиотипическими взаимодействиями на другие рецепторные молекулы (эрготипы,
ЭРГОТИП- АНТИЭРГОТИПИЧЕСКИЕ ОТВЕТЫ
По аналогии с идиотип-антиидиотипическими взаимодействиями на другие рецепторные молекулы (эрготипы,

Слайд 12РОЛЬ КОСТИМУЛИРУЮЩИХ МОЛЕКУЛ
Сигнал с TCR является важнейшим, но не единственным для
РОЛЬ КОСТИМУЛИРУЮЩИХ МОЛЕКУЛ Сигнал с TCR является важнейшим, но не единственным для

Слайд 13Особенности костимулирующих молекул
Одна из пары молекул представлена на Т-лимфоците, другая – на
Особенности костимулирующих молекул
Одна из пары молекул представлена на Т-лимфоците, другая – на

Слайд 14Эффекты костимулирующих молекул
Действие костимулирующих молекул направлено на усиление сигнала от рецептора/корецептора.
Содержат
Эффекты костимулирующих молекул
Действие костимулирующих молекул направлено на усиление сигнала от рецептора/корецептора.
Содержат
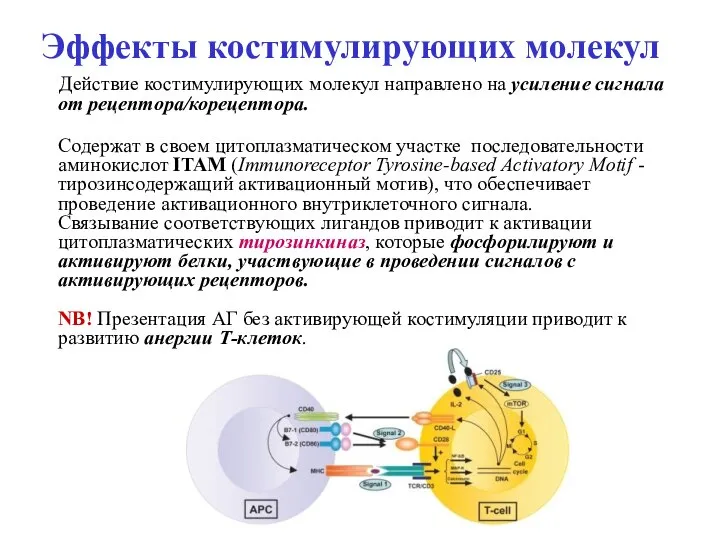
Слайд 15CD28-CD80/CD86
CD28 спонтанно экспрессируется на большинстве Т-лимфоцитов (80%).
CD80 (B7-1) не экспрессируется, СD86 (B7-2)
CD28-CD80/CD86
CD28 спонтанно экспрессируется на большинстве Т-лимфоцитов (80%).
CD80 (B7-1) не экспрессируется, СD86 (B7-2)

Слайд 16СD40-CD40L (CD154)
CD40 конститутивно экспрессируется на АПК (В-лф, ДК, МФ).
CD40L экпрессируют активированные Т-клетки,
СD40-CD40L (CD154)
CD40 конститутивно экспрессируется на АПК (В-лф, ДК, МФ).
CD40L экпрессируют активированные Т-клетки,

Слайд 17ICOSL-ICOS
ICOS (Inducible costimulator- индуцибельный костимулятор) –экспрессируется на Т-клетках, активированных через TCR и
ICOSL-ICOS
ICOS (Inducible costimulator- индуцибельный костимулятор) –экспрессируется на Т-клетках, активированных через TCR и

Слайд 18Костимулирующие молекулы, участвующие в презентации антигена
Костимулирующие молекулы, участвующие в презентации антигена
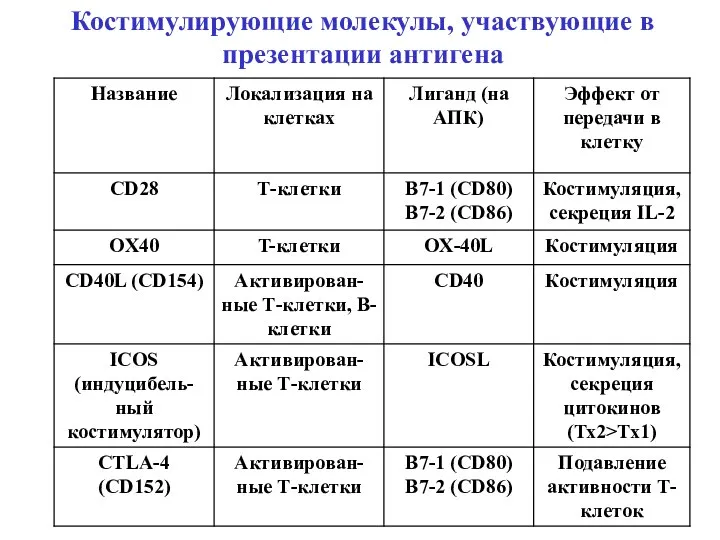
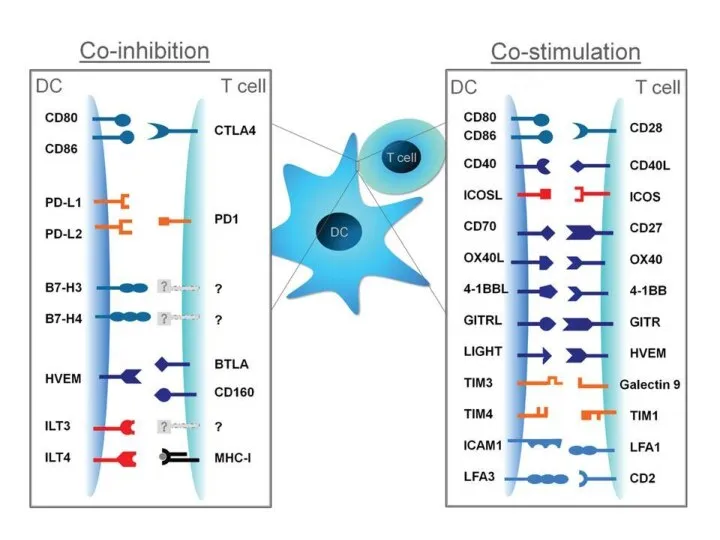
Слайд 20Коингибиторные рецепторы
Известно более 20 мембранных молекул, способных подавлять активацию клеток, вызванную
Коингибиторные рецепторы
Известно более 20 мембранных молекул, способных подавлять активацию клеток, вызванную

Слайд 21CTLA-4 -CD80/CD86
CTLA-4 (CD152)- гомолог молекулы CD28, связывается с лигандами (CD80/CD86) с большей
CTLA-4 -CD80/CD86
CTLA-4 (CD152)- гомолог молекулы CD28, связывается с лигандами (CD80/CD86) с большей

Слайд 22PD1- PD-L1, PD-L2
PD1(Programed cell death-1- запрограммированная смерть клетки) – гомолог молекулы CD28.
PD1
PD1- PD-L1, PD-L2
PD1(Programed cell death-1- запрограммированная смерть клетки) – гомолог молекулы CD28.
PD1

Слайд 23Блокаторы «контрольных иммунных точек» (immune checkpoint inhibitors) – новые подходы в иммунотерапии
Блокаторы «контрольных иммунных точек» (immune checkpoint inhibitors) – новые подходы в иммунотерапии
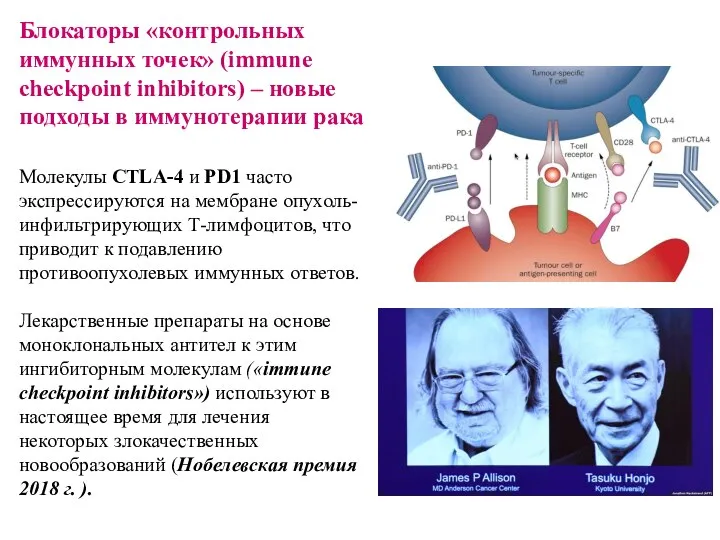
Слайд 24Адаптивные регуляторные клетки, T-хелпер 1 и T-хелпер 2, идентифицируются, главным образом, по
Адаптивные регуляторные клетки, T-хелпер 1 и T-хелпер 2, идентифицируются, главным образом, по

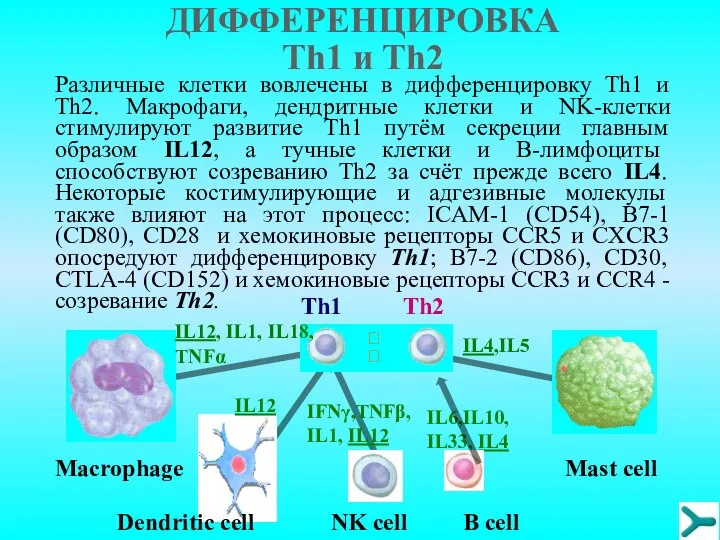
Слайд 25Различные клетки вовлечены в дифференцировку Тh1 и Тh2. Макрофаги, дендритные клетки и
Различные клетки вовлечены в дифференцировку Тh1 и Тh2. Макрофаги, дендритные клетки и
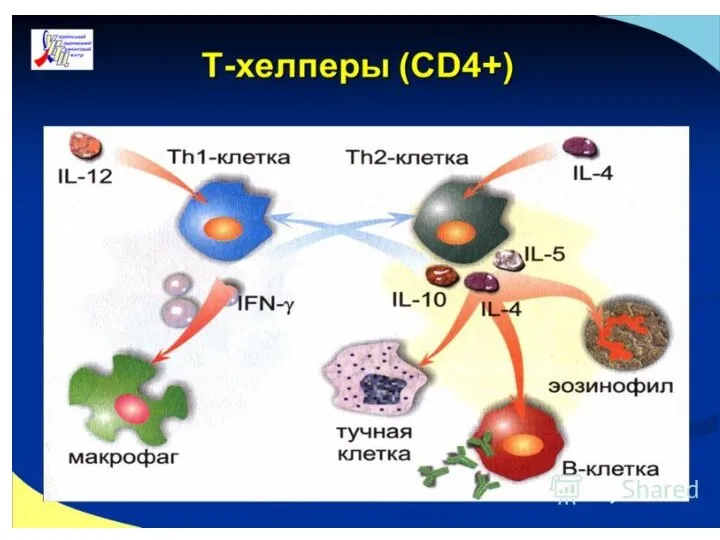
Слайд 26На физиологическом уровне, Тh1 включают T-клеточный иммунный ответ и участвуют в переключении
На физиологическом уровне, Тh1 включают T-клеточный иммунный ответ и участвуют в переключении

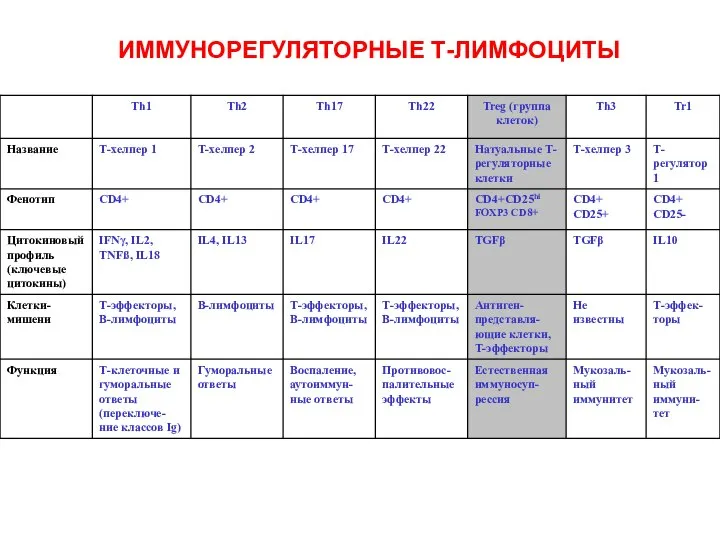
Слайд 28ИММУНОРЕГУЛЯТОРНЫЕ Т-ЛИМФОЦИТЫ
ИММУНОРЕГУЛЯТОРНЫЕ Т-ЛИМФОЦИТЫ
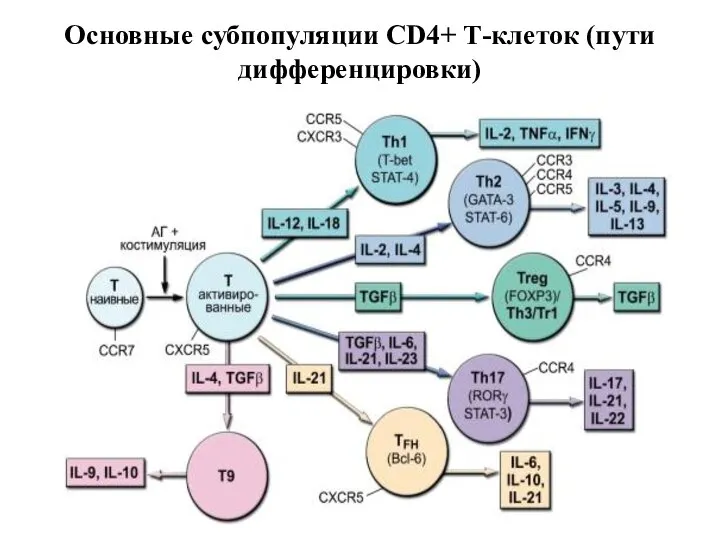
Слайд 29Основные субпопуляции CD4+ Т-клеток (пути дифференцировки)
Основные субпопуляции CD4+ Т-клеток (пути дифференцировки)
Слайд 30Натуральные и адаптивные регуляторные Т-клетки (Тreg)
Натуральные регуляторные Т-клетки (Тreg) – это группа
Натуральные и адаптивные регуляторные Т-клетки (Тreg)
Натуральные регуляторные Т-клетки (Тreg) – это группа

Слайд 31Натуральные (естественные ) регуляторные клетки
Климов В.В.
Климов В.В.
Натуральные (естественные ) регуляторные клетки
Климов В.В.
Климов В.В.
Слайд 32Адаптивные (индуцибельные) регуляторные Т-клетки
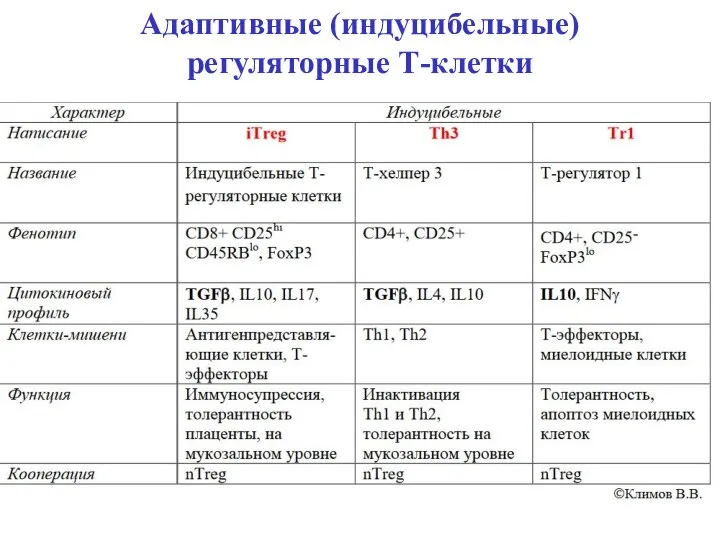
Адаптивные (индуцибельные) регуляторные Т-клетки
Слайд 33ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ
Цитокины, небольшие секретируемые белки с молекулярной массой 15-40 кДа, оказывают в
ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ
Цитокины, небольшие секретируемые белки с молекулярной массой 15-40 кДа, оказывают в

Слайд 34Системные (провоспалительные) цитокины: IL1, IL6, TNFα/β
Проиммуногенный (провоспалительный) профиль:
IL5, IL8, IL12, IL17A-F, IL18,
Системные (провоспалительные) цитокины: IL1, IL6, TNFα/β
Проиммуногенный (провоспалительный) профиль:
IL5, IL8, IL12, IL17A-F, IL18,

Слайд 35ЦИТОКИНОВЫЕ ПРОФИЛИ регуляторных лимфоцитов
Th1 (CD4+)
?
IFNγ, IL2, TNFβ, IL18
Th2 (CD4+)
?
IL4, IL5, IL6, IL10,
ЦИТОКИНОВЫЕ ПРОФИЛИ регуляторных лимфоцитов
Th1 (CD4+)
?
IFNγ, IL2, TNFβ, IL18
Th2 (CD4+)
?
IL4, IL5, IL6, IL10,

Слайд 36Адаптивные субпопуляции CD4+ Т-клеток (физиологические и патологические эффекты)
Адаптивные субпопуляции CD4+ Т-клеток (физиологические и патологические эффекты)
Слайд 37ПЕЧЁНОЧНАЯ РЕГУЛЯЦИЯ
Печень является главной “биохимической лабораторией”, которая обеспечивает общую метаболическую регуляцию гомеостаза.
ПЕЧЁНОЧНАЯ РЕГУЛЯЦИЯ
Печень является главной “биохимической лабораторией”, которая обеспечивает общую метаболическую регуляцию гомеостаза.

Слайд 38Здоровая печень – иммунологически толерогенный орган
Толерогенный режим поддерживают:
клетки Купфера, печеночные
Здоровая печень – иммунологически толерогенный орган
Толерогенный режим поддерживают:
клетки Купфера, печеночные

Слайд 39ЭНДОКРИННАЯ РЕГУЛЯЦИЯ
Гормональная регуляция, в упрощённой интерпретации, включает стимулирующее влияние на иммунные
ЭНДОКРИННАЯ РЕГУЛЯЦИЯ
Гормональная регуляция, в упрощённой интерпретации, включает стимулирующее влияние на иммунные

Слайд 40Эффекты глюкокортикоидов
В физиологических концентрациях:
- регулируют позитивную и негативную селекцию лимфоцитов в
Эффекты глюкокортикоидов
В физиологических концентрациях:
- регулируют позитивную и негативную селекцию лимфоцитов в

Слайд 41РЕГУЛЯЦИЯ СО СТОРОНЫ ЦНС
Центральная нервная система обеспечивает общую координацию иммунных процессов.
РЕГУЛЯЦИЯ СО СТОРОНЫ ЦНС
Центральная нервная система обеспечивает общую координацию иммунных процессов.

Слайд 42РОЛЬ НЕЙРОТРОПИНОВ В ИММУНОРЕГУЛЯЦИИ
Нейротропины — семейство структурно связанных факторов роста, регулирующих
РОЛЬ НЕЙРОТРОПИНОВ В ИММУНОРЕГУЛЯЦИИ
Нейротропины — семейство структурно связанных факторов роста, регулирующих

Слайд 43Нужный уровень специфичности по отношению к исходному антигену.
Нужный уровень силы иммунных ответов.
Нужный уровень специфичности по отношению к исходному антигену.
Нужный уровень силы иммунных ответов.

Слайд 44Гены иммуноглобулинов
Количество наследуемого от родителей генетического материала (ДНК), определяющего биосинтез антител
Гены иммуноглобулинов
Количество наследуемого от родителей генетического материала (ДНК), определяющего биосинтез антител

Слайд 45Синтез Н-цепей иммуноглобулинов
Рекомбинацию ДНК генов иммуноглобулинов (и генов TCR в T-лимфоцитах)
Синтез Н-цепей иммуноглобулинов
Рекомбинацию ДНК генов иммуноглобулинов (и генов TCR в T-лимфоцитах)

Слайд 46ГЕНЫ ИММУНОГЛОБУЛИНОВ
ГЕНЫ ИММУНОГЛОБУЛИНОВ
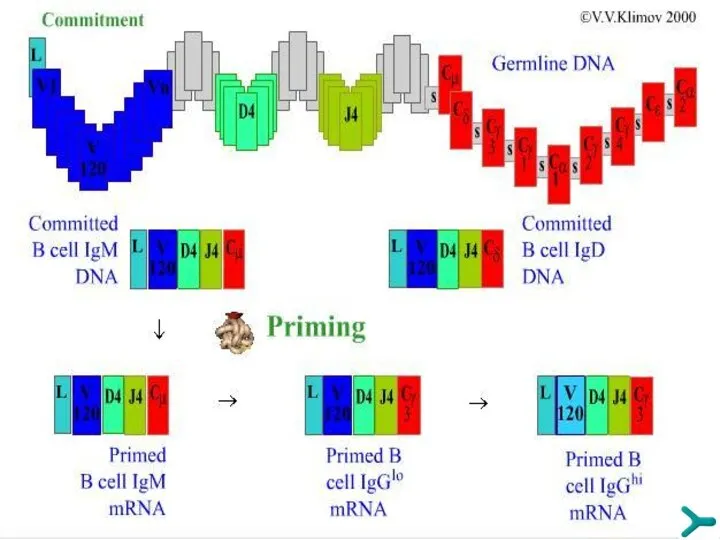
Слайд 48ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ
IgM
IgG1
IgG2/3
IgA
IgG4
IgE
T helper 1
plasma cells
IL4,IL5,IL6,IL13
IL4,IL6
IL5,IL6,IL10
IL4,IL13
IFNγ,TNF
TNF
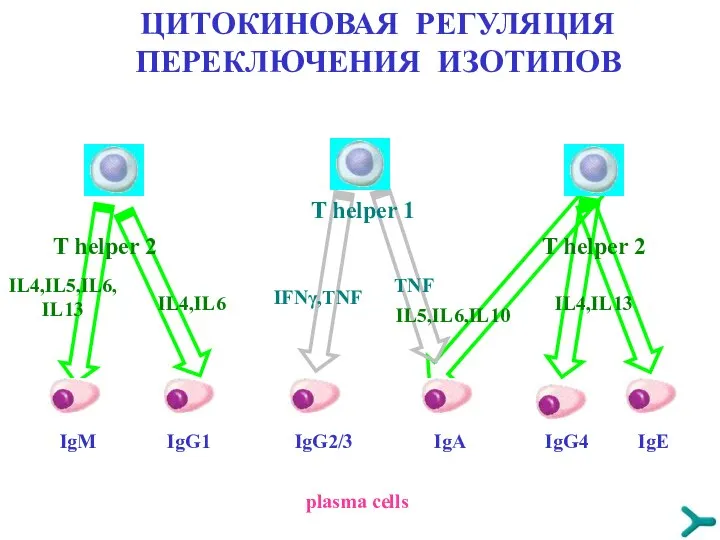
ЦИТОКИНОВАЯ РЕГУЛЯЦИЯ ПЕРЕКЛЮЧЕНИЯ ИЗОТИПОВ
IgM
IgG1
IgG2/3
IgA
IgG4
IgE
T helper 1
plasma cells
IL4,IL5,IL6,IL13
IL4,IL6
IL5,IL6,IL10
IL4,IL13
IFNγ,TNF
TNF
Слайд 49Рекомбинация ДНК при переключении изотипов иммуноглобулинов
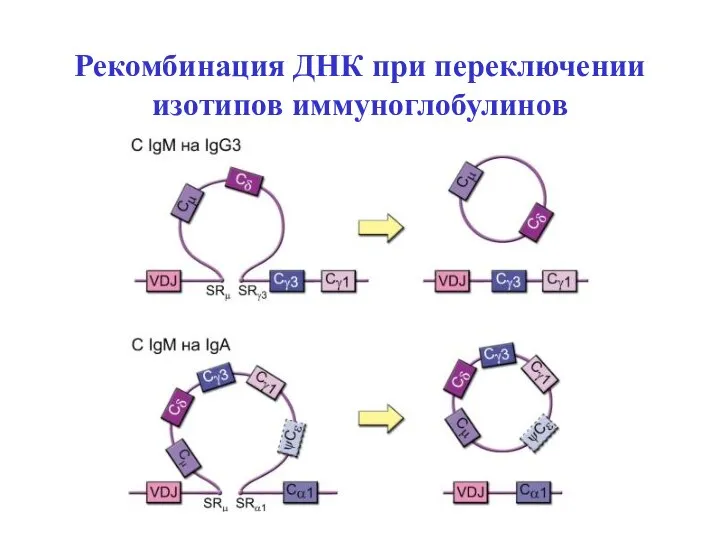
Рекомбинация ДНК при переключении изотипов иммуноглобулинов
Слайд 50Механизмы формирования разнообразия генов V-участков иммуноглобулинов и TCR
Формирование разнообразия генов для
Механизмы формирования разнообразия генов V-участков иммуноглобулинов и TCR
Формирование разнообразия генов для

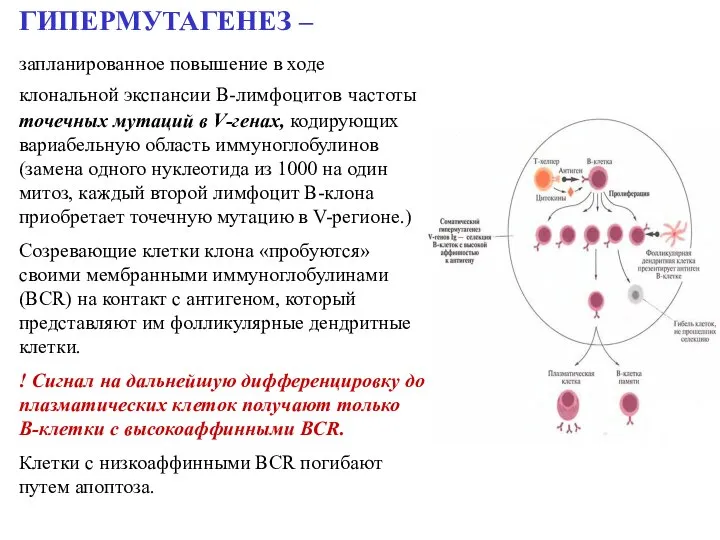
Слайд 51ГИПЕРМУТАГЕНЕЗ –
запланированное повышение в ходе клональной экспансии В-лимфоцитов частоты точечных мутаций в
ГИПЕРМУТАГЕНЕЗ –
запланированное повышение в ходе клональной экспансии В-лимфоцитов частоты точечных мутаций в
Слайд 52НОБЕЛЕВСКАЯ ПРЕМИЯ (1987)
S. TONEGAWA
Открытие генетического принципа генерации разнообразия антител
НОБЕЛЕВСКАЯ ПРЕМИЯ (1987)
S. TONEGAWA
Открытие генетического принципа генерации разнообразия антител

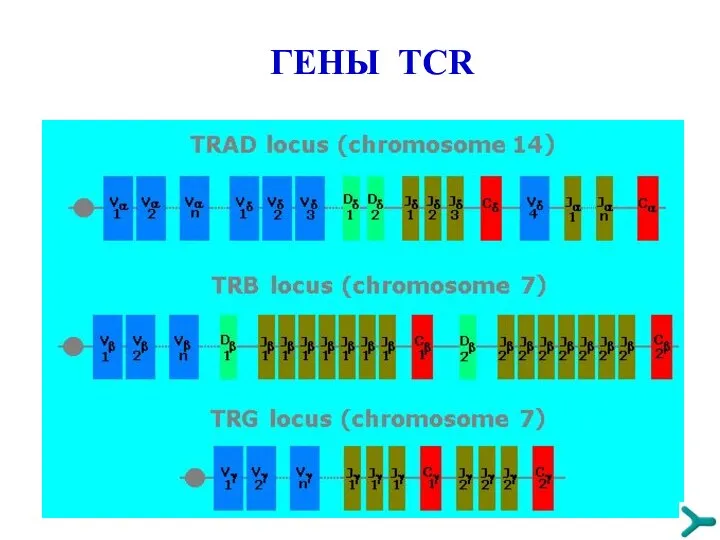
Слайд 53ГЕНЫ TCR
ГЕНЫ TCR
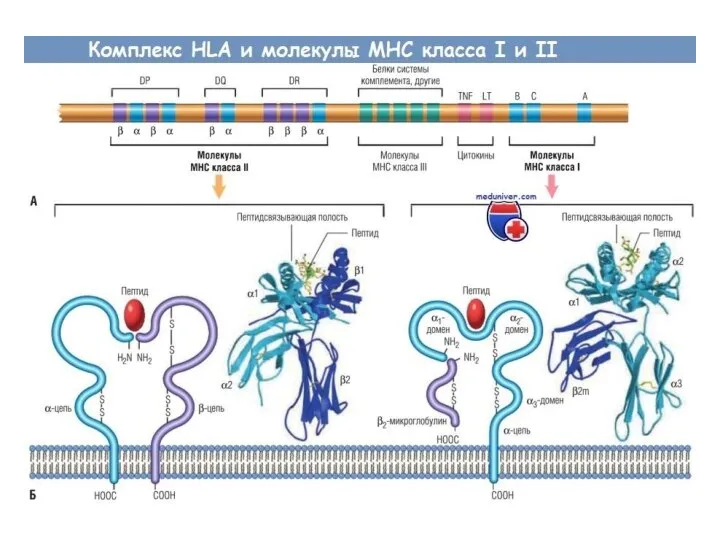
Слайд 54Желобок отдельной молекулы HLA у конкретного индивидума может загружать различные антигены с
Желобок отдельной молекулы HLA у конкретного индивидума может загружать различные антигены с

Слайд 56Генетический полиморфизм МНС
Каждый человек наследует определенный набор генов МНС (один на всю
Генетический полиморфизм МНС
Каждый человек наследует определенный набор генов МНС (один на всю

 Симптомы и синдромы при заболеваниях желудка. Хронический гастрит, язва желудка и 12-перстной кишки
Симптомы и синдромы при заболеваниях желудка. Хронический гастрит, язва желудка и 12-перстной кишки Асфиксия новорожденных
Асфиксия новорожденных Сестринский процесс при атопическом дерматите
Сестринский процесс при атопическом дерматите Трансъюгулярное внутрипеченочное портосистемное шунтирование (TIPS)
Трансъюгулярное внутрипеченочное портосистемное шунтирование (TIPS) Алкоголизм. Только факты
Алкоголизм. Только факты Протейперы
Протейперы МРТ сердца у детей: интересные случаи
МРТ сердца у детей: интересные случаи Антидоты (противоядия): назначение, область и порядок применения.Тема 4
Антидоты (противоядия): назначение, область и порядок применения.Тема 4 Ранний период беременности
Ранний период беременности Молекулярная биология болезни Альцгеймера
Молекулярная биология болезни Альцгеймера Доброякісні захворювання шийки матки, шляхи оптимізації деструктивних методів лікування
Доброякісні захворювання шийки матки, шляхи оптимізації деструктивних методів лікування Микоплазмоз. Микоплазмоздың түрлері
Микоплазмоз. Микоплазмоздың түрлері Гельминтозы. Санитарно-гигиенические мероприятия
Гельминтозы. Санитарно-гигиенические мероприятия Ситуация в области остеопороза в республике Молдова
Ситуация в области остеопороза в республике Молдова Автоматизированное рабочее место врача
Автоматизированное рабочее место врача Дородовый уход
Дородовый уход Основные поражающие факторы алкоголизма
Основные поражающие факторы алкоголизма Правила здорового питания
Правила здорового питания Комаларнинг келиб чиқиш сабабини инобатга олиниб
Комаларнинг келиб чиқиш сабабини инобатга олиниб Стрим по анатомии. Опорно-двигательная система
Стрим по анатомии. Опорно-двигательная система Композитные пломбировочные материалы
Композитные пломбировочные материалы Операции на костях
Операции на костях Физиология новорожденных. Преимущества ношения на руках
Физиология новорожденных. Преимущества ношения на руках Технологические стадии производства стерильных растворов, контроль критических стадий и промежуточной продукции
Технологические стадии производства стерильных растворов, контроль критических стадий и промежуточной продукции Основные модели взаимоотношений врача и пациента
Основные модели взаимоотношений врача и пациента Вывихи зубов
Вывихи зубов Аускультация легких
Аускультация легких Эктопические ресницы в ветеринарной офтальмологии. Тема 6
Эктопические ресницы в ветеринарной офтальмологии. Тема 6